
Serum Cytokines Predict Neurological Damage in Genetically Diverse Mouse Models

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
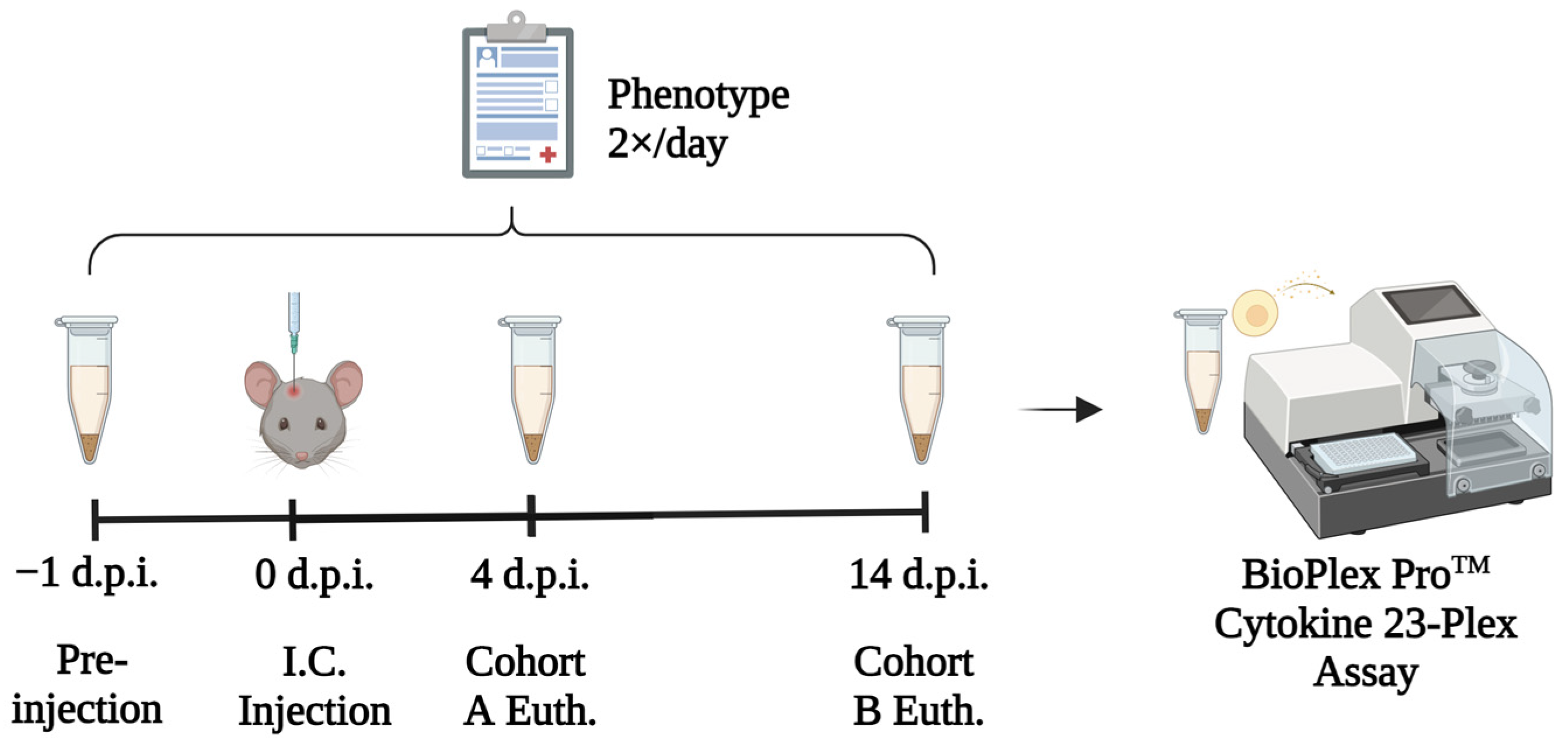
2.2. Experimental Design
