
Lipid Polymorphism of the Subchloroplast—Granum and Stroma Thylakoid Membrane—Particles. I. 31P-NMR Spectroscopy

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Granum and Stroma Thylakoid Membranes
2.2. Lipase Treatments
2.3. 31P-NMR Measurements
3. Results
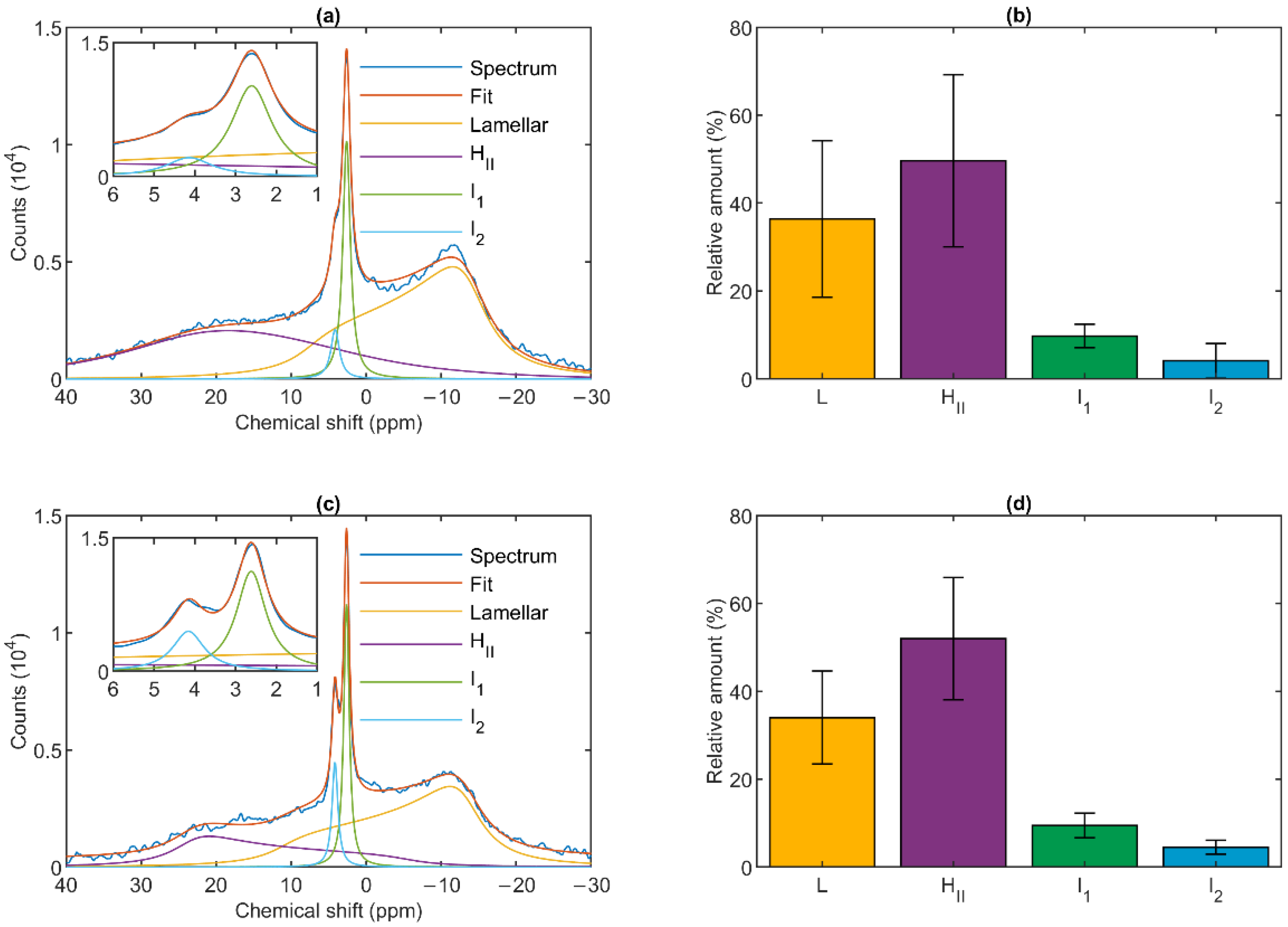
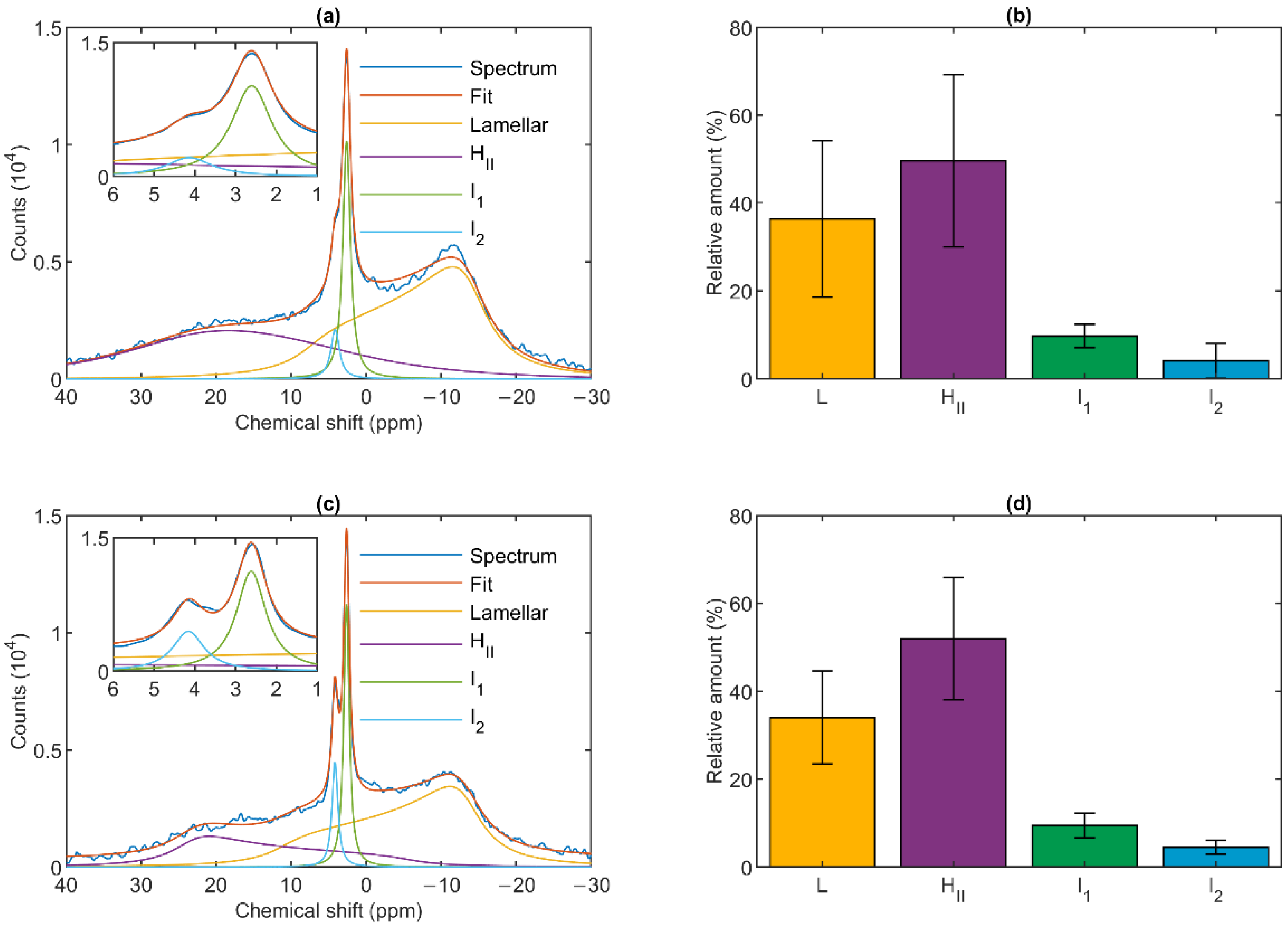
3.1. 31P-NMR Fingerprints of Lipid Phases in Isolated Granum and Stroma Thylakoid Membranes
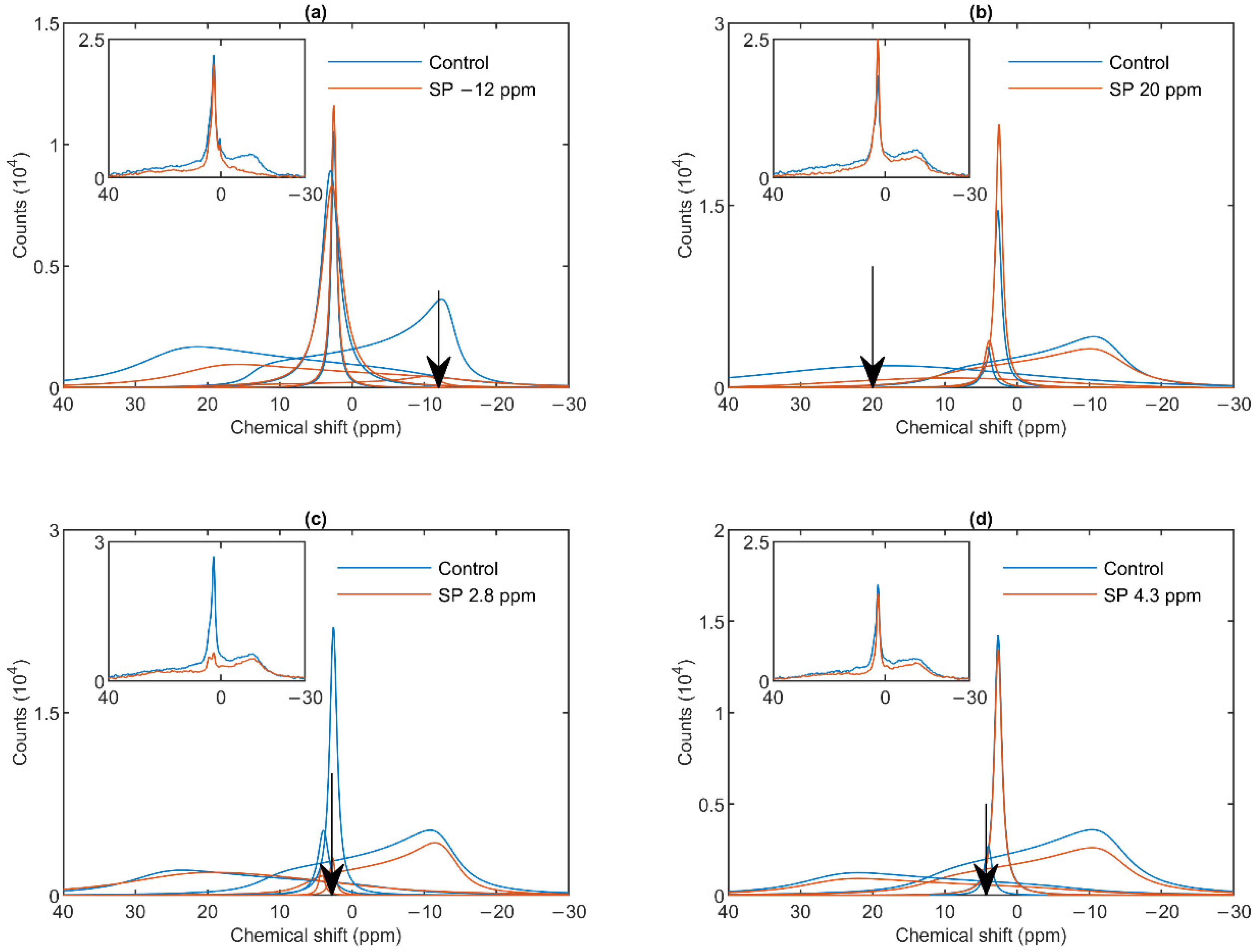
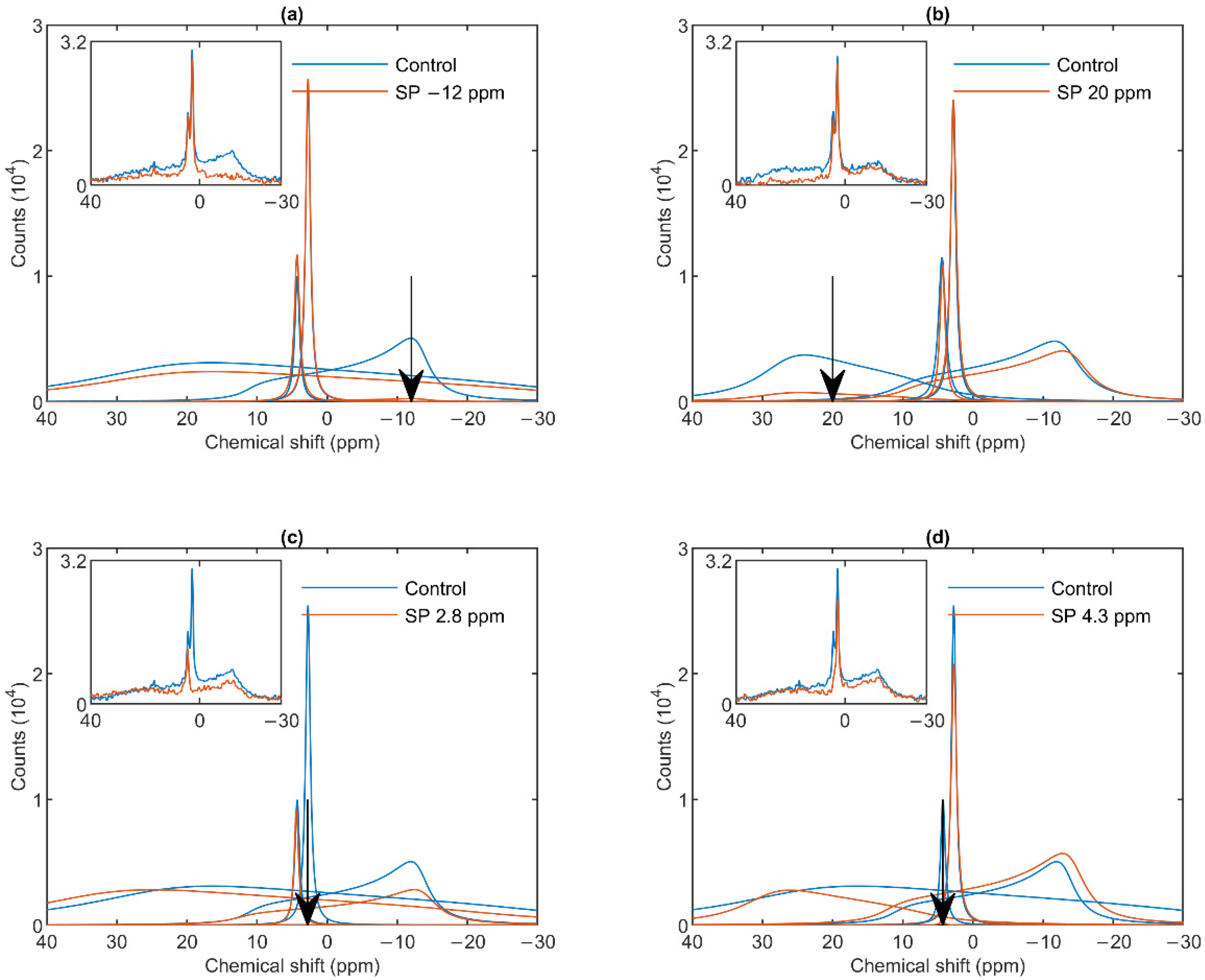
3.2. Saturation-Transfer Experiments
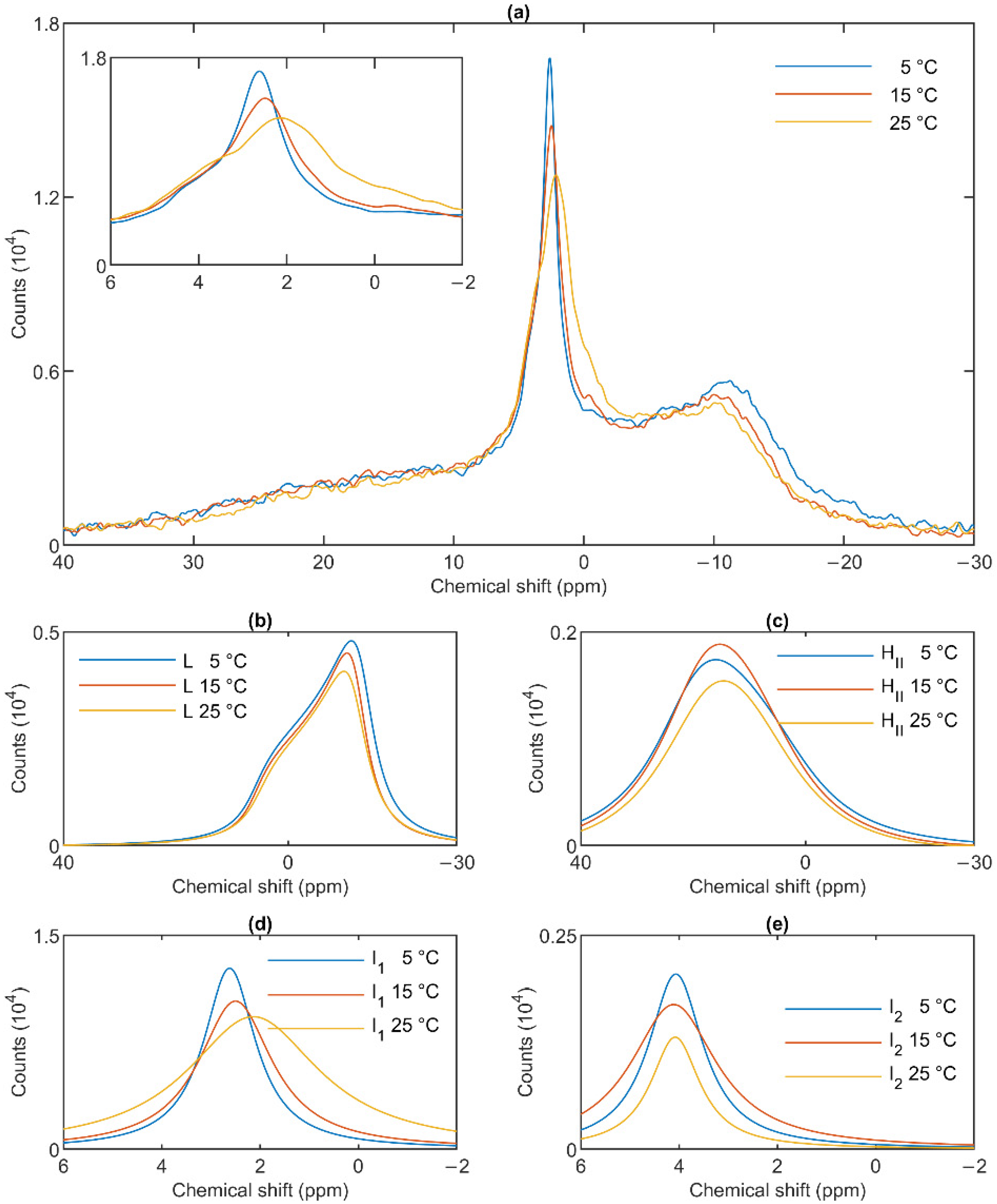
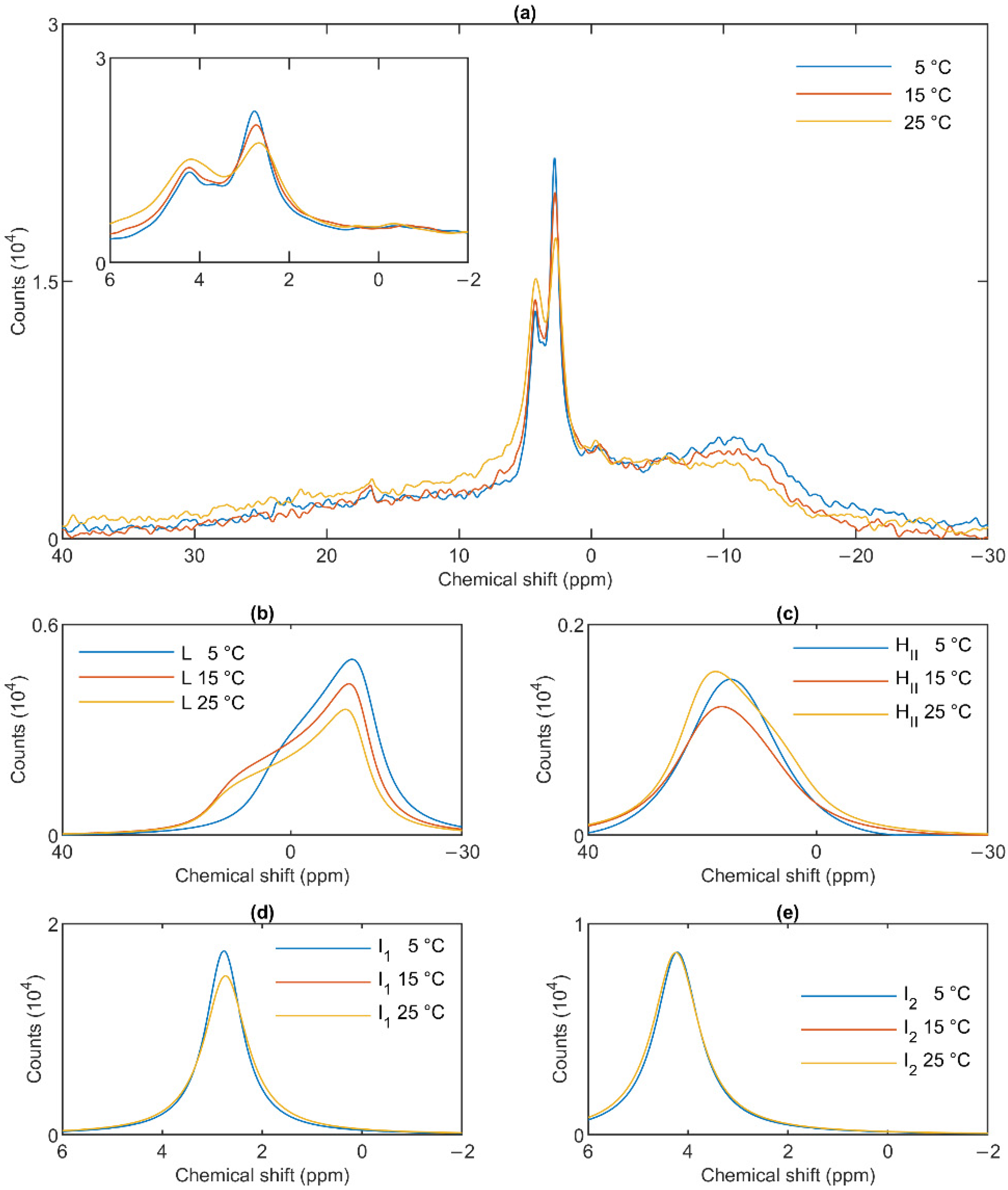
3.3. Temperature Dependences
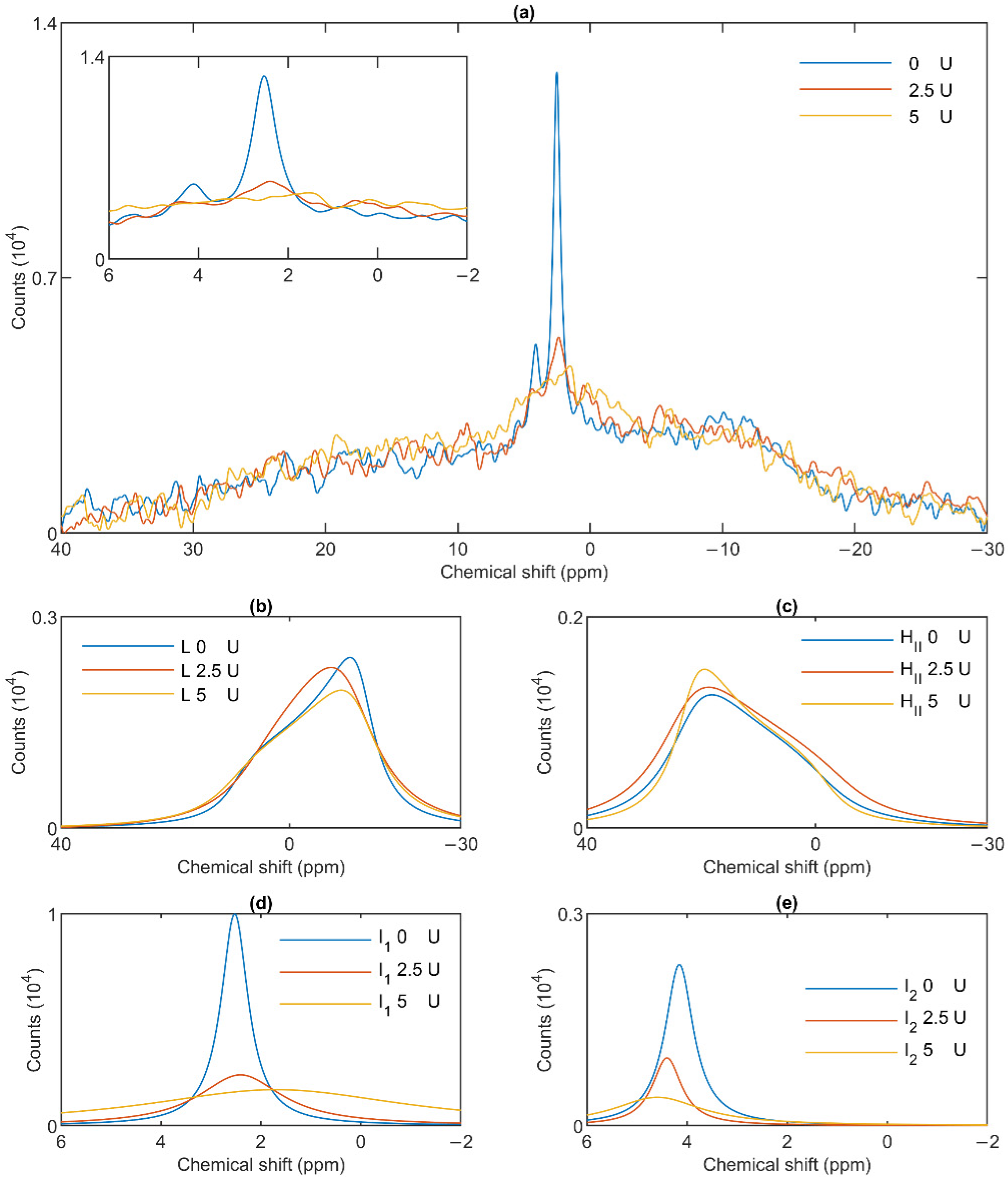
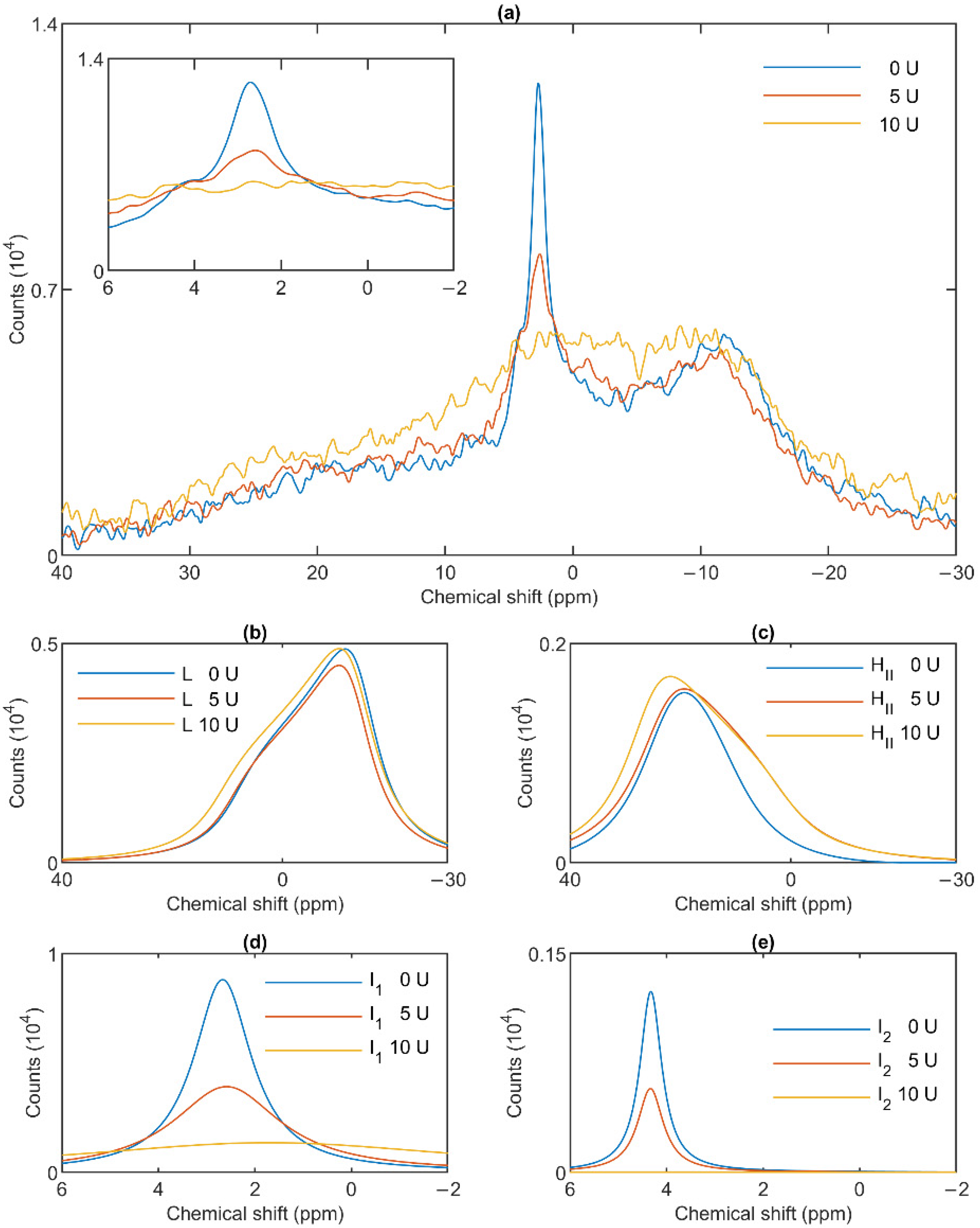
3.4. Effect of Wheat Germ Lipase
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mitchell, P. Chemiosmotic coupling in oxidative and photosynthetic phosphorylation. Biol. Rev. Camb. Philos. Soc. 1966, 41, 445–502. [Google Scholar] [CrossRef] [PubMed]
- Singer, S.J.; Nicolson, G.L. The Fluid Mosaic Model of the Structure of Cell Membranes. Science 1972, 175, 720–731. [Google Scholar] [CrossRef]
- Seddon, J.M.; Templer, R.H. Chapter 3—Polymorphism of Lipid-Water Systems. In Handbook of Biological Physics; Lipowsky, R., Sackmann, E., Eds.; Elsevier: Amsterdam, The Netherlands, 1995; Volume 1, pp. 97–160. [Google Scholar]
- Williams, W.P. The Physical Properties of Thylakoid Membrane Lipids and Their Relation to Photosynthesis. In Lipids in Photosynthesis: Structure, Function and Genetics; Paul-André, S., Norio, M., Eds.; Springer: Dordrecht, The Netherlands, 1998; pp. 103–118. [Google Scholar]
- Douce, R.; Joyard, J. Biosynthesis of Thylakoid Membrane Lipids. In Oxygenic Photosynthesis: The Light Reactions; Ort, D.R., Yocum, C.F., Heichel, I.F., Eds.; Springer: Dordrecht, The Netherlands, 1996; pp. 69–101. [Google Scholar]
