
The Protection of Zinc against Acute Cadmium Exposure: A Morphological and Molecular Study on a BBB In Vitro Model

 , , ,
, , ,  , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Treatments
2.2. Zn Treatments
2.3. MTT Assay
2.4. Flow Cytometry
2.5. Superoxide Dismutase (SOD) Enzymatic Activity
2.6. Western Blotting
2.7. Immunofluorescence Staining
2.8. Statistical Analysis
3. Results
3.1. Zinc Prevents the Cd-Dependent Reduction in Cell Viability
3.2. Reactive Oxygen Species (ROS) Quantification
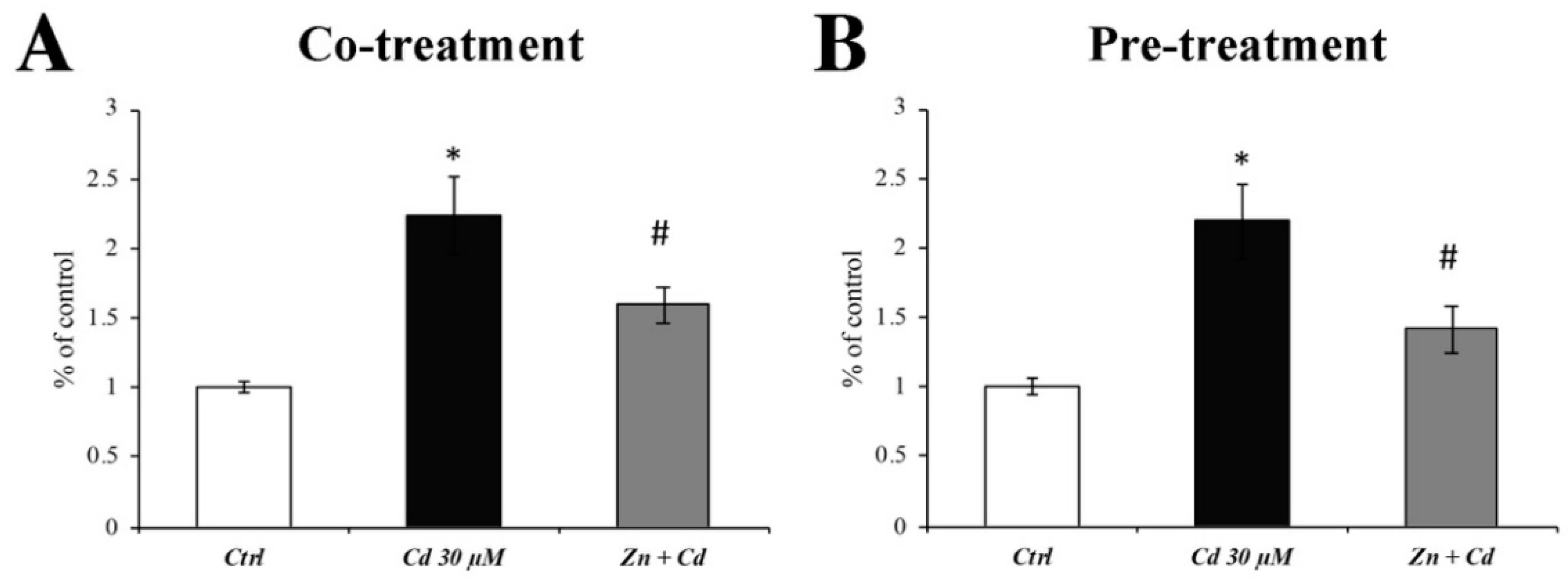
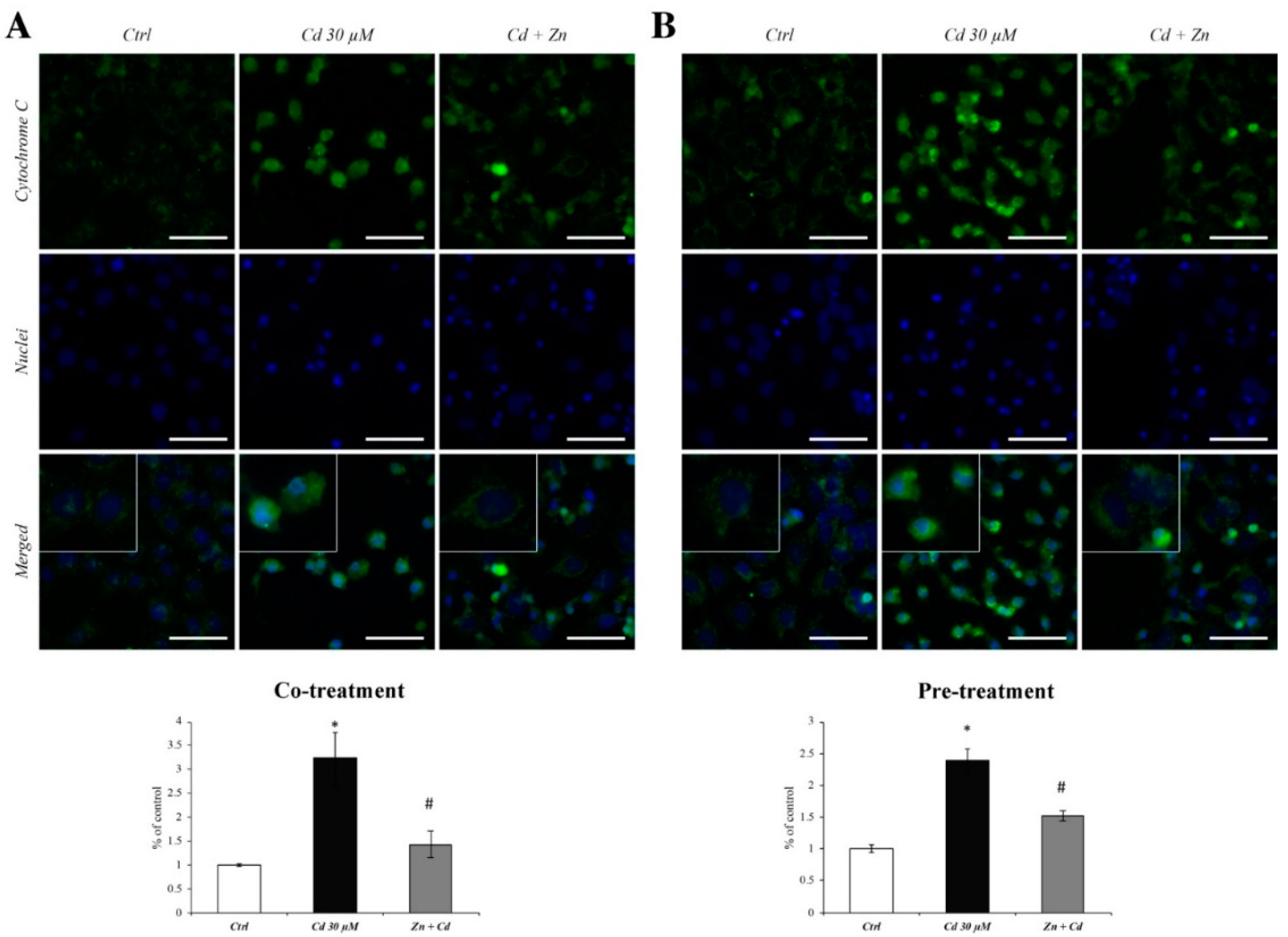
3.3. Zn Prevents the Cytochrome C Spillage from Mitochondria
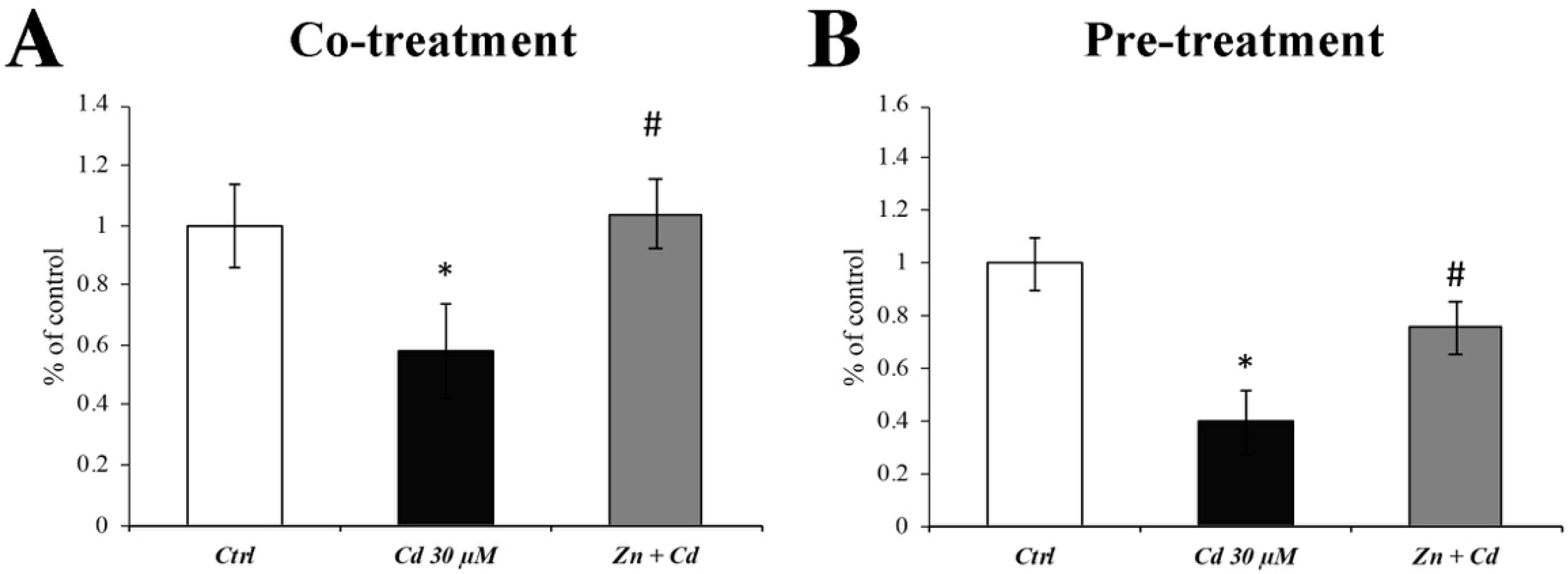
3.4. Zn Prevents the Cd-Dependent SOD Activity Decrease
3.5. Cd-Induced ER Stress Is Prevented by Zn Administration
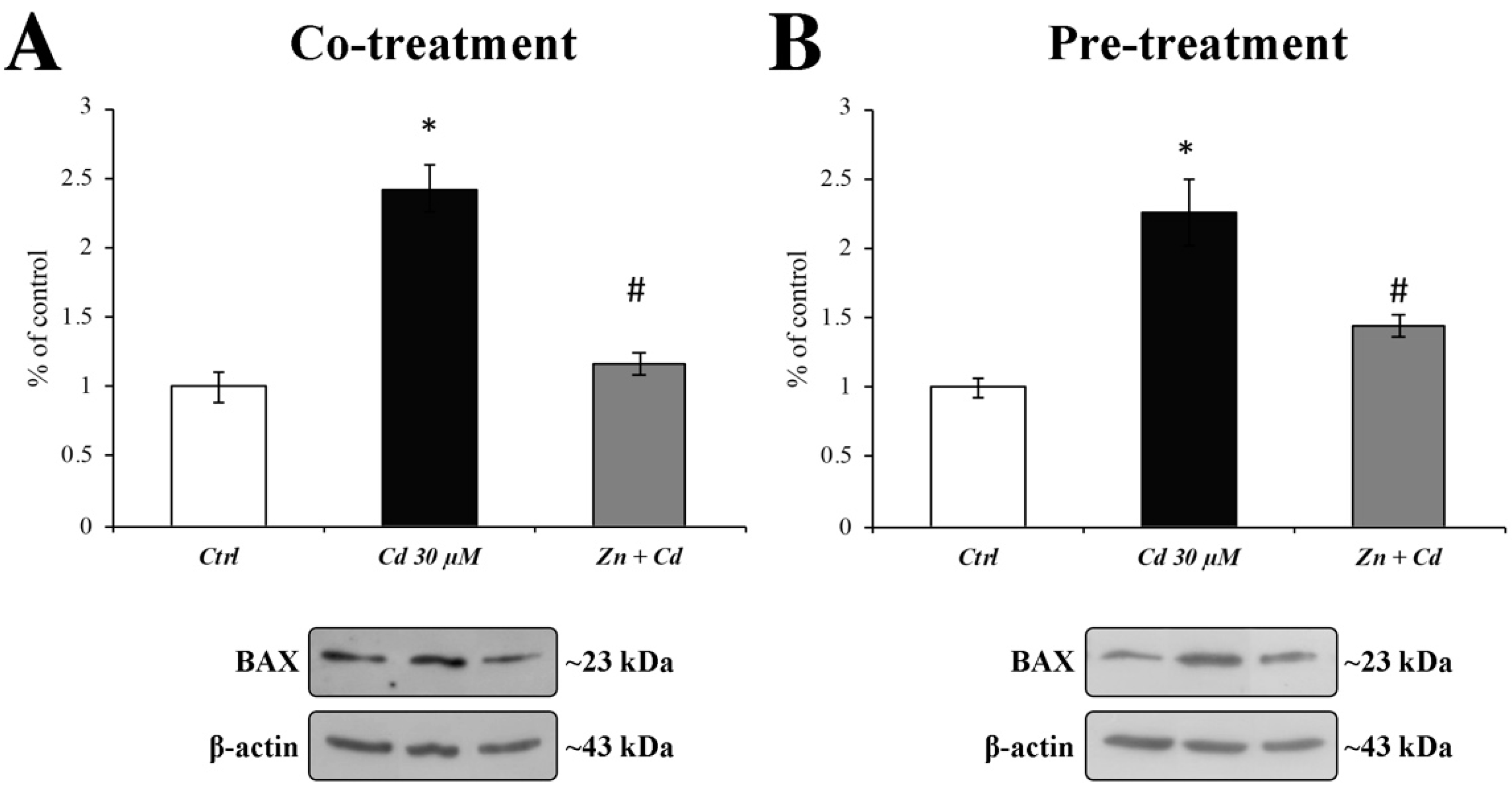
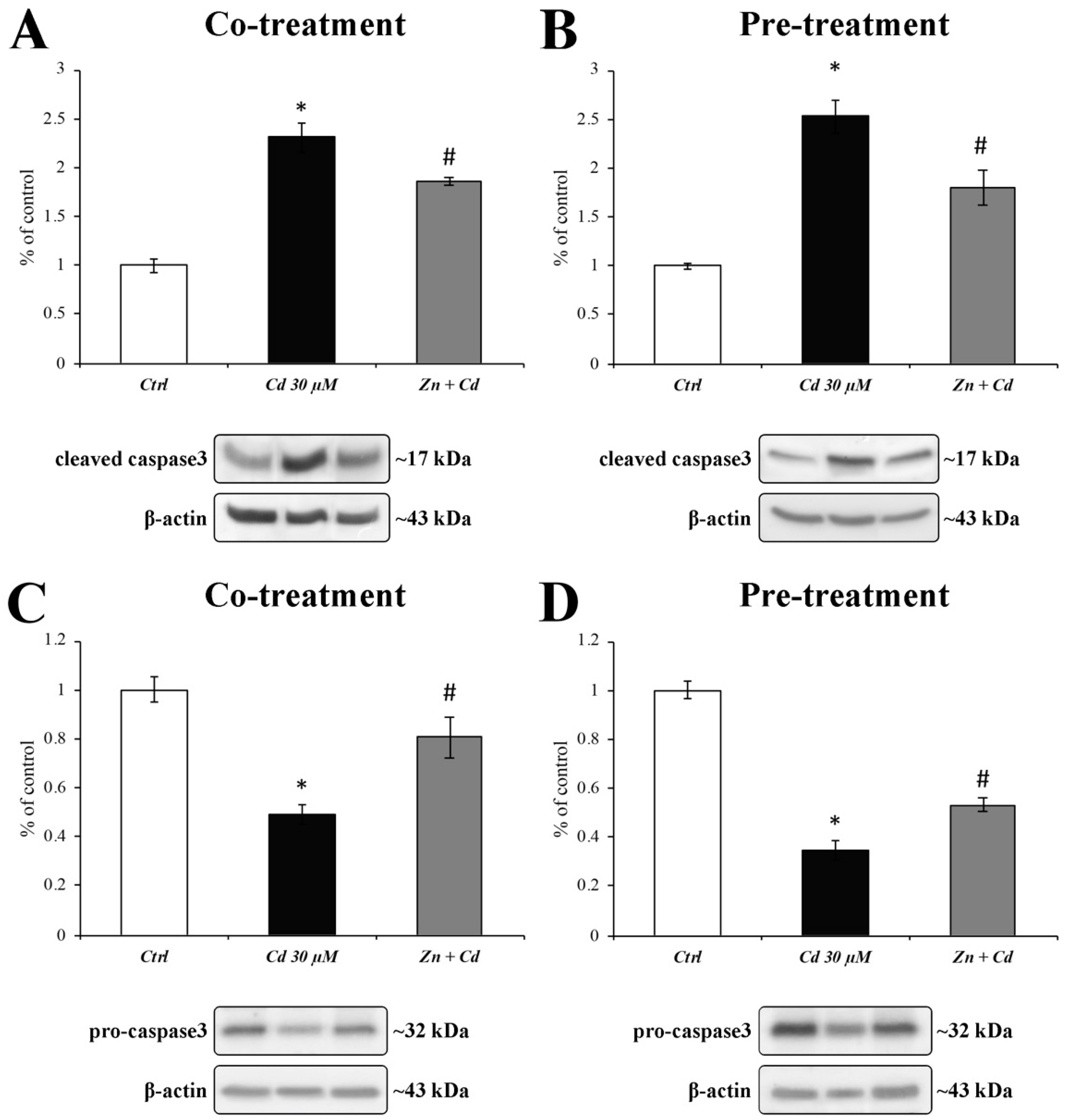
3.6. Zn Is Able to Prevent BAX Overexpression and Caspase-3 Activation
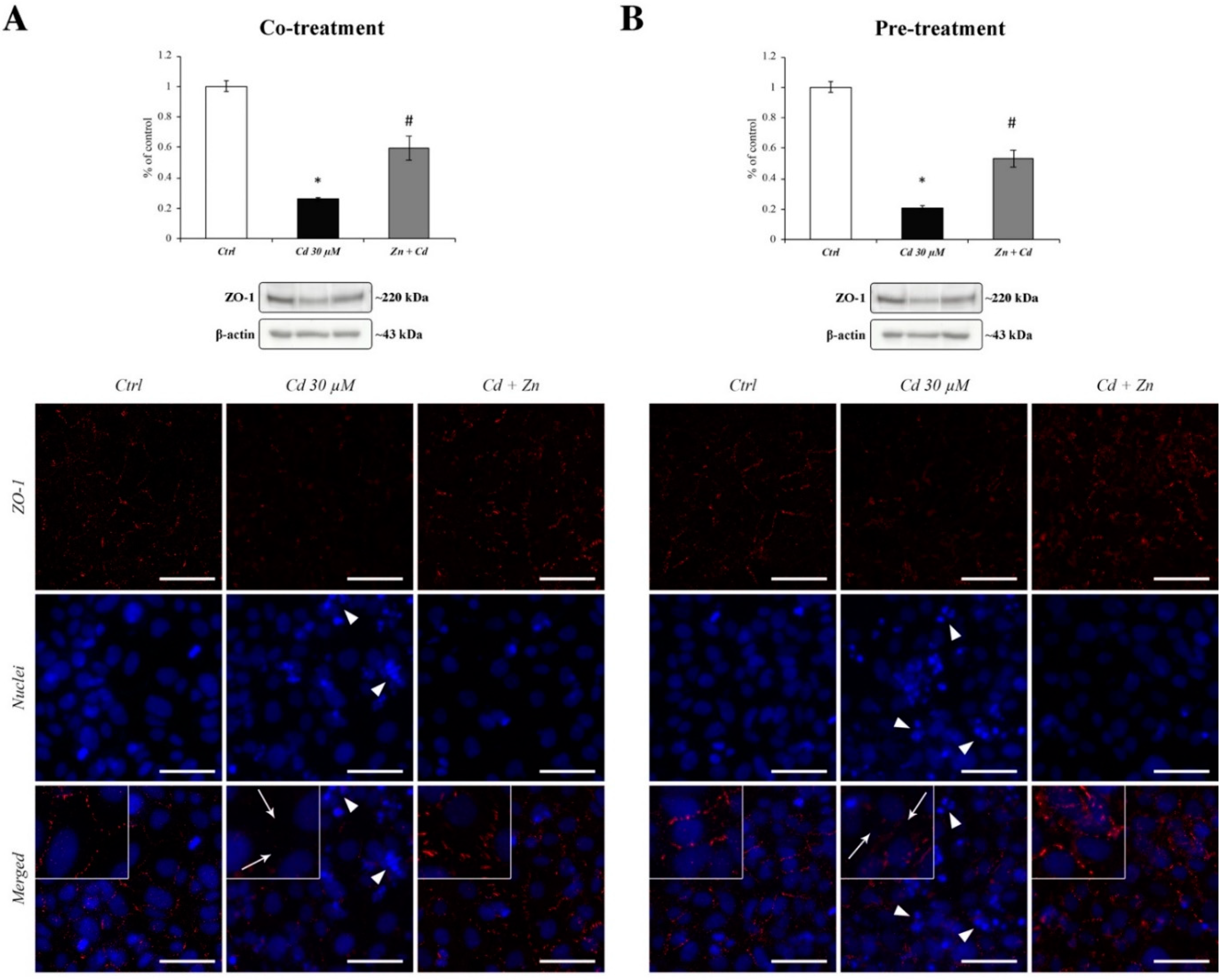
3.7. Zn-Dependent Protection of ZO-1 Tight Junction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy metal toxicity and the environment. In Molecular, Clinical and Environmental Toxicology; Experientia Supplementum; Luch, A., Ed.; Springer: Basel, Switzerland; Berlin/Heidelberg, Germany, 2012; Volume 101, pp. 133–164. ISBN 978-3-7643-8339-8. [Google Scholar]
- Nawrot, T.S.; Staessen, J.A.; Roels, H.A.; Munters, E.; Cuypers, A.; Richart, T.; Ruttens, A.; Smeets, K.; Clijsters, H.; Vangronsveld, J. Cadmium exposure in the population: From health risks to strategies of prevention. Biometals 2010, 23, 769–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, K.; Yu, Y.; Zhang, M.; Kim, K. Concentration, source, and total health risks of cadmium in multiple media in Densely Populated Areas, China. Int. J. Environ. Res. Public Health 2019, 16, 2269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Kim, Y.H.; An, H.C.; Sung, J.H.; Sim, C.S. Levels of blood lead and urinary cadmium in industrial complex residents in Ulsan. Ann. Occup. Environ. Med. 2017, 29, 26. [Google Scholar] [CrossRef] [PubMed]
- Naka, K.S.; de Cássia dos Santos Mendes, L.; de Queiroz, T.K.L.; Costa, B.N.S.; de Jesus, I.M.; de Magalhães Câmara, V.; de Oliveira Lima, M. A comparative study of cadmium levels in blood from exposed populations in an industrial area of the Amazon, Brazil. Sci. Total Environ. 2020, 698, 134309. [Google Scholar] [CrossRef]
