
Challenges in Cell Fate Acquisition to Scid-Repopulating Activity from Hemogenic Endothelium of hiPSCs Derived from AML Patients Using Forced Transcription Factor Expression

Abstract
:1. Introduction
2. Material & Methods
2.1. Human iPSC Lines
2.2. hPSC Culture
2.3. hPSC EB Differentiation
2.4. hPSC EHT Differentiation
2.5. Lentiviral Gene Transfer
2.6. Xenotransplantation
2.7. Colony Forming Unit (CFU) Assay
2.8. Western Blotting
2.9. RT-qPCR
2.10. Conventional PCR
2.11. Flow Cytometry
2.12. Gene Expression Profiling
2.13. Statistical Analysis
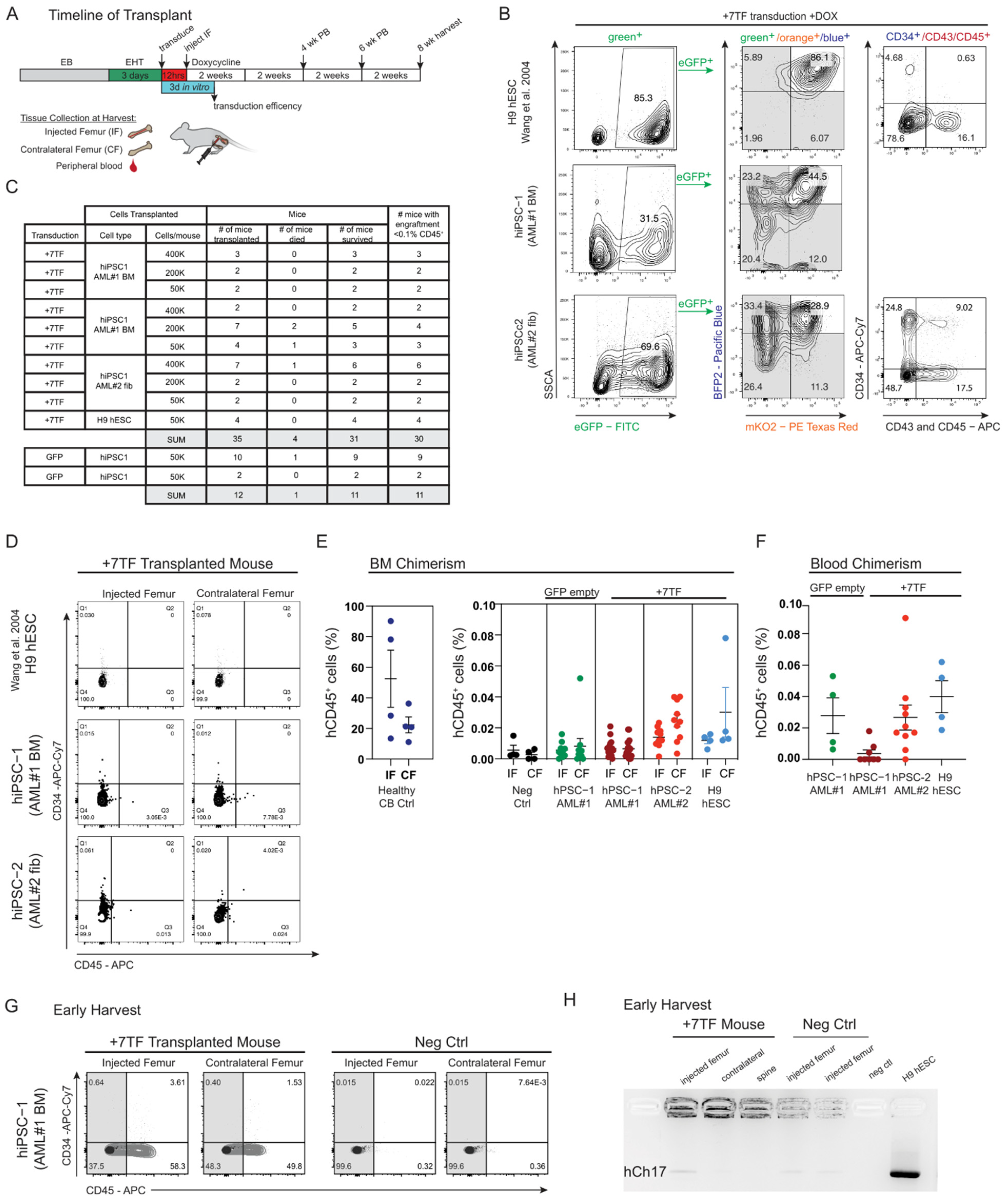
3. Results
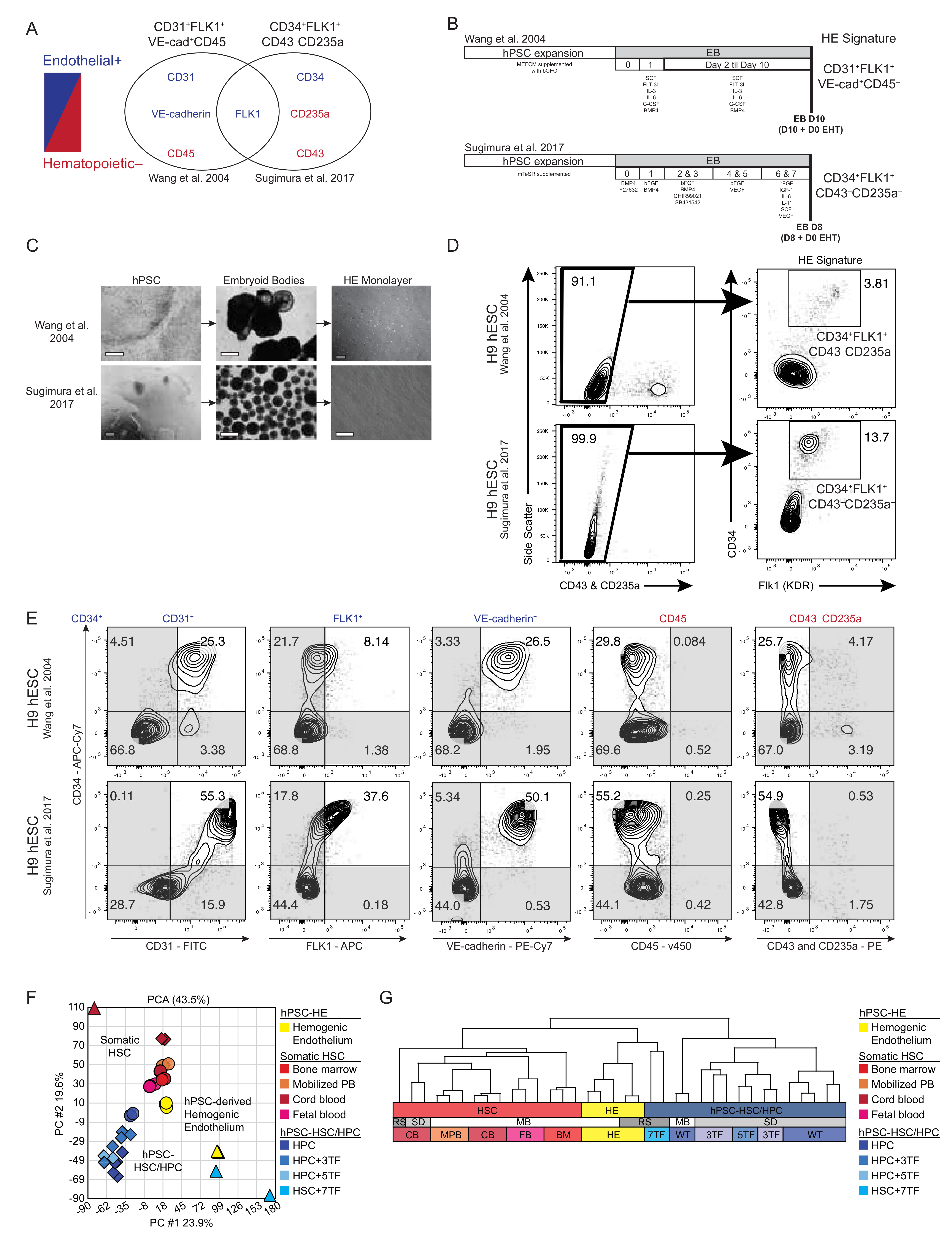
3.1. Molecular Comparison of Human HE Derivation from Pluripotent Cell
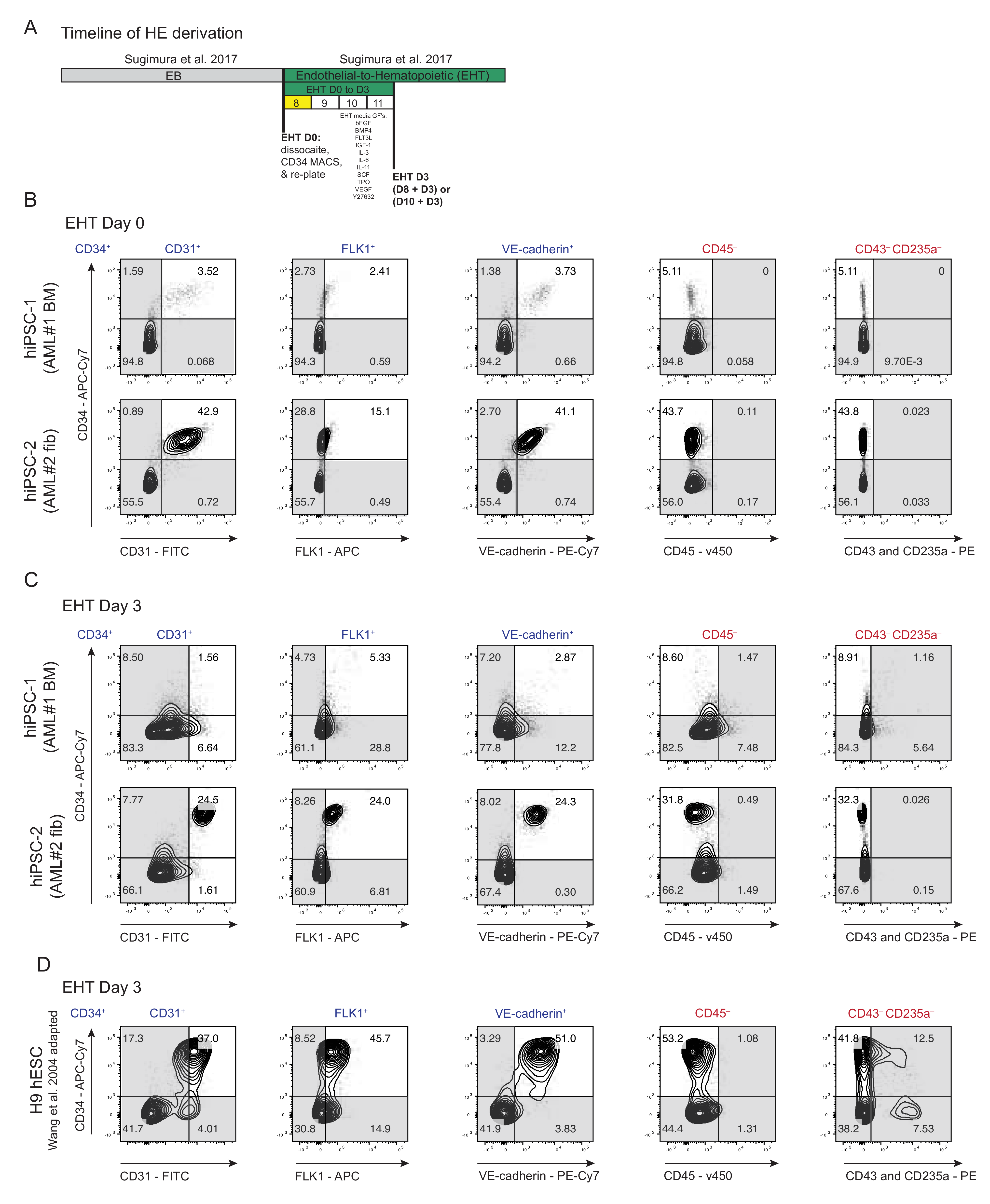
3.2. Derivation of HE from AML Patient-Specific iPSCs into Endothelial-to-Hematopoietic Transition (EHT) Conditions for Forced 7TF Expression
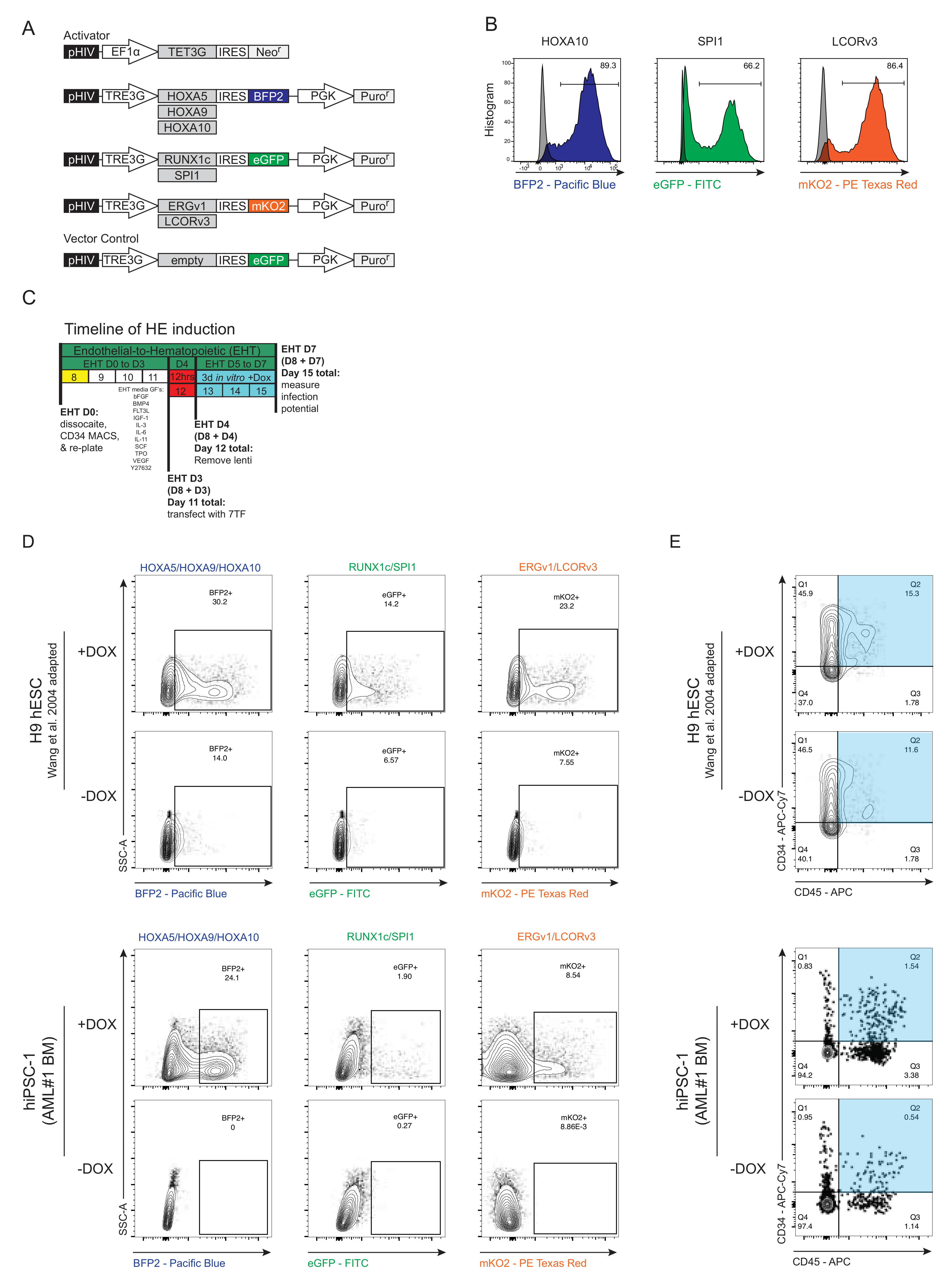
3.3. Generation and Forced Expression of 7TF under HE In Vitro Differentiation Conditions from hPSCs
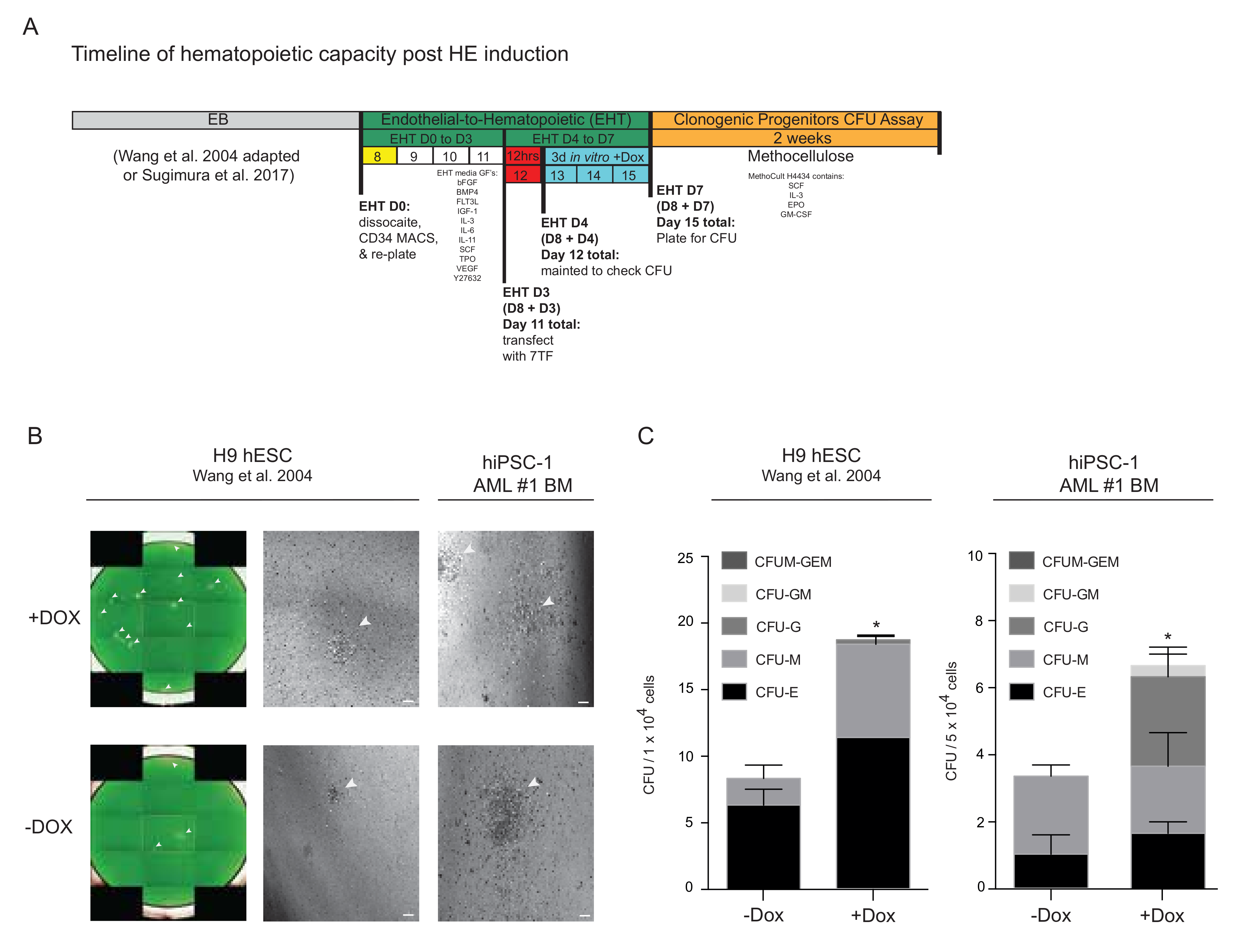
3.4. Progenitor Capacity from hPSC-Derived HE with 7TF Expression
3.5. SRC Engraftment Potential of TF-Induced Hemogenic Precursors Derived from Healthy iPSCs Established from AML Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Doulatov, S.; Notta, F.; Eppert, K.; Nguyen, L.T.; Ohashi, P.S.; Dick, J.E. Revised Map of the Human Progenitor Hierarchy Shows the Origin of Macrophages and Dendritic Cells in Early Lymphoid Development. Nat. Immunol. 2010, 11, 585–593. [Google Scholar] [CrossRef]
- Notta, F.; Doulatov, S.; Laurenti, E.; Poeppl, A.; Jurisica, I.; Dick, J.E. Isolation of Single Human Hematopoietic Stem Cells Capable of Long-Term Multilineage Engraftment. Science 2011, 333, 218–221. [Google Scholar] [CrossRef]
- Hope, K.; Bhatia, M. Clonal Interrogation of Stem Cells. Nat. Methods 2011, 8, S36–S40. [Google Scholar] [CrossRef]
- Kondo, M.; Wagers, A.J.; Manz, M.G.; Prohaska, S.S.; Scherer, D.C.; Beilhack, G.F.; Shizuru, J.A.; Weissman, I.L. Biology of Hematopoietic Stem Cells and Progenitors: Implications for Clinical Application. Annu. Rev. Immunol. 2003, 21, 759–806. [Google Scholar] [CrossRef]
- Kørbling, M.; Estrov, Z. Adult Stem Cells for Tissue Repair—A New Therapeutic Concept? N. Engl. J. Med. 2003, 349, 570–582. [Google Scholar] [CrossRef] [Green Version]
- Sureda, A.; Bader, P.; Cesaro, S.; Dreger, P.; Duarte, R.F.; Dufour, C.; Falkenburg, J.H.F.; Farge-Bancel, D.; Gennery, A.; Kröger, N.; et al. Indications for Allo- and Auto-SCT for Haematological Diseases, Solid Tumours and Immune Disorders: Current Practice in Europe, 2015. Bone Marrow Transplant. 2015, 50, 1037–1056. [Google Scholar] [CrossRef] [Green Version]
- Löwenberg, B.; Downing, J.R.; Burnett, A. Acute Myeloid Leukemia. N. Engl. J. Med. 1999, 341, 1051–1062. [Google Scholar] [CrossRef]
- Shipley, J.L.; Butera, J.N. Acute Myelogenous Leukemia. Exp. Hematol. 2009, 37, 649–658. [Google Scholar] [CrossRef]