- Luzzati, V.; Reisshus, F.; Rivas, E.; Gulikkrz, T. Structure and polymorphism in lipid-water systems and their possible biological implications. Ann. N. Y. Acad. Sci. 1966, 137, 409–413. [Google Scholar] [CrossRef]
- Luzzati, V.; Delacroix, H.; Gulik, A.; Gulik-Krzywicki, T.; Mariani, P.; Vargas, R. The cubic phases of lipids. In Lipid Polymorphism and Membrane Properties; Epand, R.M., Ed.; Current Topics in Membranes; Elsevier: Amsterdam, The Netherlands, 1997; Volume 44, pp. 3–24. [Google Scholar]
- Epand, R.M. Lipid polymorphism and protein-lipid interactions. Biochim. Biophys. Acta 1998, 1376, 353–368. [Google Scholar] [CrossRef]
- Shipley, G.G.; Green, J.P.; Nichols, B.W. Phase behavior of monogalactosyl, digalactosyl, and sulfoquinovosyl diglycerides. Biochim. Biophys. Acta 1973, 311, 531–544. [Google Scholar] [CrossRef]
- van Eerden, F.J.; de Jong, D.H.; de Vries, A.H.; Wassenaar, T.A.; Marrink, S.J. Characterization of thylakoid lipid membranes from cyanobacteria and higher plants by molecular dynamics simulations. Biochim. Biophys. Acta 2015, 1848, 1319–1330. [Google Scholar] [CrossRef] [Green Version]
- Demé, B.; Cataye, C.; Block, M.A.; Maréchal, E.; Jouhet, J. Contribution of galactoglycerolipids to the 3-dimensional architecture of thylakoids. FASEB J. 2014, 28, 3373–3383. [Google Scholar] [CrossRef] [Green Version]
- Simidjiev, I.; Stoylova, S.; Amenitsch, H.; Javorfi, T.; Mustárdy, L.; Laggner, P.; Holzenburg, A.; Garab, G. Self-assembly of large, ordered lamellae from non-bilayer lipids and integral membrane proteins in vitro. Proc. Natl. Acad. Sci. USA 2000, 97, 1473–1476. [Google Scholar] [CrossRef] [Green Version]
