
Potential Therapeutic Effect of Micrornas in Extracellular Vesicles from Mesenchymal Stem Cells against SARS-CoV-2

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. EV Isolation and Nanoparticle Tracking Analysis (NTA)
2.3. EVs Staining
2.4. Western Blotting
2.5. Immunocytochemistry (ICC)
2.6. RNA Extraction and Quantitative PCR (qPCR)
2.7. Antiviral Activity Assay
2.8. Indirect Immunofluorescence Assay
2.9. Transfection and Reporter Assay
2.10. RNA Sequencing and Data Processing
2.11. MiRNA Target Prediction and Functional Analysis of Binding Sites
2.12. Statistics
3. Results
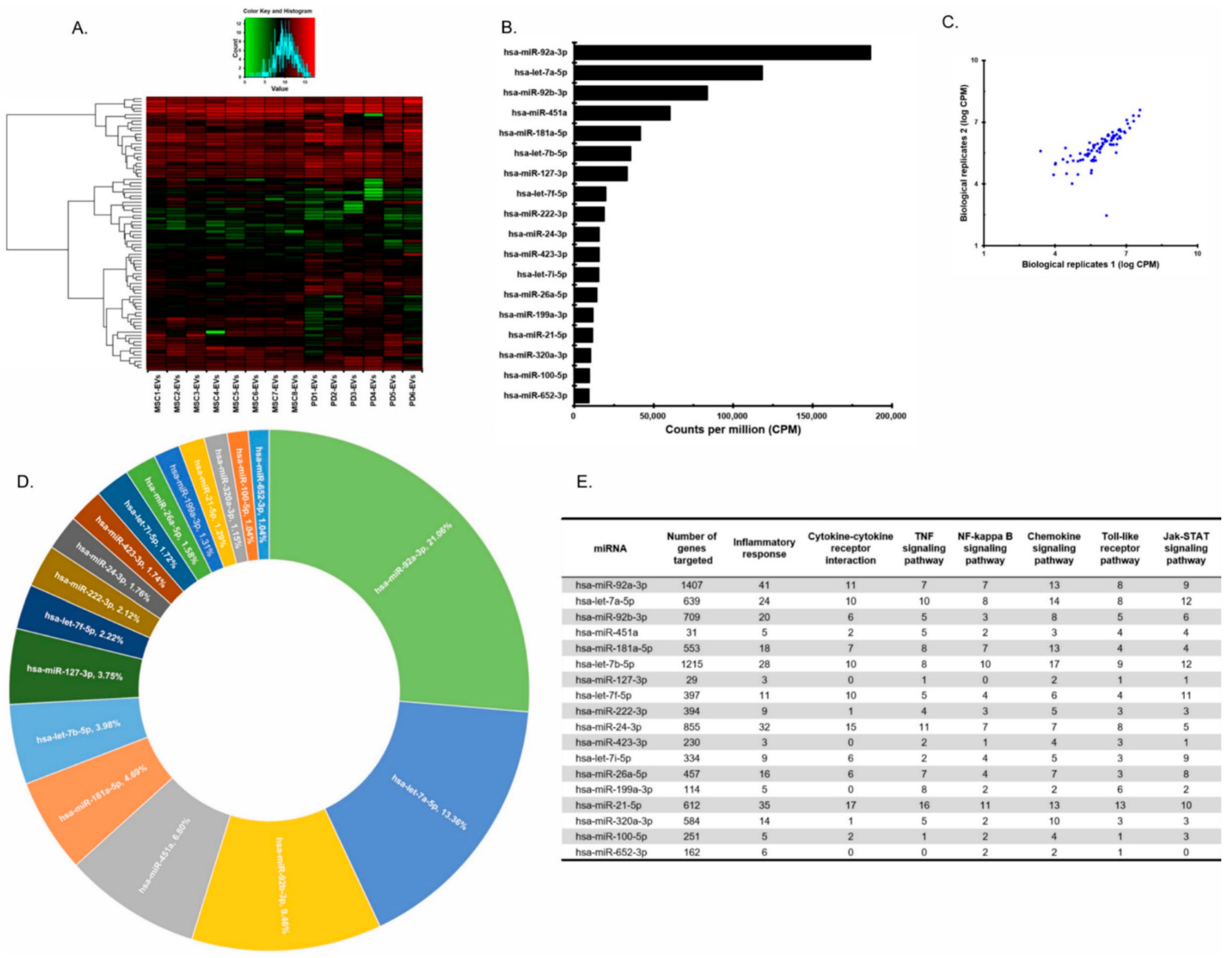
3.1. Profiles of MiRNAs of pMSC-EVs and Placenta EVs
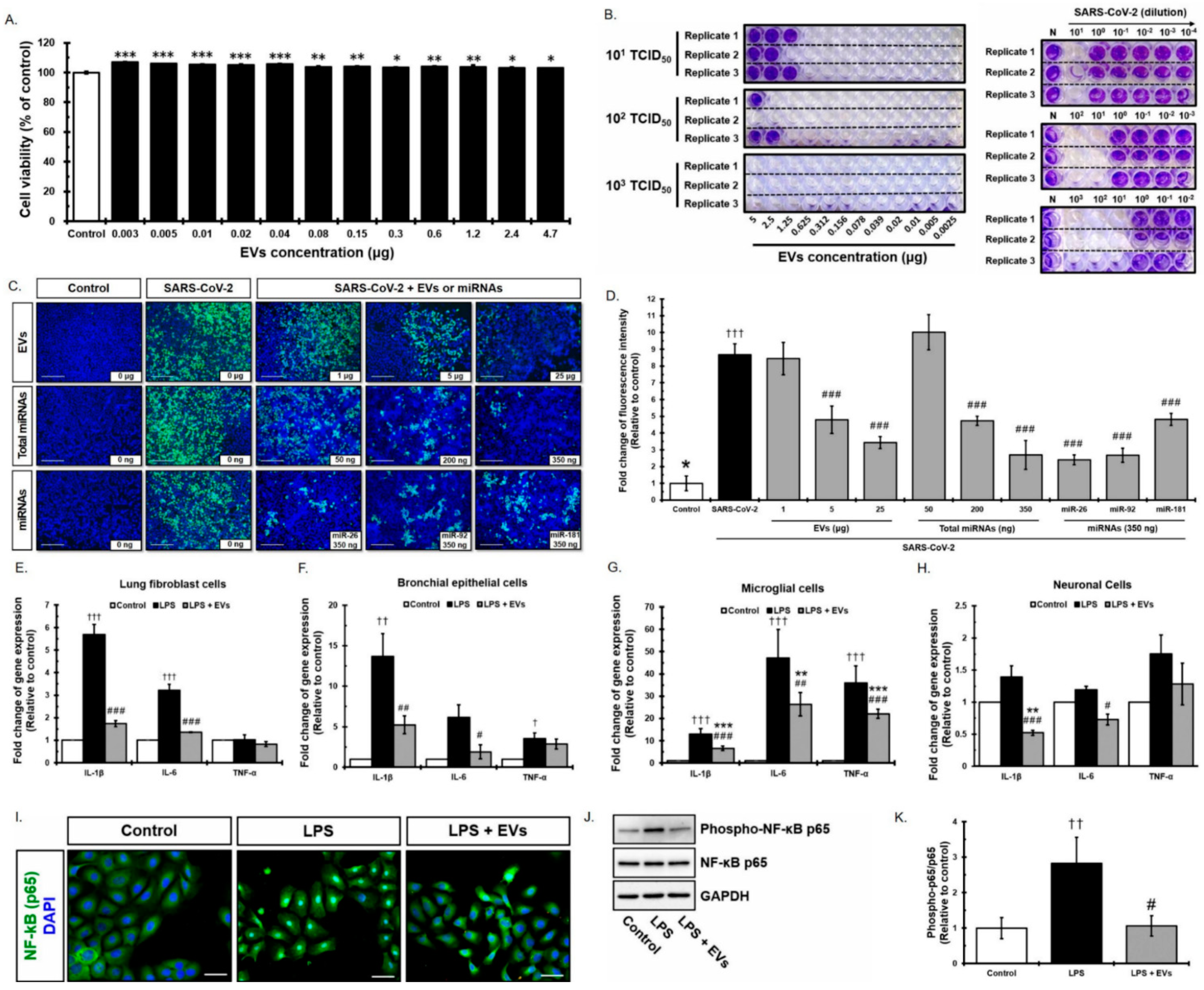
3.2. Antiviral Effect of Placenta-Derived EVs against SARS-CoV-2
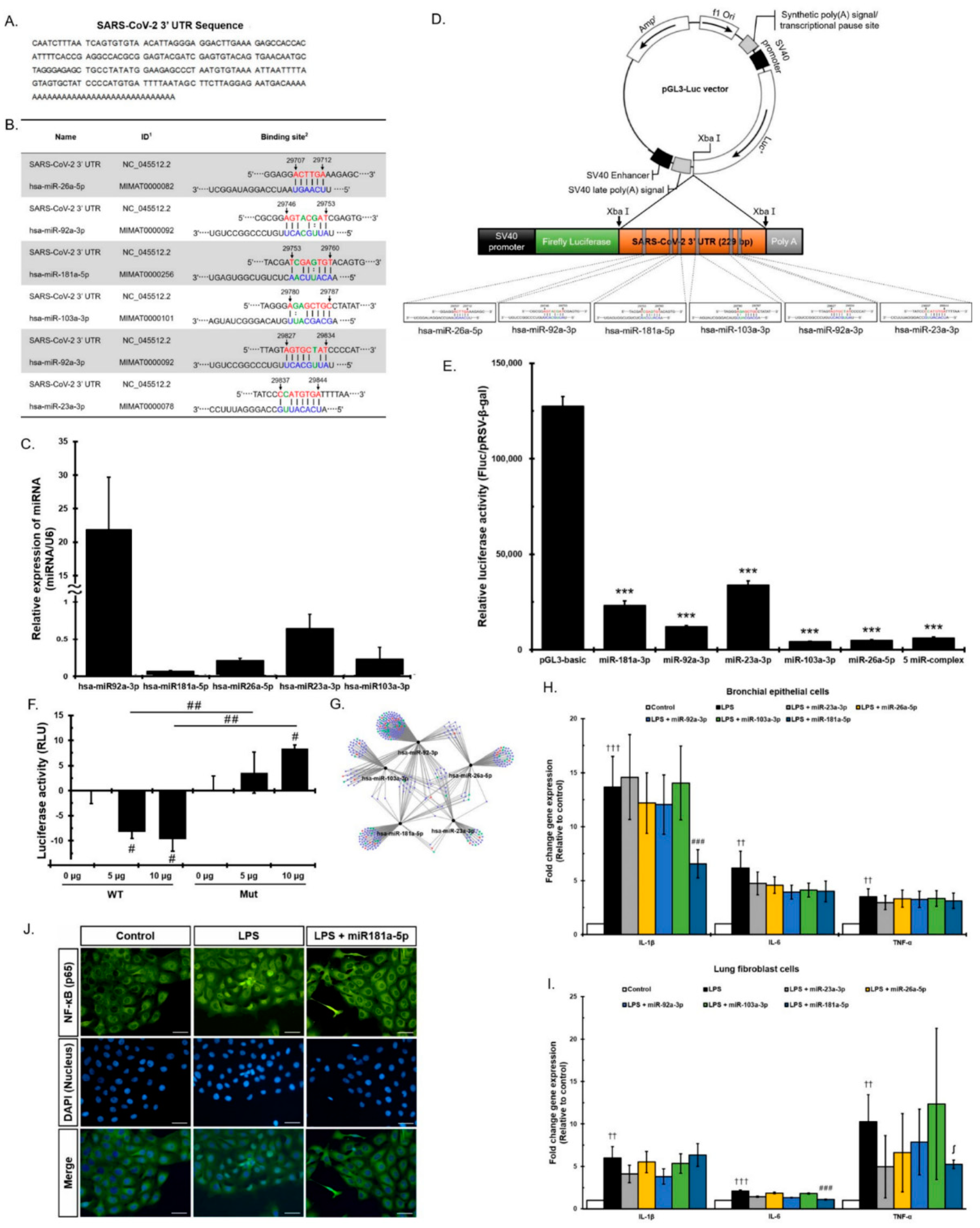
3.3. Direct Viral Effect of MiRNAs in EVs on SARS-CoV-2
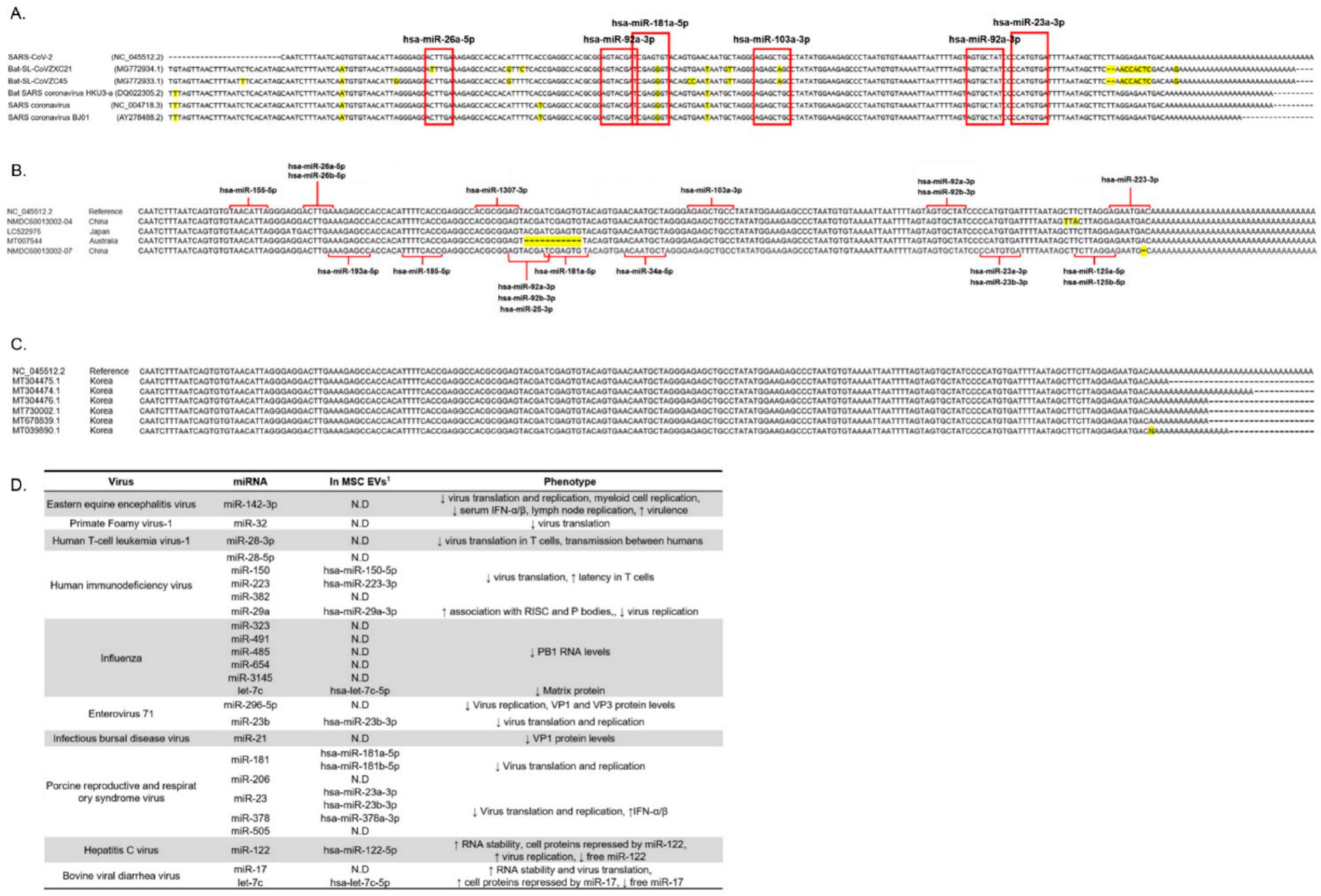
3.4. The 3′ UTR Region of Several Coronaviruses Is Highly Conserved Even in Recent SARS-CoV-2 Mutations
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bernard, N.F.; David, M.K.; Peter, M.H. Fields Virology, 5th ed.; Wolters Kluwer Health/Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007. [Google Scholar]
- Lu, J.; Clark, A.G. Impact of microRNA regulation on variation in human gene expression. Genome Res. 2012, 22, 1243–1254. [Google Scholar] [CrossRef] [Green Version]
- Trobaugh, D.W.; Klimstra, W.B. MicroRNA Regulation of RNA Virus Replication and Pathogenesis. Trends Mol. Med. 2017, 23, 80–93. [Google Scholar] [CrossRef]
- Ameres, S.L.; Zamore, P.D. Diversifying microRNA sequence and function. Nat. Rev. Mol. Cell Biol. 2013, 14, 475–488. [Google Scholar] [CrossRef]