- Boyd, A.L.; Campbell, C.J.V.; Hopkins, C.I.; Fiebig-Comyn, A.; Russell, J.; Ulemek, J.; Foley, R.; Leber, B.; Xenocostas, A.; Collins, T.J.; et al. Niche Displacement of Human Leukemic Stem Cells Uniquely Allows Their Competitive Replacement with Healthy HSPCs. J. Exp. Med. 2014, 211, 1925–1935. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, A.M.; Norden, J.; Li, S.; Hromadnikova, I.; Schmid, C.; Schmetzer, H.; Jochem-Kolb, H. Graft-versus-Leukemia Effect Following Hematopoietic Stem Cell Transplantation for Leukemia. Front. Immunol. 2017, 8, 496. [Google Scholar] [CrossRef]
- Salci, K.R.; McIntyre, B.A.; Bhatia, M. Foundational Concepts of Cell Fate Conversion to the Hematopoietic Lineage. Curr. Opin. Genet. Dev. 2013, 23, 585–590. [Google Scholar] [CrossRef]
- Dzierzak, E.; Bigas, A. Blood Development: Hematopoietic Stem Cell Dependence and Independence. Cell Stem Cell 2018, 22, 639–651. [Google Scholar] [CrossRef] [Green Version]
- Zambidis, E.T.; Sinka, L.; Tavian, M.; Jokubaitis, V.; Park, T.S.; Simmons, P.; Péault, B. Emergence of Human Angiohematopoietic Cells in Normal Development and from Cultured Embryonic Stem Cells. Ann. N. Y. Acad. Sci. 2007, 1106, 223–232. [Google Scholar] [CrossRef]
- Medvinsky, A.; Dzierzak, E. Definitive Hematopoiesis Is Autonomously Initiated by the AGM Region. Cell 1996, 86, 897–906. [Google Scholar] [CrossRef] [Green Version]
- Bertrand, J.Y.; Chi, N.C.; Santoso, B.; Teng, S.; Stainier, D.Y.R.; Traver, D. Haematopoietic Stem Cells Derive Directly from Aortic Endothelium during Development. Nature 2010, 464, 108–111. [Google Scholar] [CrossRef] [Green Version]
- De Bruijn, M.F.T.R.; Ma, X.; Robin, C.; Ottersbach, K.; Sanchez, M.-J.; Dzierzak, E. Hematopoietic Stem Cells Localize to the Endothelial Cell Layer in the Midgestation Mouse Aorta. Immunity 2002, 16, 673–683. [Google Scholar] [CrossRef] [Green Version]
- Eilken, H.M.; Nishikawa, S.-I.; Schroeder, T. Continuous Single-Cell Imaging of Blood Generation from Haemogenic Endothelium. Nature 2009, 457, 896–900. [Google Scholar] [CrossRef]
- Zovein, A.C.; Hofmann, J.J.; Lynch, M.; French, W.J.; Turlo, K.A.; Yang, Y.; Becker, M.S.; Zanetta, L.; Dejana, E.; Gasson, J.C.; et al. Fate Tracing Reveals the Endothelial Origin of Hematopoietic Stem Cells. Cell Stem Cell 2008, 3, 625–636. [Google Scholar] [CrossRef] [Green Version]
- Dzierzak, E.; Speck, N.A. Of Lineage and Legacy: The Development of Mammalian Hematopoietic Stem Cells. Nat. Immunol. 2008, 9, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, D.S.; Hanson, E.T.; Lewis, R.L.; Auerbach, R.; Thomson, J.A. Hematopoietic Colony-Forming Cells Derived from Human Embryonic Stem Cells. Proc. Natl. Acad. Sci. USA 2001, 98, 10716–10721. [Google Scholar] [CrossRef] [Green Version]
- Vodyanik, M.A.; Bork, J.A.; Thomson, J.A.; Slukvin, I.I. Human Embryonic Stem Cell-Derived CD34+ Cells: Efficient Production in the Coculture with OP9 Stromal Cells and Analysis of Lymphohematopoietic Potential. Blood 2005, 105, 617–626. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Woll, P.S.; Morris, J.K.; Linehan, J.L.; Kaufman, D.S. Hematopoietic Engraftment of Human Embryonic Stem Cell-Derived Cells Is Regulated by Recipient Innate Immunity. Stem Cells Dayt. Ohio 2006, 24, 1370–1380. [Google Scholar] [CrossRef]