- Rietveld, A.; van Kemenade, T.J.J.M.; Hak, T.; Verkleij, A.J.; de Kruijff, B. The effect of cytochrome-c-oxidase on lipid polymorphism of model membranes containing cardiolipin. Eur. J. Biochem. 1987, 164, 137–140. [Google Scholar] [CrossRef]
- Simidjiev, I.; Barzda, V.; Mustardy, L.; Garab, G. Role of thylakoid lipids in the structural flexibility of lamellar aggregates of the isolated light-harvesting chlorophyll a/b complex of photosystem II. Biochemistry 1998, 37, 4169–4173. [Google Scholar] [CrossRef] [PubMed]
- Nicolson, G.L. The Fluid-Mosaic Model of Membrane Structure: Still relevant to understanding the structure, function and dynamics of biological membranes after more than 40 years. Biochim. Biophys. Acta 2014, 1838, 1451–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Kruijff, B. Biomembranes—Lipids beyond the bilayer. Nature 1997, 386, 129–130. [Google Scholar] [CrossRef] [PubMed]
- van den Brink-van der Laan, E.; Killian, J.A.; de Kruijff, B. Nonbilayer lipids affect peripheral and integral membrane proteins via changes in the lateral pressure profile. Biochim. Biophys. Acta 2004, 1666, 275–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tietz, S.; Leuenberger, M.; Höhner, R.; Olson, A.H.; Fleming, G.R.; Kirchhoff, H. A proteoliposome-based system reveals how lipids control photosynthetic light harvesting. J. Biol. Chem. 2020. [Google Scholar] [CrossRef] [Green Version]
- Brown, M.F. Curvature Forces in Membrane Lipid-Protein Interactions. Biochemistry 2012, 51, 9782–9795. [Google Scholar] [CrossRef] [Green Version]
- Bagatolli, L.A.; Ipsen, J.H.; Simonsen, A.C.; Mouritsen, O.G. An outlook on organization of lipids in membranes: Searching for a realistic connection with the organization of biological membranes. Prog. Lipid Res. 2010, 49, 378–389. [Google Scholar] [CrossRef]