- Hussain, C.M.; Keçili, R. Environmental pollution and environmental analysis. In Modern Environmental Analysis Techniques for Pollutants; Elsevier: Amsterdam, The Netherlands, 2020; pp. 1–36. ISBN 978-0-12-816934-6. [Google Scholar]
- Amiri, A.; Mirhoseiny, Z. Cadmium contamination of drinking water and its treatment using biological chelators. J. Occup. Health Epidemiol. 2015, 4, 223–228. [Google Scholar] [CrossRef] [Green Version]
- Sherlock, J.C. Cadmium in Foods and the Diet. Experientia 1984, 40, 152–156. [Google Scholar] [CrossRef]
- Turner, A. Cadmium pigments in consumer products and their health risks. Sci. Total Environ. 2019, 657, 1409–1418. [Google Scholar] [CrossRef]
- Hogervorst, J.; Plusquin, M.; Vangronsveld, J.; Nawrot, T.; Cuypers, A.; Van Hecke, E.; Roels, H.A.; Carleer, R.; Staessen, J.A. House dust as possible route of environmental exposure to cadmium and lead in the adult general population. Environ. Res. 2007, 103, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Ganguly, K.; Levänen, B.; Palmberg, L.; Åkesson, A.; Lindén, A. Cadmium in tobacco smokers: A neglected link to lung disease? Eur. Respir. Rev. 2018, 27, 170122. [Google Scholar] [CrossRef]
- Sethi, P.K.; Khandelwal, D.; Sethi, N. Cadmium exposure: Health hazards of silver cottage industry in developing countries. J. Med. Toxicol. 2006, 2, 14–15. [Google Scholar] [CrossRef] [Green Version]
- Genchi, G.; Sinicropi, M.S.; Lauria, G.; Carocci, A.; Catalano, A. The effects of cadmium toxicity. Int. J. Environ. Res. Public Health 2020, 17, 3782. [Google Scholar] [CrossRef] [PubMed]
- Siu, E.R.; Mruk, D.D.; Porto, C.S.; Cheng, C.Y. Cadmium-induced testicular injury. Toxicol. Appl. Pharmacol. 2009, 238, 240–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostrzewska, A.; Sobieszek, A. Diverse actions of cadmium on the smooth muscle myosin phosphorylation system. FEBS Lett. 1990, 263, 381–384. [Google Scholar] [CrossRef] [Green Version]
- Peters, J.L.; Perlstein, T.S.; Perry, M.J.; McNeely, E.; Weuve, J. Cadmium exposure in association with history of stroke and heart failure. Environ. Res. 2010, 110, 199–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branca, J.J.V.; Morucci, G.; Pacini, A. Cadmium-induced neurotoxicity: Still much ado. Neural Regen. Res. 2018, 13, 1879–1882. [Google Scholar]