- Wang, L.; Menendez, P.; Shojaei, F.; Li, L.; Mazurier, F.; Dick, J.E.; Cerdan, C.; Levac, K.; Bhatia, M. Generation of Hematopoietic Repopulating Cells from Human Embryonic Stem Cells Independent of Ectopic HOXB4 Expression. J. Exp. Med. 2005, 201, 1603–1614. [Google Scholar] [CrossRef]
- Reid, J.C.; Tanasijevic, B.; Golubeva, D.; Boyd, A.L.; Porras, D.P.; Collins, T.J.; Bhatia, M. CXCL12/CXCR4 Signaling Enhances Human PSC-Derived Hematopoietic Progenitor Function and Overcomes Early In Vivo Transplantation Failure. Stem Cell Rep. 2018, 10, 1625–1641. [Google Scholar] [CrossRef]
- Perrimon, N.; Pitsouli, C.; Shilo, B.-Z. Signaling Mechanisms Controlling Cell Fate and Embryonic Patterning. Cold Spring Harb. Perspect. Biol. 2012, 4, a005975. [Google Scholar] [CrossRef] [Green Version]
- Dick, J.E. Normal and Leukemic Human Stem Cells Assayed in SCID Mice. Semin. Immunol. 1996, 8, 197–206. [Google Scholar] [CrossRef]
- Doulatov, S.; Notta, F.; Laurenti, E.; Dick, J.E. Hematopoiesis: A Human Perspective. Cell Stem Cell 2012, 10, 120–136. [Google Scholar] [CrossRef] [Green Version]
- Salci, K.R.; Lee, J.-H.; Laronde, S.; Dingwall, S.; Kushwah, R.; Fiebig-Comyn, A.; Leber, B.; Foley, R.; Dal Cin, A.; Bhatia, M. Cellular Reprogramming Allows Generation of Autologous Hematopoietic Progenitors From AML Patients That Are Devoid of Patient-Specific Genomic Aberrations. Stem Cells Dayt. Ohio 2015, 33, 1839–1849. [Google Scholar] [CrossRef]
- Doulatov, S.; Vo, L.T.; Chou, S.S.; Kim, P.G.; Arora, N.; Li, H.; Hadland, B.K.; Bernstein, I.D.; Collins, J.J.; Zon, L.I.; et al. Induction of Multipotential Hematopoietic Progenitors from Human Pluripotent Stem Cells via Respecification of Lineage-Restricted Precursors. Cell Stem Cell 2013, 13, 459–470. [Google Scholar] [CrossRef] [Green Version]
- Lis, R.; Karrasch, C.C.; Poulos, M.G.; Kunar, B.; Redmond, D.; Duran, J.G.B.; Badwe, C.R.; Schachterle, W.; Ginsberg, M.; Xiang, J.; et al. Conversion of Adult Endothelium to Immunocompetent Haematopoietic Stem Cells. Nature 2017, 545, 439–445. [Google Scholar] [CrossRef]
- Sugimura, R.; Jha, D.K.; Han, A.; Soria-Valles, C.; da Rocha, E.L.; Lu, Y.-F.; Goettel, J.A.; Serrao, E.; Rowe, R.G.; Malleshaiah, M.; et al. Haematopoietic Stem and Progenitor Cells from Human Pluripotent Stem Cells. Nature 2017, 545, 432–438. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-H.; Salci, K.R.; Reid, J.C.; Orlando, L.; Tanasijevic, B.; Shapovalova, Z.; Bhatia, M. Brief Report: Human Acute Myeloid Leukemia Reprogramming to Pluripotency Is a Rare Event and Selects for Patient Hematopoietic Cells Devoid of Leukemic Mutations. Stem Cells Dayt. Ohio 2017, 35, 2095–2102. [Google Scholar] [CrossRef] [Green Version]
- Chadwick, K.; Wang, L.; Li, L.; Menendez, P.; Murdoch, B.; Rouleau, A.; Bhatia, M. Cytokines and BMP-4 Promote Hematopoietic Differentiation of Human Embryonic Stem Cells. Blood 2003, 102, 906–915. [Google Scholar] [CrossRef]