- Garab, G.; Lohner, K.; Laggner, P.; Farkas, T. Self-regulation of the lipid content of membranes by non-bilayer lipids: A hypothesis. Trends Plant. Sci. 2000, 5, 489–494. [Google Scholar] [CrossRef]
- Garab, G.; Ughy, B.; Goss, R. Role of MGDG and Non-bilayer Lipid Phases in the Structure and Dynamics of Chloroplast Thylakoid Membranes. In Lipids in Plant and Algae Development; Nakamura, Y., Li-Beisson, Y., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 127–157. [Google Scholar]
- Kirchhoff, H. Chloroplast ultrastructure in plants. New Phytol. 2019, 223, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Krumova, S.B.; Dijkema, C.; de Waard, P.; Van As, H.; Garab, G.; van Amerongen, H. Phase behaviour of phosphatidylglycerol in spinach thylakoid membranes as revealed by P-31-NMR. Biochim. Biophys. Acta 2008, 1778, 997–1003. [Google Scholar] [CrossRef] [Green Version]
- Cullis, P.R.; de Kruijff, B. Lipid polymorphism and the functional roles of lipids in biological-membranes. Biochim. Biophys. Acta 1979, 559, 399–420. [Google Scholar] [CrossRef]
- Schiller, J.; Muller, M.; Fuchs, B.; Arnold, K.; Huster, D. 31P NMR Spectroscopy of Phospholipids: From Micelles to Membranes. Curr. Anal. Chem. 2007, 3, 283–301. [Google Scholar] [CrossRef]
- Watts, A. NMR of Lipids. In Encyclopedia of Biophysics; Roberts, G.C.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1727–1738. [Google Scholar]
- Páli, T.; Garab, G.; Horváth, L.I.; Kóta, Z. Functional significance of the lipid-protein interface in photosynthetic membranes. Cell Mol. Life Sci. 2003, 60, 1591–1606. [Google Scholar] [CrossRef] [PubMed]
- Harańczyk, H.; Strzalka, K.; Bayerl, T.; Klose, G.; Blicharski, J.S. P-31 NMR measurements in photosynthetic membranes of wheat. Photosynthetica 1985, 19, 414–416. [Google Scholar]