- Eichhorn, S.W.; Guo, H.; Mcgeary, S.E.; Rodriguez-Mias, R.A.; Shin, C.; Baek, D.; Hsu, S.H.; Ghoshal, K.; Villen, J.; Bartel, D.P. mRNA destabilization is the dominant effect of mammalian microRNAs by the time substantial repression ensues. Mol. Cell 2014, 56, 104–115. [Google Scholar] [CrossRef] [Green Version]
- Brian, L.H.; Olga, A.M.; Dzung, C.T.; James, M.S.; Alexander, G.P. MicroRNA targeting of neurotropic flavivirus: Effective control of virus escape and reversion to neurovirulent phenotype. J. Virol. 2012, 86, 5647–5659. [Google Scholar] [CrossRef] [Green Version]
- Derek, W.T.; Christina, L.G.; Chengqun, S.; Andrew, D.H.; Eryu, W.; Elik, C.; Alexander, M.; Scott, C.W.; Kate, D.R.; William, B.K. RNA viruses can hijack vertebrate microRNAs to suppress innate immunity. Nature 2014, 506, 245–248. [Google Scholar] [CrossRef] [Green Version]
- Theo, W.D. Functions of the 3′-untranslated regions of positive strand RNA viral genomes. Annu. Rev. Phytopathol. 1999, 37, 151–174. [Google Scholar] [CrossRef]
- Dong, Y.; Julian, L.L. The structure and functions of coronavirus genomic 3’ and 5’ ends. Virus Res. 2015, 206, 120–133. [Google Scholar] [CrossRef]
- Yanez-Mo, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borras, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosaka, N.; Igughi, H.; Yoshioka, Y.; Takeshita, F.; Matsuki, Y.; Ochiya, T. Secretory mechanisms and intercellular transfer of microRNAs in living cells. J. Biol. Chem. 2010, 285, 17442–17452. [Google Scholar] [CrossRef] [Green Version]
- Okoye, I.S.; Coomes, S.M.; Pelly, V.S.; Czieso, S.; Papayannopoulos, V.; Tolmachova, T.; Seabra, M.C.; Wilson, M.S. MicroRNA-containing T-regulatory-cell-derived exosomes suppress pathogenic T helper 1 cells. Immunity 2014, 41, 89–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, K.S.; Bandeira, E.; Shelke, G.V.; Lasser, C.; Lotvall, J. Enhancement of therapeutic potential of mesenchymal stem cell-derived extracellular vesicles. Stem Cell Res. Ther. 2019, 10, 288. [Google Scholar] [CrossRef]