- Orlando, L.; Tanasijevic, B.; Nakanishi, M.; Reid, J.C.; García-Rodríguez, J.L.; Chauhan, K.D.; Porras, D.P.; Aslostovar, L.; Lu, J.D.; Shapovalova, Z.; et al. Phosphorylation State of the Histone Variant H2A.X Controls Human Stem and Progenitor Cell Fate Decisions. Cell Rep. 2021, 34, 108818. [Google Scholar] [CrossRef]
- Nakamura, T.; Okamoto, I.; Sasaki, K.; Yabuta, Y.; Iwatani, C.; Tsuchiya, H.; Seita, Y.; Nakamura, S.; Yamamoto, T.; Saitou, M. A Developmental Coordinate of Pluripotency among Mice, Monkeys and Humans. Nature 2016, 537, 57–62. [Google Scholar] [CrossRef]
- Lange, L.; Morgan, M.; Schambach, A. The Hemogenic Endothelium: A Critical Source for the Generation of PSC-Derived Hematopoietic Stem and Progenitor Cells. Cell. Mol. Life Sci. 2021, 78, 4143–4160. [Google Scholar] [CrossRef]
- Wang, L.; Li, L.; Shojaei, F.; Levac, K.; Cerdan, C.; Menendez, P.; Martin, T.; Rouleau, A.; Bhatia, M. Endothelial and Hematopoietic Cell Fate of Human Embryonic Stem Cells Originates from Primitive Endothelium with Hemangioblastic Properties. Immunity 2004, 21, 31–41. [Google Scholar] [CrossRef] [Green Version]
- Ditadi, A.; Sturgeon, C.M.; Tober, J.; Awong, G.; Kennedy, M.; Yzaguirre, A.D.; Azzola, L.; Ng, E.S.; Stanley, E.G.; French, D.L.; et al. Human Definitive Haemogenic Endothelium and Arterial Vascular Endothelium Represent Distinct Lineages. Nat. Cell Biol. 2015, 17, 580–591. [Google Scholar] [CrossRef] [Green Version]
- Ditadi, A.; Sturgeon, C.M. Directed Differentiation of Definitive Hemogenic Endothelium and Hematopoietic Progenitors from Human Pluripotent Stem Cells. Methods 2016, 101, 65–72. [Google Scholar] [CrossRef]
- McIntyre, B.A.S.; Ramos-Mejia, V.; Rampalli, S.; Mechael, R.; Lee, J.-H.; Alev, C.; Sheng, G.; Bhatia, M. Gli3-Mediated Hedgehog Inhibition in Human Pluripotent Stem Cells Initiates and Augments Developmental Programming of Adult Hematopoiesis. Blood 2013, 121, 1543–1552. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.B.; Werbowetski-Ogilvie, T.E.; Lee, J.-H.; McIntyre, B.A.S.; Schnerch, A.; Hong, S.-H.; Park, I.-H.; Daley, G.Q.; Bernstein, I.D.; Bhatia, M. Notch-HES1 Signaling Axis Controls Hemato-Endothelial Fate Decisions of Human Embryonic and Induced Pluripotent Stem Cells. Blood 2013, 122, 1162–1173. [Google Scholar] [CrossRef] [Green Version]
- Nishikawa, S.I.; Nishikawa, S.; Hirashima, M.; Matsuyoshi, N.; Kodama, H. Progressive Lineage Analysis by Cell Sorting and Culture Identifies FLK1+VE-Cadherin+ Cells at a Diverging Point of Endothelial and Hemopoietic Lineages. Dev. Camb. Engl. 1998, 125, 1747–1757. [Google Scholar] [CrossRef]
- Testa, U.; Labbaye, C.; Castelli, G.; Pelosi, E. Oxidative Stress and Hypoxia in Normal and Leukemic Stem Cells. Exp. Hematol. 2016, 44, 540–560. [Google Scholar] [CrossRef] [Green Version]
- Suda, T.; Takubo, K.; Semenza, G.L. Metabolic Regulation of Hematopoietic Stem Cells in the Hypoxic Niche. Cell Stem Cell 2011, 9, 298–310. [Google Scholar] [CrossRef] [Green Version]
- Miharada, K.; Karlsson, G.; Rehn, M.; Rörby, E.; Siva, K.; Cammenga, J.; Karlsson, S. Hematopoietic Stem Cells Are Regulated by Cripto, as an Intermediary of HIF-1α in the Hypoxic Bone Marrow Niche. Ann. N. Y. Acad. Sci. 2012, 1266, 55–62. [Google Scholar] [CrossRef]