- Branca, J.J.V.; Maresca, M.; Morucci, G.; Mello, T.; Becatti, M.; Pazzagli, L.; Colzi, I.; Gonnelli, C.; Carrino, D.; Paternostro, F.; et al. Effects of cadmium on ZO-1 tight junction integrity of the blood brain barrier. Int. J. Mol. Sci. 2019, 20, 6010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branca, J.J.V.; Fiorillo, C.; Carrino, D.; Paternostro, F.; Taddei, N.; Gulisano, M.; Pacini, A.; Becatti, M. Cadmium-Induced oxidative stress: Focus on the central nervous system. Antioxidants 2020, 9, 492. [Google Scholar] [CrossRef]
- Prasad, A.S.; Bao, B. Molecular Mechanisms of Zinc as a pro-antioxidant mediator: Clinical therapeutic implications. Antioxidants 2019, 8, 164. [Google Scholar] [CrossRef] [Green Version]
- Maret, W. The redox biology of redox-inert zinc ions. Free Radic. Biol. Med. 2019, 134, 311–326. [Google Scholar] [CrossRef] [Green Version]
- Sabolić, I.; Breljak, D.; Škarica, M.; Herak-Kramberger, C.M. Role of metallothionein in cadmium traffic and toxicity in kidneys and other mammalian organs. Biometals 2010, 23, 897–926. [Google Scholar] [CrossRef]
- Hall, J.L. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot. 2002, 53, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Babula, P.; Masarik, M.; Adam, V.; Eckschlager, T.; Stiborova, M.; Trnkova, L.; Skutkova, H.; Provaznik, I.; Hubalek, J.; Kizek, R. Mammalian metallothioneins: Properties and functions. Metallomics 2012, 4, 739. [Google Scholar] [CrossRef] [PubMed]
- Pinter, T.B.J.; Irvine, G.W.; Stillman, M.J. Domain selection in Metallothionein 1A: Affinity-controlled mechanisms of Zinc binding and cadmium exchange. Biochemistry 2015, 54, 5006–5016. [Google Scholar] [CrossRef] [PubMed]
- Tandon, S.K.; Singh, S.; Prasad, S.; Khandekar, K.; Dwivedi, V.K.; Chatterjee, M.; Mathur, N. Reversal of cadmium induced oxidative stress by chelating agent, antioxidant or their combination in rat. Toxicol. Lett. 2003, 145, 211–217. [Google Scholar] [CrossRef]
- Sandbichler, A.; Höckner, M. Cadmium protection strategies—A hidden trade-off? Int. J. Mol. Sci. 2016, 17, 139. [Google Scholar] [CrossRef] [Green Version]
- Brzóska, M.M.; Borowska, S.; Tomczyk, M. Antioxidants as a potential preventive and therapeutic strategy for cadmium. CDT 2016, 17, 1350–1384. [Google Scholar] [CrossRef]
- Thévenod, F.; Fels, J.; Lee, W.-K.; Zarbock, R. Channels, transporters and receptors for cadmium and cadmium complexes in eukaryotic cells: Myths and facts. Biometals 2019, 32, 469–489. [Google Scholar] [CrossRef]
- Powell, S.R. The antioxidant properties of zinc. J. Nutr. 2000, 130, 1447S–1454S. [Google Scholar] [CrossRef] [Green Version]