- Harańczyk, H.; Strzalka, K.; Dietrich, W.; Blicharski, J.S. P-31-NMR observation of the temperature and glycerol induced non-lamellar phase-formation in wheat thylakoid membranes. J. Biol. Phys. 1995, 21, 125–139. [Google Scholar] [CrossRef]
- Duchene, S.; Siegenthaler, P.A. Do glycerolipids display lateral heterogeneity in the thylakoid membrane? Lipids 2000, 35, 739–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garab, G.; Ughy, B.; de Waard, P.; Akhtar, P.; Javornik, U.; Kotakis, C.; Šket, P.; Karlický, V.; Materová, Z.; Špunda, V.; et al. Lipid polymorphism in chloroplast thylakoid membranes—as revealed by P-31-NMR and timeresolved merocyanine fluorescence spectroscopy. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Krumova, S.B.; Koehorst, R.B.M.; Bóta, A.; Páli, T.; van Hoek, A.; Garab, G.; van Amerongen, H. Temperature dependence of the lipid packing in thylakoid membranes studied by time- and spectrally resolved fluorescence of Merocyanine 540. Biochim. Biophys. Acta 2008, 1778, 2823–2833. [Google Scholar] [CrossRef] [Green Version]
- Kotakis, C.; Akhtar, P.; Zsiros, O.; Garab, G.; Lambrev, P.H. Increased thermal stability of photosystem II and the macro-organization of thylakoid membranes, induced by co-solutes, associated with changes in the lipid-phase behaviour of thylakoid membranes. Photosynthetica 2018, 56, 254–264. [Google Scholar] [CrossRef]
- Ughy, B.; Karlický, V.; Dlouhý, O.; Javornik, U.; Materová, Z.; Zsiros, O.; Šket, P.; Plavec, J.; Špunda, V.; Garab, G. Lipid-polymorphism of plant thylakoid membranes. Enhanced non-bilayer lipid phases associated with increased membrane permeability. Physiol. Plant. 2019, 166, 278–287. [Google Scholar] [CrossRef]