- Lee, J.; Dykstra, B.; Spencer, J.A.; Kenney, L.L.; Greiner, D.L.; Shultz, L.D.; Brehm, M.A.; Lin, C.P.; Sackstein, R.; Rossi, D.J. MRNA-Mediated Glycoengineering Ameliorates Deficient Homing of Human Stem Cell-Derived Hematopoietic Progenitors. J. Clin. Investig. 2017, 127, 2433–2437. [Google Scholar] [CrossRef] [Green Version]
- Jaatinen, T.; Hemmoranta, H.; Hautaniemi, S.; Niemi, J.; Nicorici, D.; Laine, J.; Yli-Harja, O.; Partanen, J. Global Gene Expression Profile of Human Cord Blood-Derived CD133+ Cells. Stem Cells Dayt. Ohio 2006, 24, 631–641. [Google Scholar] [CrossRef]
- Eppert, K.; Takenaka, K.; Lechman, E.R.; Waldron, L.; Nilsson, B.; van Galen, P.; Metzeler, K.H.; Poeppl, A.; Ling, V.; Beyene, J.; et al. Stem Cell Gene Expression Programs Influence Clinical Outcome in Human Leukemia. Nat. Med. 2011, 17, 1086–1093. [Google Scholar] [CrossRef]
- Cioch, M.; Jawniak, D.; Wach, M.; Mańko, J.; Radomska, K.; Borowska, H.; Szczepanek, A.; Hus, M. Autologous Hematopoietic Stem Cell Transplantation for Adults With Acute Myeloid Leukemia. Transplant. Proc. 2016, 48, 1814–1817. [Google Scholar] [CrossRef]
- Simancikova, I.; Bojtarova, E.; Hrubisko, M.; Farkas, F.; Horvathova, D.; Sopko, L.; Batorova, A.; Mistrik, M. Autologous Hematopoietic Stem Cell Transplantation for Acute Myeloid Leukemia—Single Center Experience. Neoplasma 2017, 64, 738–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zittoun, R.A.; Mandelli, F.; Willemze, R.; de Witte, T.; Labar, B.; Resegotti, L.; Leoni, F.; Damasio, E.; Visani, G.; Papa, G. Autologous or Allogeneic Bone Marrow Transplantation Compared with Intensive Chemotherapy in Acute Myelogenous Leukemia. European Organization for Research and Treatment of Cancer (EORTC) and the Gruppo Italiano Malattie Ematologiche Maligne Dell’Adulto (GIMEMA) Leukemia Cooperative Groups. N. Engl. J. Med. 1995, 332, 217–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kissa, K.; Herbomel, P. Blood Stem Cells Emerge from Aortic Endothelium by a Novel Type of Cell Transition. Nature 2010, 464, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Sturgeon, C.M.; Ditadi, A.; Awong, G.; Kennedy, M.; Keller, G. Wnt Signaling Controls the Specification of Definitive and Primitive Hematopoiesis from Human Pluripotent Stem Cells. Nat. Biotechnol. 2014, 32, 554–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoch, C.; Schnittger, S.; Klaus, M.; Kern, W.; Hiddemann, W.; Haferlach, T. AML with 11q23/MLL Abnormalities as Defined by the WHO Classification: Incidence, Partner Chromosomes, FAB Subtype, Age Distribution, and Prognostic Impact in an Unselected Series of 1897 Cytogenetically Analyzed AML Cases. Blood 2003, 102, 2395–2402. [Google Scholar] [CrossRef] [Green Version]
- Salek-Ardakani, S.; Smooha, G.; de Boer, J.; Sebire, N.J.; Morrow, M.; Rainis, L.; Lee, S.; Williams, O.; Izraeli, S.; Brady, H.J.M. ERG Is a Megakaryocytic Oncogene. Cancer Res. 2009, 69, 4665–4673. [Google Scholar] [CrossRef] [Green Version]