- Pasquale, M.; Valeria, P.; Silvia, Z.; Azzurra, S.; Daniele, G.; Barbara, R.; Laura, B.; Francesco, A. Microfluidic Tools for Enhanced Characterization of Therapeutic Stem Cells and Prediction of Their Potential Antimicrobial Secretome. Antibiotics 2021, 10, 750. [Google Scholar] [CrossRef]
- Barile, L.; Lionetti, V.; Cervio, E.; Matteucci, M.; Gherghiceanu, M.; Popescu, L.M.; Torre, T.; Siclari, F.; Moccetti, T.; Vassalli, G. Extracellular vesicles from human cardiac progenitor cells inhibit cardiomyocyte apoptosis and improve cardiac function after myocardial infarction. Cardiovasc. Res. 2014, 103, 530–541. [Google Scholar] [CrossRef] [PubMed]
- Pocsfalvi, G.; Mammadova, R.; Ramos Juarez, A.P.; Bokka, R.; Trepiccione, F.; Capasso, G. COVID-19 and Extracellular Vesicles: An Intriguing Interplay. Kidney Blood Press. Res. 2020, 45, 661–670. [Google Scholar] [CrossRef]
- Moon, J.; Schwarz, S.C.; Lee, H.S.; Kang, J.M.; Lee, Y.E.; Kim, B.; Sung, M.Y.; Hoglinger, G.; Wegner, F.; Kim, J.S.; et al. Preclinical Analysis of Fetal Human Mesencephalic Neural Progenitor Cell Lines: Characterization and Safety In Vitro and In Vivo. Stem Cells Transl. Med. 2017, 6, 576–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. The Subread aligner: Fast, accurate and scalable read mapping by seed-and-vote. Nucleic Acids Res. 2013, 41, e108. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Aparicio-Puerta, E.; Lebron, R.; Rueda, A.; Gomez-Martin, C.; Giannoukakos, S.; Jaspez, D.; Medina, J.M.; Zubkovic, A.; Jurak, I.; Fromm, B.; et al. sRNAbench and sRNAtoolbox 2019: Intuitive fast small RNA profiling and differential