- Carrino, D.; Branca, J.J.V.; Becatti, M.; Paternostro, F.; Morucci, G.; Gulisano, M.; Di Cesare Mannelli, L.; Pacini, A. Alcohol-induced blood-brain barrier impairment: An in vitro study. Int. J. Environ. Res. Public Health 2021, 18, 2683. [Google Scholar] [CrossRef]
- Veszelka, S.; Tóth, A.; Walter, F.R.; Tóth, A.E.; Gróf, I.; Mészáros, M.; Bocsik, A.; Hellinger, É.; Vastag, M.; Rákhely, G.; et al. Comparison of a rat primary cell-based blood-brain barrier model with epithelial and brain endothelial cell lines: Gene expression and drug transport. Front. Mol. Neurosci. 2018, 11, 166. [Google Scholar] [CrossRef]
- Branca, J.J.V.; Morucci, G.; Maresca, M.; Tenci, B.; Cascella, R.; Paternostro, F.; Ghelardini, C.; Gulisano, M.; Di Cesare Mannelli, L.; Pacini, A. Selenium and zinc: Two key players against cadmium-induced neuronal toxicity. Toxicol. In Vitro 2018, 48, 159–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branca, J.J.V.; Morucci, G.; Becatti, M.; Carrino, D.; Ghelardini, C.; Gulisano, M.; Di Cesare Mannelli, L.; Pacini, A. Cannabidiol protects dopaminergic neuronal cells from cadmium. Int. J. Environ. Res. Public Health 2019, 16, 4420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- del Pino, J.; Zeballos, G.; Anadón, M.J.; Moyano, P.; Díaz, M.J.; García, J.M.; Frejo, M.T. Cadmium-induced cell death of basal forebrain cholinergic neurons mediated by muscarinic M1 receptor blockade, increase in GSK-3β enzyme, β-Amyloid and Tau protein levels. Arch. Toxicol. 2015, 90, 1081–1092. [Google Scholar] [CrossRef] [PubMed]
- Messner, B.; Türkcan, A.; Ploner, C.; Laufer, G.; Bernhard, D. Cadmium overkill: Autophagy, apoptosis and necrosis signalling in endothelial cells exposed to cadmium. Cell. Mol. Life Sci. 2016, 73, 1699–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.-H.; Shih, C.-M.; Huang, C.-J.; Lin, C.-M.; Chou, C.-M.; Tsai, M.-L.; Liu, T.P.; Chiu, J.-F.; Chen, C.-T. Effects of cadmium on structure and enzymatic activity of Cu, Zn-SOD and oxidative status in neural cells. J. Cell. Biochem. 2006, 98, 577–589. [Google Scholar] [CrossRef]
- Dąbrowski, A.; Onopiuk, B.M.; Car, H.; Onopiuk, P.; Dąbrowska, Z.N.; Rogalska, J.; Brzóska, M.M.; Dąbrowska, E. Beneficial impact of an extract from the berries of Aronia melanocarpa L. on the oxidative-reductive status of the submandibular gland of rats exposed to cadmium. Antioxidants 2020, 9, 185. [Google Scholar] [CrossRef] [Green Version]