- Tomlins, S.A.; Rhodes, D.R.; Perner, S.; Dhanasekaran, S.M.; Mehra, R.; Sun, X.-W.; Varambally, S.; Cao, X.; Tchinda, J.; Kuefer, R.; et al. Recurrent Fusion of TMPRSS2 and ETS Transcription Factor Genes in Prostate Cancer. Science 2005, 310, 644–648. [Google Scholar] [CrossRef]
- Sorensen, P.H.; Lessnick, S.L.; Lopez-Terrada, D.; Liu, X.F.; Triche, T.J.; Denny, C.T. A Second Ewing’s Sarcoma Translocation, t(21;22), Fuses the EWS Gene to Another ETS-Family Transcription Factor, ERG. Nat. Genet. 1994, 6, 146–151. [Google Scholar] [CrossRef]
- Jalaguier, S.; Teyssier, C.; Nait Achour, T.; Lucas, A.; Bonnet, S.; Rodriguez, C.; Elarouci, N.; Lapierre, M.; Cavaillès, V. Complex Regulation of LCoR Signaling in Breast Cancer Cells. Oncogene 2017, 36, 4790–4801. [Google Scholar] [CrossRef]
- Cawthorne, C.; Swindell, R.; Stratford, I.J.; Dive, C.; Welman, A. Comparison of Doxycycline Delivery Methods for Tet-Inducible Gene Expression in a Subcutaneous Xenograft Model. J. Biomol. Tech. JBT 2007, 18, 120–123. [Google Scholar]
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic Stem Cell Lines Derived from Human Blastocysts. Science 1998, 282, 1145–1147. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Vodyanik, M.A.; Smuga-Otto, K.; Antosiewicz-Bourget, J.; Frane, J.L.; Tian, S.; Nie, J.; Jonsdottir, G.A.; Ruotti, V.; Stewart, R.; et al. Induced Pluripotent Stem Cell Lines Derived from Human Somatic Cells. Science 2007, 318, 1917–1920. [Google Scholar] [CrossRef]
- Choi, K.-D.; Vodyanik, M.A.; Slukvin, I.I. Generation of Mature Human Myelomonocytic Cells through Expansion and Differentiation of Pluripotent Stem Cell-Derived Lin-CD34+CD43+CD45+ Progenitors. J. Clin. Investig. 2009, 119, 2818–2829. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, M.; Awong, G.; Sturgeon, C.M.; Ditadi, A.; LaMotte-Mohs, R.; Zúñiga-Pflücker, J.C.; Keller, G. T Lymphocyte Potential Marks the Emergence of Definitive Hematopoietic Progenitors in Human Pluripotent Stem Cell Differentiation Cultures. Cell Rep. 2012, 2, 1722–1735. [Google Scholar] [CrossRef] [Green Version]
- Risueño, R.M.; Sachlos, E.; Lee, J.-H.; Lee, J.B.; Hong, S.-H.; Szabo, E.; Bhatia, M. Inability of Human Induced Pluripotent Stem Cell-Hematopoietic Derivatives to Downregulate MicroRNAs in Vivo Reveals a Block in Xenograft Hematopoietic Regeneration. Stem Cells Dayt. Ohio 2012, 30, 131–139. [Google Scholar] [CrossRef]
- Narayan, A.D.; Chase, J.L.; Lewis, R.L.; Tian, X.; Kaufman, D.S.; Thomson, J.A.; Zanjani, E.D. Human Embryonic Stem Cell-Derived Hematopoietic Cells Are Capable of Engrafting Primary as Well as Secondary Fetal Sheep Recipients. Blood 2006, 107, 2180–2183. [Google Scholar] [CrossRef] [Green Version]
- Chang, K.-H.; Nelson, A.M.; Cao, H.; Wang, L.; Nakamoto, B.; Ware, C.B.; Papayannopoulou, T. Definitive-like Erythroid Cells Derived from Human Embryonic Stem Cells Coexpress High Levels of Embryonic and Fetal Globins with Little or No Adult Globin. Blood 2006, 108, 1515–1523. [Google Scholar] [CrossRef] [Green Version]