expression. Nucleic Acids Res. 2019, 47, W530–W535. [Google Scholar] [CrossRef] [Green Version]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Proteomics: Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Kertesz, M.; Iovino, N.; Unnerstall, U.; Gaul, U.; Segal, E. The role of site accessibility in microRNA target recognition. Nat. Genet. 2007, 39, 1278–1284. [Google Scholar] [CrossRef]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wang, X. miRDB: An online database for prediction of functional microRNA targets. Nucleic Acids Res. 2020, 48, D127–D131. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.Y.; Lin, Y.C.; Li, J.; Huang, K.Y.; Shrestha, S.; Hong, H.C.; Tang, Y.; Chen, Y.G.; Jin, C.N.; Yu, Y.; et al. miRTarBase 2020: Updates to the experimentally validated microRNA-target interaction database. Nucleic Acids Res. 2020, 48, D148–D154. [Google Scholar] [CrossRef] [Green Version]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Luo, W.; Brouwer, C. Pathview: An R/Bioconductor package for pathway-based data integration and visualization. Bioinformatics 2013, 29, 1830–1831. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Kikuguchi, C.; Nishijima, S.; Nagashima, T.; Takahashi, A.; Okada, M.; Yamamoto, T. Postnatal liver functional maturation requires Cnot complex-mediated decay of mRNAs encoding cell cycle and immature liver genes. Development 2019, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benali-Furet, N.L.; Chami, M.; Houel, L.; De Giorgi, F.; Vernejoul, F.; Lagorce, D.; Buscail, L.; Bartenschlager, R.; Ichas, F.; Rizzuto, R.; et al. Hepatitis C virus core triggers apoptosis in liver cells by inducing ER stress and ER calcium depletion. Oncogene 2005, 24, 4921–4933. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Ren, A.; Wang, X.; Fan, X.; Zhao, Y.; Gao, G.F.; Cleary, P.; Wang, B. Influenza viral neuraminidase primes bacterial coinfection through TGF-beta-mediated expression of host cell receptors. Proc. Natl. Acad. Sci. USA 2015, 112, 238–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, P.; Bailey-Bucktrout, S.; Xi, Y.; Xu, D.; Du, D.; Zhang, Q.; Xiang, W.; Liu, J.; Melton, A.; Sheppard, D.; et al. Innate antiviral host defense attenuates TGF-beta function through IRF3-mediated suppression of Smad signaling. Mol. Cell 2014, 56, 723–737. [Google Scholar] [CrossRef] [Green Version]
- Khailany, R.A.; Safdar, M.; Ozaslan, M. Genomic characterization of a novel SARS-CoV-2. Gene Rep. 2020, 19, 100682. [Google Scholar] [CrossRef]
- Hausser, J.; Syed, A.P.; Bilen, B.; Zavolan, M. Analysis of CDS-located miRNA target sites suggests that they can effectively inhibit translation. Genome Res. 2013, 23, 604–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.L.; Grey, F.E.; Uhrlaub, J.L.; Nikolich-Zugich, J.; Hirsch, A.J. Induction of the cellular microRNA, Hs_154, by West Nile virus contributes to virus-mediated apoptosis through repression of antiapoptotic factors. J. Virol. 2012, 86, 5278–5287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krutzfeldt, J.; Rajewsky, N.; Braich, R.; Rajeev, K.G.; Tuschl, T.; Manoharan, M.; Stoffel, M. Silencing of microRNAs in vivo with ‘antagomirs’. Nature 2005, 438, 685–689. [Google Scholar] [CrossRef]
- Rupaimoole, R.; Han, H.D.; Lopez-Berestein, G.; Sood, A.K. MicroRNA therapeutics: Principles, expectations, and challenges. Chin. J. Cancer 2011, 30, 368–370. [Google Scholar] [CrossRef] [PubMed]
- Grein, J.; Ohmagari, N.; Shin, D.; Diaz, G.; Asperges, E.; Castagna, A.; Feldt, T.; Green, G.; Green, M.L.; Lescure, F.X.; et al. Compassionate Use of Remdesivir for Patients with Severe Covid-19. N. Engl. J. Med. 2020, 382, 2327–2336. [Google Scholar] [CrossRef] [PubMed]