- Polykretis, P.; Delfino, G.; Petrocelli, I.; Cervo, R.; Tanteri, G.; Montori, G.; Perito, B.; Branca, J.J.V.; Morucci, G.; Gulisano, M. Evidence of immunocompetence reduction induced by cadmium exposure in Honey Bees (Apis mellifera). Environ. Pollut. 2016, 218, 826–834. [Google Scholar] [CrossRef]
- Junsi, M.; Takahashi Yupanqui, C.; Usawakesmanee, W.; Slusarenko, A.; Siripongvutikorn, S. Thunbergia laurifolia leaf extract increased levels of antioxidant enzymes and protected human cell-lines in vitro against cadmium. Antioxidants 2020, 9, 47. [Google Scholar] [CrossRef] [Green Version]
- Ledda, C.; Cannizzaro, E.; Lovreglio, P.; Vitale, E.; Stufano, A.; Montana, A.; Li Volti, G.; Rapisarda, V. Exposure to toxic heavy metals can influence homocysteine metabolism? Antioxidants 2019, 9, 30. [Google Scholar] [CrossRef] [Green Version]
- Tobwala, S.; Wang, H.-J.; Carey, J.; Banks, W.; Ercal, N. Effects of lead and cadmium on brain endothelial cell survival, monolayer permeability, and crucial oxidative stress markers in an in vitro model of the blood-brain barrier. Toxics 2014, 2, 258–275. [Google Scholar] [CrossRef] [Green Version]
- Negri, S.; Faris, P.; Rosti, V.; Antognazza, M.R.; Lodola, F.; Moccia, F. Endothelial TRPV1 as an emerging molecular target to promote therapeutic angiogenesis. Cells 2020, 29, 1341. [Google Scholar] [CrossRef] [PubMed]
- Negri, S.; Faris, P.; Berra-Romani, R.; Guerra, G.; Moccia, F. Endothelial transient receptor potential channels and vascular remodeling: Extracellular Ca2+ entry for angiogenesis, arteriogenesis and vasculogenesis. Front. Physiol. 2020, 10, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuran Ercal, B.S.P.; Hande Gurer-Orhan, B.S.P.; Nukhet Aykin-Burns, B.S.P. Toxic metals and oxidative stress part I: Mechanisms involved in Me-Tal induced oxidative damage. Curr. Top. Med. Chem. 2001, 1, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.-H.; Chen, C.-M. The role of oxidative stress in Parkinson’s disease. Antioxidants 2020, 9, 597. [Google Scholar] [CrossRef]
- Patra, R.C.; Rautray, A.K.; Swarup, D. Oxidative Stress in Lead and Cadmium Toxicity and Its Amelioration. Vet. Med. Int. 2011, 2011, 457327. [Google Scholar] [CrossRef] [Green Version]
- Zoidis, E.; Papadomichelakis, G.; Pappas, A.C.; Theodorou, G.; Fegeros, K. Effects of selenium and cadmium on breast muscle fatty-acid composition and gene expression of liver antioxidant proteins in broilers. Antioxidants 2019, 8, 147. [Google Scholar] [CrossRef] [Green Version]