- Chicha, L.; Feki, A.; Boni, A.; Irion, O.; Hovatta, O.; Jaconi, M. Human Pluripotent Stem Cells Differentiated in Fully Defined Medium Generate Hematopoietic CD34- and CD34+ Progenitors with Distinct Characteristics. PLoS ONE 2011, 6, e14733. [Google Scholar] [CrossRef] [Green Version]
- Ledran, M.H.; Krassowska, A.; Armstrong, L.; Dimmick, I.; Renström, J.; Lang, R.; Yung, S.; Santibanez-Coref, M.; Dzierzak, E.; Stojkovic, M.; et al. Efficient Hematopoietic Differentiation of Human Embryonic Stem Cells on Stromal Cells Derived from Hematopoietic Niches. Cell Stem Cell 2008, 3, 85–98. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, N.; Yamazaki, S.; Yamaguchi, T.; Okabe, M.; Masaki, H.; Takaki, S.; Otsu, M.; Nakauchi, H. Generation of Engraftable Hematopoietic Stem Cells from Induced Pluripotent Stem Cells by Way of Teratoma Formation. Mol. Ther. J. Am. Soc. Gene Ther. 2013, 21, 1424–1431. [Google Scholar] [CrossRef] [Green Version]
- Amabile, G.; Welner, R.S.; Nombela-Arrieta, C.; D’Alise, A.M.; Di Ruscio, A.; Ebralidze, A.K.; Kraytsberg, Y.; Ye, M.; Kocher, O.; Neuberg, D.S.; et al. In Vivo Generation of Transplantable Human Hematopoietic Cells from Induced Pluripotent Stem Cells. Blood 2013, 121, 1255–1264. [Google Scholar] [CrossRef]
- Pereira, C.-F.; Chang, B.; Qiu, J.; Niu, X.; Papatsenko, D.; Hendry, C.E.; Clark, N.R.; Nomura-Kitabayashi, A.; Kovacic, J.C.; Ma’ayan, A.; et al. Induction of a Hemogenic Program in Mouse Fibroblasts. Cell Stem Cell 2013, 13, 205–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riddell, J.; Gazit, R.; Garrison, B.S.; Guo, G.; Saadatpour, A.; Mandal, P.K.; Ebina, W.; Volchkov, P.; Yuan, G.-C.; Orkin, S.H.; et al. Reprogramming Committed Murine Blood Cells to Induced Hematopoietic Stem Cells with Defined Factors. Cell 2014, 157, 549–564. [Google Scholar] [CrossRef] [Green Version]
- Sandler, V.M.; Lis, R.; Liu, Y.; Kedem, A.; James, D.; Elemento, O.; Butler, J.M.; Scandura, J.M.; Rafii, S. Reprogramming Human Endothelial Cells to Haematopoietic Cells Requires Vascular Induction. Nature 2014, 511, 312–318. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Biological (Y/N)/BSL Level | Cat # |
|---|---|---|
| ERG (transcript variant 1) ORF clone | Y, level 1 | EX-Z1500-Lv165 |
| LCOR (transcript variant 3) ORF clone | Y, level 1 | EX-E2088-Lv165 |
| HOXA10 ORF clone | Y, level 1 | EX-Z5789-Lv165 |
| HOXA5 ORF clone | Y, level 1 | EX-F0180-Lv165 |
| HOXA9 ORF clone | Y, level 1 | EX-P0078-Lv165 |
| Antibody | Company | Cat # |
|---|---|---|
| Rb mAb to RUNX1 | Abcam | ab92336 |
| Rb mAb to PU.1/SPI1 | Abcam | ab76543 |
| Rb mAb to HOXA5 | Abcam | ab140636 |
| HOXA9 (rabbit polyclonal IgG) | EMD Millipore Corp | 07–178 |
| HOXA10 | Abcam | ab191470 |
| Rb mAb to ERG | Abcam | ab92513 |
| Rb mAb to LCOR | Abcam | ab171086 |
| Ms mAB to GAPDH | Abcam | ab8245 |
| Gene Name | Gene Forward (5′–3′) | Reverse (5′–3′) |
|---|---|---|
| RUNX1c | CGT ACC CAC AGT GCT TCA TGA | GGC ATC GTG GAC GTC TCT AGA |
| SPI1 | GCC AAA CGC ACG AGT ATT ACC | GGG TGG AAG TCC CAG TAA TGG |
| HOXA5 | ACC CCA GAT CTA CCC CTG GAT | CGG GCC GCC TAT GTT G |
| HOXA9 | ATG AGA GCG GCG GAG ACA | CCA GTT GGC TGC TGG GTT A |
| HOXA10 | AAA GCC TCG CCG GAG AA | GCC AGT TGG CTG CGT TTT |