- Mohammadipoor, A.; Antebi, B.; Batchinsky, A.I.; Cancio, L.C. Therapeutic potential of products derived from mesenchymal stem/stromal cells in pulmonary disease. Respir. Res. 2018, 19, 218. [Google Scholar] [CrossRef]
- Gomzikova, M.O.; James, V.; Rizvanov, A.A. Therapeutic Application of Mesenchymal Stem Cells Derived Extracellular Vesicles for Immunomodulation. Front. Immunol. 2019, 10, 2663. [Google Scholar] [CrossRef]
- Fujita, Y.; Yoshioka, Y.; Ito, S.; Araya, J.; Kuwano, K.; Ochiya, T. Intercellular communication by extracellular vesicles and their microRNAs in asthma. Clin. Ther. 2014, 36, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Reiner, A.T.; Witwer, K.W.; Van Balkom, B.W.M.; De Beer, J.; Brodie, C.; Corteling, R.L.; Gabrielsson, S.; Gimona, M.; Ibrahim, A.G.; De Kleijn, D.; et al. Concise Review: Developing Best-Practice Models for the Therapeutic Use of Extracellular Vesicles. Stem Cells Transl. Med. 2017, 6, 1730–1739. [Google Scholar] [CrossRef] [Green Version]
- Harrell, C.R.; Jovicic, N.; Djonov, V.; Arsenijevic, N.; Volarevic, V. Mesenchymal Stem Cell-Derived Exosomes and Other Extracellular Vesicles as New Remedies in the Therapy of Inflammatory Diseases. Cells 2019, 8, 1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khatri, M.; Richardson, L.A.; Meulia, T. Mesenchymal stem cell-derived extracellular vesicles attenuate influenza virus-induced acute lung injury in a pig model. Stem Cell Res. Ther. 2018, 9, 17. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.C.; Lai, L.C. The potential roles of stem cell-derived extracellular vesicles as a therapeutic tool. Ann. Transl. Med. 2019, 7, 693. [Google Scholar] [CrossRef]