- Dubey, S.K.; Ram, M.S.; Krishna, K.V.; Saha, R.N.; Singhvi, G.; Agrawal, M.; Ajazuddin; Saraf, S.; Saraf, S.; Alexander, A. Recent expansions on cellular models to uncover the scientific barriers towards drug development for Alzheimer’s disease. Cell. Mol. Neurobiol. 2019, 39, 181–209. [Google Scholar] [CrossRef]
- Wilhelm, I.; Fazakas, C.; Krizbai, I.A. In vitro models of the blood-brain barrier. Acta Neurobiol. Exp. 2011, 71, 113–128. [Google Scholar]
- Roux, F.; Durieu-Trautmann, O.; Chaverot, N.; Claire, M.; Mailly, P.; Bourre, J.-M.; Strosberg, A.D.; Couraud, P.-O. Regulation of gamma-glutamyl transpeptidase and alkaline phosphatase activities in immortalized rat brain microvessel endothelial cells. J. Cell. Physiol. 1994, 159, 101–113. [Google Scholar] [CrossRef]
- Regina, A.; Koman, A.; Piciotti, M.; El Hafny, B.; Center, M.S.; Bergmann, R.; Couraud, P.-O.; Roux, F. Mrp1 Multidrug resistance-associated protein and P-Glycoprotein expression in rat brain microvessel endothelial cells. J. Neurochem. 2002, 71, 705–715. [Google Scholar] [CrossRef]
- Krizbai, I.; Szabó, G.; Deli, M.; Maderspach, K.; Lehel, C.; Oláh, Z.; Wolff, J.R.; Joó, F. Expression of protein Kinase C family members in the cerebral endothelial cells. J. Neurochem. 2002, 65, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Fábián, G.; Szabó, C.; Bozó, B.; Greenwood, J.; Adamson, P.; Deli, M.A.; Joó, F.; Krizbai, I.A.; Szû, M. Expression of G-Protein subtypes in cultured cerebral endothelial cells. Neurochem. Int. 1998, 33, 179–185. [Google Scholar] [CrossRef]
- Smith, J.P.; Drewes, L.R. Modulation of monocarboxylic acid Transporter-1 kinetic function by the CAMP signaling pathway in rat brain endothelial cells. J. Biol. Chem. 2006, 281, 2053–2060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilorget, A.; Demeule, M.; Barakat, S.; Marvaldi, J.; Luis, J.; Béliveau, R. Modulation of P-Glycoprotein function by Sphingosine Kinase-1 in brain endothelial cells: Sphingosine Kinase-1 Stimulates P-Gp transport activity. J. Neurochem. 2007, 100, 1203–1210. [Google Scholar] [CrossRef]
- Barakat, S.; Turcotte, S.; Demeule, M.; Lachambre, M.-P.; Régina, A.; Baggetto, L.G.; Béliveau, R. Regulation of brain endothelial cells migration and angiogenesis by P-Glycoprotein/Caveolin-1 interaction. Biochem. Biophys. Res. Commun. 2008, 372, 440–446. [Google Scholar] [CrossRef]
- Pan, W.; Yu, Y.; Cain, C.M.; Nyberg, F.; Couraud, P.O.; Kastin, A.J. Permeation of growth hormone across the blood-brain barrier. Endocrinology 2005, 146, 4898–4904. [Google Scholar] [CrossRef]