| ERG | GAA CGA GCG CAG AGT TAT CGT | TGC CGC ACA TGG TCT GTA CT |
| LCOR | CTC AGT CAG AAC CTA GCG AAC AAG | GCC AGC ACA TGG ACT TTT CTT A |
| GAPDH | CCA CAT CGC TCA GAC ACC AT | GCG CCC AAT ACG ACC AAA T |
| Human DNA | Gene Forward (5′–3′) | Reverse (5′–3′) |
|---|---|---|
| alpha-satellite, chromosome 17 | GGGATAATTTCAGCTGACTAAACAG | TTCCGTTTAGTTAGGTGCAGTTATC |
| TRE3G-TF-FP | CTG GAG CAA TTC CAC AAC AC | |
| RUNX1c | CAA CGC CTC GCT CAT CTT | |
| SPI1 | GGA GCT CCG TGA AGT TGT TC | |
| HOXA5 | AGA TCC ATG CCA TTG TAG CC | |
| HOXA9 | CTT GGA CTG GAA GCT GCA C | |
| HOXA10 | CAG CTC TGC AGC CCG TAG | |
| ERGv1 | GTT CCT TGA GCC ATT CAC CT | |
| LCOR | GGT CCA GAG GTG AGT CTT GG |
| Antigen | Reactivity | Conjugated | Clone | Supplier |
|---|---|---|---|---|
| CD34 | Human | APC-Cy7 | 581 | BD Horizon Cat # 343514 |
| CD31 | Human | FITC | BD Horizon Cat # 555445 | |
| FLK1 | Human | APC | 89106 | BD Horizon Cat # 560495 |
| VE-cadherin | Human | PE-Cy7 | 16B1 | BD Horizon Cat # 25-1449-41 |
| CD45 | Human | V450 | 2D1 | BD Horizon Cat # 642275 |
| CD43 | Human | PE | 1G10 | BD Horizon Cat # 560199 |
| CD235a | Human | PE | GA-R2 | BD Horizon Cat # IM22114 |
| CD45 | Human | APC | 2D1 | BD Horizon Cat # 340943 |
| Lab Source | GEO ID | Symbol in Study | Samples Used | Sample IDs | Platform Technology | Total Annotated Genes |
|---|---|---|---|---|---|---|
| Bhatia | GSE3823 | circle | 18 | U133A; GSM87705 to GSM87716, GSM87729 to GSM87734 | HG U133A | 13,462 |
| Daley | GSE49938 | diamond | 17 | GSM1210379 to GSM1210384, GSM1210388 to GSM1210392, GSM1210401 to GSM121406 | HG U133A Plus2 | 23,520 |
| Daley | GSE83719 | triangle | 5 | All; GSM2214010 to GSM2299187 | Illumina NextSeq 500 | 25,855 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Porras, D.P.; Reid, J.C.; Tanasijevic, B.; Golubeva, D.; Boyd, A.L.; Bhatia, M. Challenges in Cell Fate Acquisition to Scid-Repopulating Activity from Hemogenic Endothelium of hiPSCs Derived from AML Patients Using Forced Transcription Factor Expression. Cells 2022, 11, 1915. https://doi.org/10.3390/cells11121915
Porras DP, Reid JC, Tanasijevic B, Golubeva D, Boyd AL, Bhatia M. Challenges in Cell Fate Acquisition to Scid-Repopulating Activity from Hemogenic Endothelium of hiPSCs Derived from AML Patients Using Forced Transcription Factor Expression. Cells. 2022; 11(12):1915. https://doi.org/10.3390/cells11121915
Chicago/Turabian StylePorras, Deanna P., Jennifer C. Reid, Borko Tanasijevic, Diana Golubeva, Allison L. Boyd, and Mickie Bhatia. 2022. "Challenges in Cell Fate Acquisition to Scid-Repopulating Activity from Hemogenic Endothelium of hiPSCs Derived from AML Patients Using Forced Transcription Factor Expression" Cells 11, no. 12: 1915. https://doi.org/10.3390/cells11121915
APA StylePorras, D. P., Reid, J. C., Tanasijevic, B., Golubeva, D., Boyd, A. L., & Bhatia, M. (2022). Challenges in Cell Fate Acquisition to Scid-Repopulating Activity from Hemogenic Endothelium of hiPSCs Derived from AML Patients Using Forced Transcription Factor Expression. Cells, 11(12), 1915. https://doi.org/10.3390/cells11121915