- Seo, Y.; Kim, H.S.; Hong, I.S. Stem Cell-Derived Extracellular Vesicles as Immunomodulatory Therapeutics. Stem Cells Int. 2019, 5126156. [Google Scholar] [CrossRef]
- Xie, M.; Xiong, W.; She, Z.; Wen, Z.; Abdirahman, A.S.; Wan, W.; Wen, C. Immunoregulatory Effects of Stem Cell-Derived Extracellular Vesicles on Immune Cells. Front. Immunol. 2020, 11, 13. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
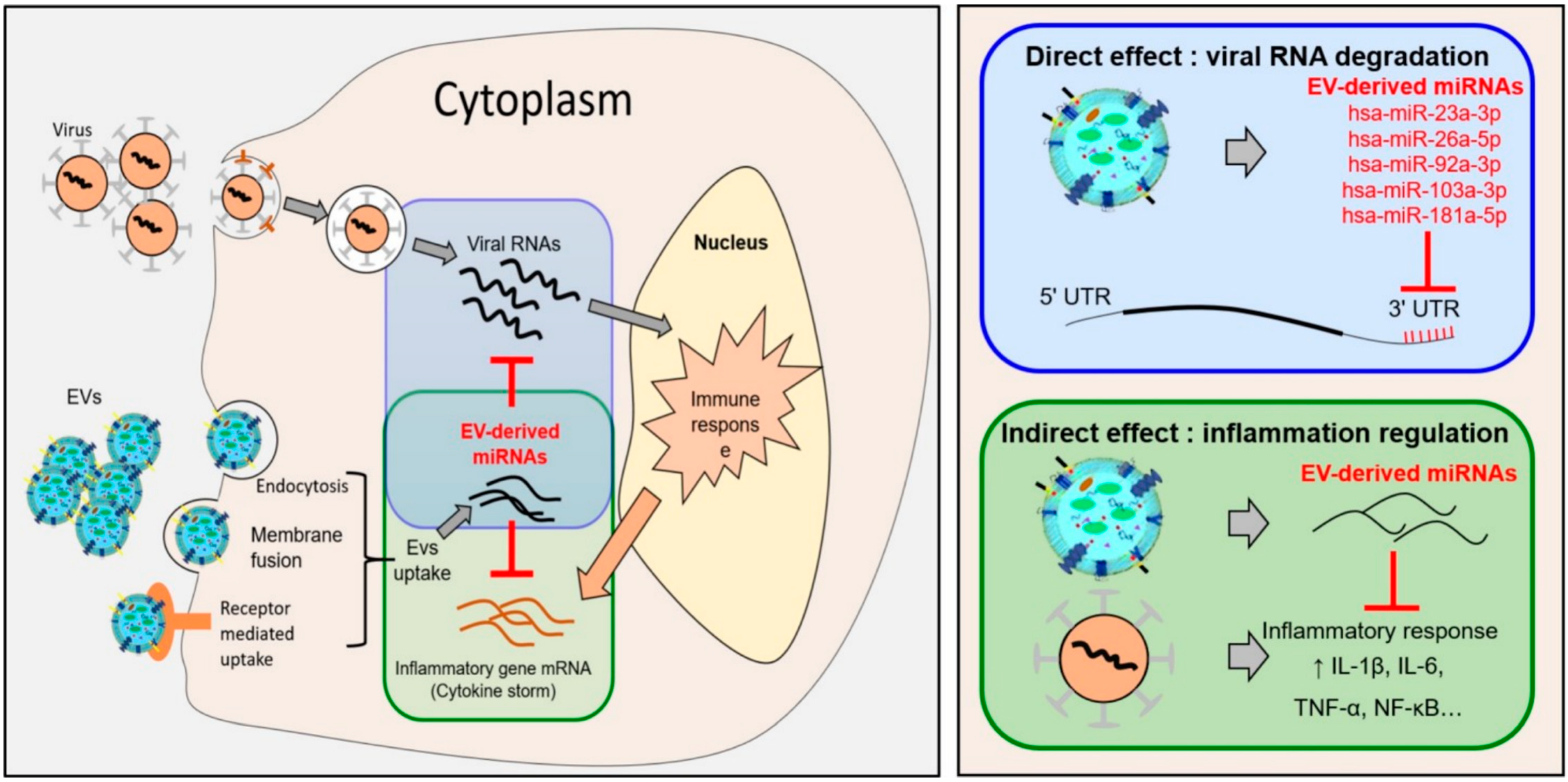{kind=link}
| PITA 3’UTR Binding Prediction 1 | mirDB Unconventional Target Sites Prediction 2 | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| microRNA | Expression (%) 3 | Seed Location 4 | Seed Match Length 5 | Mismatch 6 | G:U Wobble 7 | microRNA-Target Hybridization Energy 8 | Score 9 | Seed Location 10 | Rank 11 |
| hsa-miR-92a-3p | 22.23476 | 29,746 | 8 | 1 | 1 | −13.8 | − | − | 1 |
| hsa-miR-92b-3p | 9.978815 | 29,746 | 8 | 1 | 1 | −18.8 | − | − | 3 |
| hsa-miR-181a-5p | 4.92776 | 29,753 | 8 | 1 | 1 | −18.7 | 70 | 7410, 7529, 8221, 9016, 11,400, 12,216, 18,516, 20,783, 27,948 | 5 |
| hsa-miR-26a-5p | 1.672265 | 29,707 | 6 | 0 | 0 | −14.9 | 68 | 454, 9596, 20,513, 27,848, 29,707 | 12 |
| hsa-miR-34a-5p | 0.925787 | 29,768 | 8 | 1 | 1 | −13.25 | − | − | 20 |
| hsa-miR-23a-3p | 0.790515 | 29,837 | 8 | 1 | 0 | −8.1 | 79 | 6458, 7908, 15,302, 21,244 | 23 |
| hsa-miR-125b-5p | 0.389675 | 29,856 | 8 | 1 | 1 | −11.9 | − | − | 36 |
| hsa-miR-125a-5p | 0.324866 | 29,856 | 8 | 1 | 1 | −10.4 | − | − | 37 |
| hsa-miR-103a-3p | 0.269252 | 29,780 | 8 | 1 | 1 | −12.6 | 85 | 8827, 13,089, 14,561, 14,780, 22,345, 24,235, 25,319, 26,371, 27,101, 28,734, 28,920, 29,461 | 39 |
| hsa-miR-223-3p | 0.233216 | 29,863 | 8 | 1 | 1 | −5.6 | − | − | 45 |
| hsa-miR-25-3p | 0.153562 | 29,746 | 8 | 1 | 1 | −15.5 | − | − | 51 |