- Costas, J. The highly pleiotropic gene SLC39A8 as an opportunity to gain insight into the molecular pathogenesis of schizophrenia. Am. J. Med. Genet. 2018, 177, 274–283. [Google Scholar] [CrossRef] [Green Version]
- Steimle, B.L.; Smith, F.M.; Kosman, D.J. The solute carriers ZIP8 and ZIP14 regulate manganese accumulation in brain microvascular endothelial cells and control brain manganese levels. J. Biol. Chem. 2019, 294, 19197–19208. [Google Scholar] [CrossRef]
- Marreiro, D.; Cruz, K.; Morais, J.; Beserra, J.; Severo, J.; de Oliveira, A. Zinc and oxidative stress: Current mechanisms. Antioxidants 2017, 6, 24. [Google Scholar] [CrossRef]
- Brzóska, M.M.; Kozłowska, M.; Rogalska, J.; Gałażyn-Sidorczuk, M.; Roszczenko, A.; Smereczański, N.M. Enhanced zinc intake protects against oxidative stress and its consequences in the brain: A study in an in vivo rat model of cadmium exposure. Nutrients 2021, 13, 478. [Google Scholar] [CrossRef]
- Souza, V.; del C. Escobar, M.; Bucio, L.; Hernández, E.; Gutiérrez-Ruiz, M.C. Zinc pretreatment prevents hepatic stellate cells from cadmium-produced oxidative damage. Cell Biol. Toxicol. 2004, 20, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Cha, M.; Lee, B.H. Neuroprotective effect of antioxidants in the brain. Int. J. Mol. Sci. 2020, 21, 7152. [Google Scholar] [CrossRef] [PubMed]
- Chatzidimitriou, E.; Bisaccia, P.; Corrà, F.; Bonato, M.; Irato, P.; Manuto, L.; Toppo, S.; Bakiu, R.; Santovito, G. Copper/Zinc superoxide dismutase from the crocodile icefish chionodraco hamatus: Antioxidant defense at constant Sub-Zero temperature. Antioxidants 2020, 9, 325. [Google Scholar] [CrossRef] [PubMed]
- Heller, R.A.; Sperl, A.; Seelig, J.; Haubruck, P.; Bock, T.; Werner, T.; Besseling, A.; Sun, Q.; Schomburg, L.; Moghaddam, A.; et al. Zinc concentration dynamics indicate neurological impairment odds after traumatic spinal cord injury. Antioxidants 2020, 9, 421. [Google Scholar] [CrossRef] [PubMed]
- Qi, Z.; Liu, K.J. The interaction of zinc and the blood-brain barrier under physiological and ischemic conditions. Toxicol. Appl. Pharmacol. 2019, 364, 114–119. [Google Scholar] [CrossRef]
- Lehmann, H.M.; Brothwell, B.B.; Volak, L.P.; Bobilya, D.J. Zinc status influences zinc transport by porcine brain capillary endothelial cells. J. Nutr. 2002, 132, 2763–2768. [Google Scholar] [CrossRef] [Green Version]