- Dlouhý, O.; Kurasová, I.; Karlický, V.; Javornik, U.; Šket, P.; Petrova, N.Z.; Krumova, S.B.; Plavec, J.; Ughy, B.; Špunda, V.; et al. Modulation of non-bilayer lipid phases and the structure and functions of thylakoid membranes: Effects on the water-soluble enzyme violaxanthin de-epoxidase. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Mustárdy, L.; Buttle, K.; Steinbach, G.; Garab, G. The Three-Dimensional Network of the Thylakoid Membranes in Plants: Quasihelical Model of the Granum-Stroma Assembly. Plant. Cell 2008, 20, 2552–2557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bussi, Y.; Shimoni, E.; Weiner, A.; Kapon, R.; Charuvi, D.; Nevo, R.; Efrati, E.; Reich, Z. Fundamental helical geometry consolidates the plant photosynthetic membrane. Proc. Natl. Acad. Sci. USA 2019, 116, 22366–22375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimoni, E.; Rav-Hon, O.; Ohad, I.; Brumfeld, V.; Reich, Z. Three-dimensional organization of higher-plant chloroplast thylakoid membranes revealed by electron tomography. Plant. Cell 2005, 17, 2580–2586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, B.; Anderson, J.M. Lateral heterogenity in the distribution of chlorophyll-protein complexes of the thylakoid membranes of spinach-chloroplast. Biochim. Biophys. Acta 1980, 593, 427–440. [Google Scholar] [CrossRef]
- Anderson, J.M.; Boardman, N.K. Fractionation of photochemical systems of photosynthesis. I. Chlorophyll contents and photochemical activities of particles isolated from spinach chloroplasts. Biochim. Biophys. Acta 1966, 112, 403–421. [Google Scholar] [CrossRef]
- Peters, F.; van Spanning, R.; Kraayenhof, R. Studies on well coupled photosystem I-enriched sub-chloroplast vesicles—optimization of ferredoxin-mediated cyclic phosphorylation and electric-potential generation. Biochim. Biophys. Acta 1983, 724, 159–165. [Google Scholar] [CrossRef]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous-equations for assaying chlorophyll-a and chlorophyll-b extracted with 