| hsa-miR-26b-5p | 0.149508 | 29,707 | 6 | 0 | 0 | −11.3 | 68 | 454, 9596, 20,513, 27,848, 29,707 | 52 |
| hsa-miR-193a-5p | 0.130145 | 29,712 | 8 | 1 | 0 | −12.1 | − | − | 57 |
| hsa-miR-1307-3p | 0.129069 | 29,740 | 8 | 1 | 0 | −30.9 | − | − | 58 |
| hsa-miR-155-5p | 0.098556 | 29,693 | 8 | 1 | 0 | −8.3 | 51 | 864, 5209, 17,197, 25,074 | 63 |
| hsa-miR-185-5p | 0.050131 | 29,726 | 8 | 1 | 1 | −11.31 | − | − | 69 |
| hsa-miR-23b-3p | 0.02939 | 29,837 | 8 | 1 | 0 | −11.3 | 79 | 6458, 7908, 15,302, 21,244 | 74 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.H.; Choi, Y.; Lim, C.-W.; Park, J.-M.; Yu, S.-H.; Kim, Y.; Han, H.J.; Kim, C.-H.; Song, Y.-S.; Kim, C.; et al. Potential Therapeutic Effect of Micrornas in Extracellular Vesicles from Mesenchymal Stem Cells against SARS-CoV-2. Cells 2021, 10, 2393. https://doi.org/10.3390/cells10092393
Park JH, Choi Y, Lim C-W, Park J-M, Yu S-H, Kim Y, Han HJ, Kim C-H, Song Y-S, Kim C, et al. Potential Therapeutic Effect of Micrornas in Extracellular Vesicles from Mesenchymal Stem Cells against SARS-CoV-2. Cells. 2021; 10(9):2393. https://doi.org/10.3390/cells10092393
Chicago/Turabian StylePark, Jae Hyun, Yuri Choi, Chul-Woo Lim, Ji-Min Park, Shin-Hye Yu, Yujin Kim, Hae Jung Han, Chun-Hyung Kim, Young-Sook Song, Chul Kim, and et al. 2021. "Potential Therapeutic Effect of Micrornas in Extracellular Vesicles from Mesenchymal Stem Cells against SARS-CoV-2" Cells 10, no. 9: 2393. https://doi.org/10.3390/cells10092393
APA StylePark, J. H., Choi, Y., Lim, C.-W., Park, J.-M., Yu, S.-H., Kim, Y., Han, H. J., Kim, C.-H., Song, Y.-S., Kim, C., Yu, S. R., Oh, E. Y., Lee, S.-M., & Moon, J. (2021). Potential Therapeutic Effect of Micrornas in Extracellular Vesicles from Mesenchymal Stem Cells against SARS-CoV-2. Cells, 10(9), 2393. https://doi.org/10.3390/cells10092393