- Bobilya, D.J.; Gauthier, N.A.; Karki, S.; Olley, B.J.; Thomas, W.K. Longitudinal changes in zinc transport kinetics, metallothionein and zinc transporter expression in a blood–brain barrier model in response to a moderately excessive zinc environment. J. Nutr. Biochem. 2008, 19, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Murphy, M.P. Mitochondrial dysfunction indirectly elevates ROS production by the endoplasmic reticulum. Cell Metab. 2013, 18, 145–146. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, A.P.; Klocke, B.J.; Ballestas, M.E.; Roth, K.A. CHOP Potentially Co-Operates with FOXO3a in neuronal cells to regulate PUMA and BIM expression in response to ER stress. PLoS ONE 2012, 7, e39586. [Google Scholar] [CrossRef]
- Zheng, J.-Y. Overexpression of bax induces apoptosis and enhances drug sensitivity of hepatocellular cancer-9204 cells. World J. Gastroenterol. 2005, 11, 3498. [Google Scholar] [CrossRef]
- Song, Y.; Xue, Y.; Liu, X.; Wang, P.; Liu, L. Effects of acute exposure to aluminum on blood–brain barrier and the protection of zinc. Neurosci. Lett. 2008, 445, 42–46. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cotreatment | Complete Growth Medium | ZnCl2 | CdCl2 |
| 0 h | + | - | - |
| 24 h (8 h for ROS evaluation) | - | + | + |
| Pretreatment | Complete Growth Medium | ZnCl2 | CdCl2 |
| 0 h | + | - | - |
| 24 h (8 h for ROS evaluation) | - | + | - |
| 48 h (16 h for ROS evaluation) | - | - | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Branca, J.J.V.; Carrino, D.; Paternostro, F.; Morucci, G.; Fiorillo, C.; Nicoletti, C.; Gulisano, M.; Ghelardini, C.; Di Cesare Mannelli, L.; Becatti, M.; et al. The Protection of Zinc against Acute Cadmium Exposure: A Morphological and Molecular Study on a BBB In Vitro Model. Cells 2022, 11, 1646. https://doi.org/10.3390/cells11101646
Branca JJV, Carrino D, Paternostro F, Morucci G, Fiorillo C, Nicoletti C, Gulisano M, Ghelardini C, Di Cesare Mannelli L, Becatti M, et al. The Protection of Zinc against Acute Cadmium Exposure: A Morphological and Molecular Study on a BBB In Vitro Model. Cells. 2022; 11(10):1646. https://doi.org/10.3390/cells11101646
Chicago/Turabian StyleBranca, Jacopo J. V., Donatello Carrino, Ferdinando Paternostro, Gabriele Morucci, Claudia Fiorillo, Claudio Nicoletti, Massimo Gulisano, Carla Ghelardini, Lorenzo Di Cesare Mannelli, Matteo Becatti, and et al. 2022. "The Protection of Zinc against Acute Cadmium Exposure: A Morphological and Molecular Study on a BBB In Vitro Model" Cells 11, no. 10: 1646. https://doi.org/10.3390/cells11101646