4 different solvents—verification of the concentration of chlorophyll standards by atomic-absorption spectroscopy. Biochim. Biophys. Acta 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Kublicki, M.; Koszelewski, D.; Brodzka, A.; Ostaszewski, R. Wheat germ lipase: Isolation, purification and applications. Crit. Rev. Biotechnol. 2021, 1–17. [Google Scholar] [CrossRef]
- Massiot, D.; Fayon, F.; Capron, M.; King, I.; Le Calvé, S.; Alonso, B.; Durand, J.O.; Bujoli, B.; Gan, Z.H.; Hoatson, G. Modelling one- and two-dimensional solid-state NMR spectra. Magn. Reson. Chem. 2002, 40, 70–76. [Google Scholar] [CrossRef]
- de Kruijff, B.; Morris, G.A.; Cullis, P.R. Application of P-31-NMR saturation transfer techniques to investigate phospholipid motion and organization in model and biological-membranes. Biochim. Biophys. Acta 1980, 598, 206–211. [Google Scholar] [CrossRef]
- Levitt, M.H. Spin Dynamics: Basics of Nuclear Magnetic Resonance; John Wiley & Sons: Hoboken, NJ, USA, 2008; chapter 15 (Motion and relaxation). [Google Scholar]
- Mustardy, L.; Garab, G. Granum revisited. A three-dimensional model—where things fall into place. Trends Plant. Sci. 2003, 8, 117–122. [Google Scholar] [CrossRef]
- Garab, G. Hierarchical organization and structural flexibility of thylakoid membranes. Biochim. Biophys. Acta 2014, 1837, 481–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, L.L.; Huang, Z.H.; Chang, S.H.; Wang, W.D.; Wang, J.F.; Kuang, T.Y.; Han, G.Y.; Shen, J.R.; Zhang, X. Structure of a C2S2M2N2 type PSII-LHCII supercomplex from the green alga Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2019, 116, 21246–21255. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.W.; Ma, J.; Su, X.D.; Cao, P.; Chang, W.R.; Liu, Z.F.; Zhang, X.Z.; Li, M. Structure of the maize photosystem I supercomplex with light-harvesting complexes I and II. Science 2018, 360, 1109–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasanov, S.E.; Kim, A.A.; Yaguzhinsky, L.S.; Dagda, R.K. Non-bilayer structures in mitochondrial membranes regulate ATP synthase activity. Biochim. Biophys. Acta 2018, 1860, 586–599. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
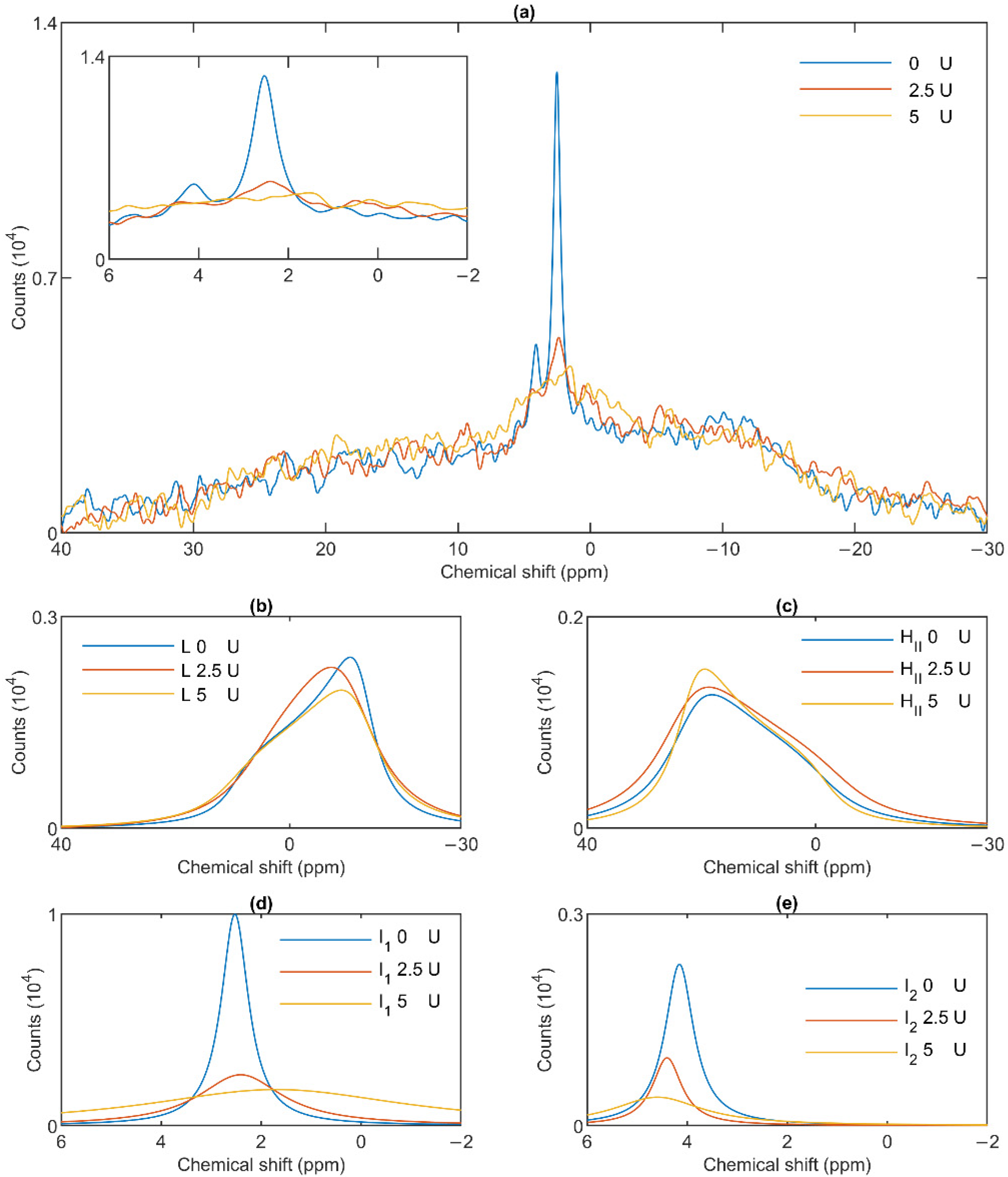{kind=link}
| Granum | Stroma | |||
|---|---|---|---|---|
| FWHM | Position (ppm) | FWHM | Position (ppm) | |
| I1 | 1.22 ± 0.16 | 2.66 ± 0.07 | 0.71 ± 0.10 | 2.68 ± 0.16 |
| I2 | 1.51 ± 0.96 | 3.98 ± 0.40 | 0.89 ± 0.13 | 4.19 ± 0.13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dlouhý, O.; Javornik, U.; Zsiros, O.; Šket, P.; Karlický, V.; Špunda, V.; Plavec, J.; Garab, G. Lipid Polymorphism of the Subchloroplast—Granum and Stroma Thylakoid Membrane—Particles. I. 31P-NMR Spectroscopy. Cells 2021, 10, 2354. https://doi.org/10.3390/cells10092354
Dlouhý O, Javornik U, Zsiros O, Šket P, Karlický V, Špunda V, Plavec J, Garab G. Lipid Polymorphism of the Subchloroplast—Granum and Stroma Thylakoid Membrane—Particles. I. 31P-NMR Spectroscopy. Cells. 2021; 10(9):2354. https://doi.org/10.3390/cells10092354
Chicago/Turabian StyleDlouhý, Ondřej, Uroš Javornik, Ottó Zsiros, Primož Šket, Václav Karlický, Vladimír Špunda, Janez Plavec, and Győző Garab. 2021. "Lipid Polymorphism of the Subchloroplast—Granum and Stroma Thylakoid Membrane—Particles. I. 31P-NMR Spectroscopy" Cells 10, no. 9: 2354. https://doi.org/10.3390/cells10092354
APA StyleDlouhý, O., Javornik, U., Zsiros, O., Šket, P., Karlický, V., Špunda, V., Plavec, J., & Garab, G. (2021). Lipid Polymorphism of the Subchloroplast—Granum and Stroma Thylakoid Membrane—Particles. I. 31P-NMR Spectroscopy. Cells, 10(9), 2354. https://doi.org/10.3390/cells10092354