2.3. Mouse Management
2.4. Qualitative Neurological Phenotyping
2.5. Serum Collection and Euthanasia
2.6. Cytokine and Chemokine Assays
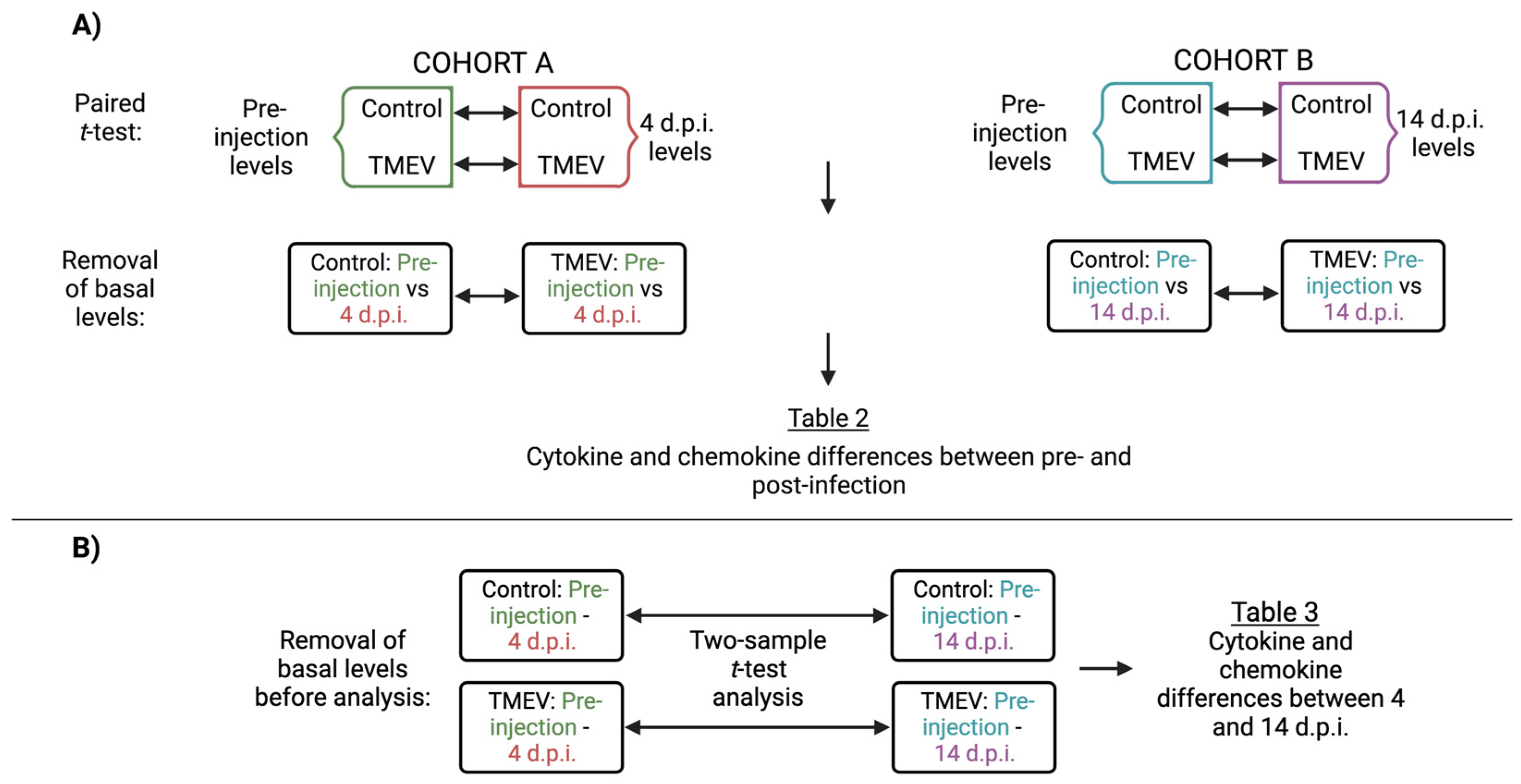
2.7. Statistics
3. Results
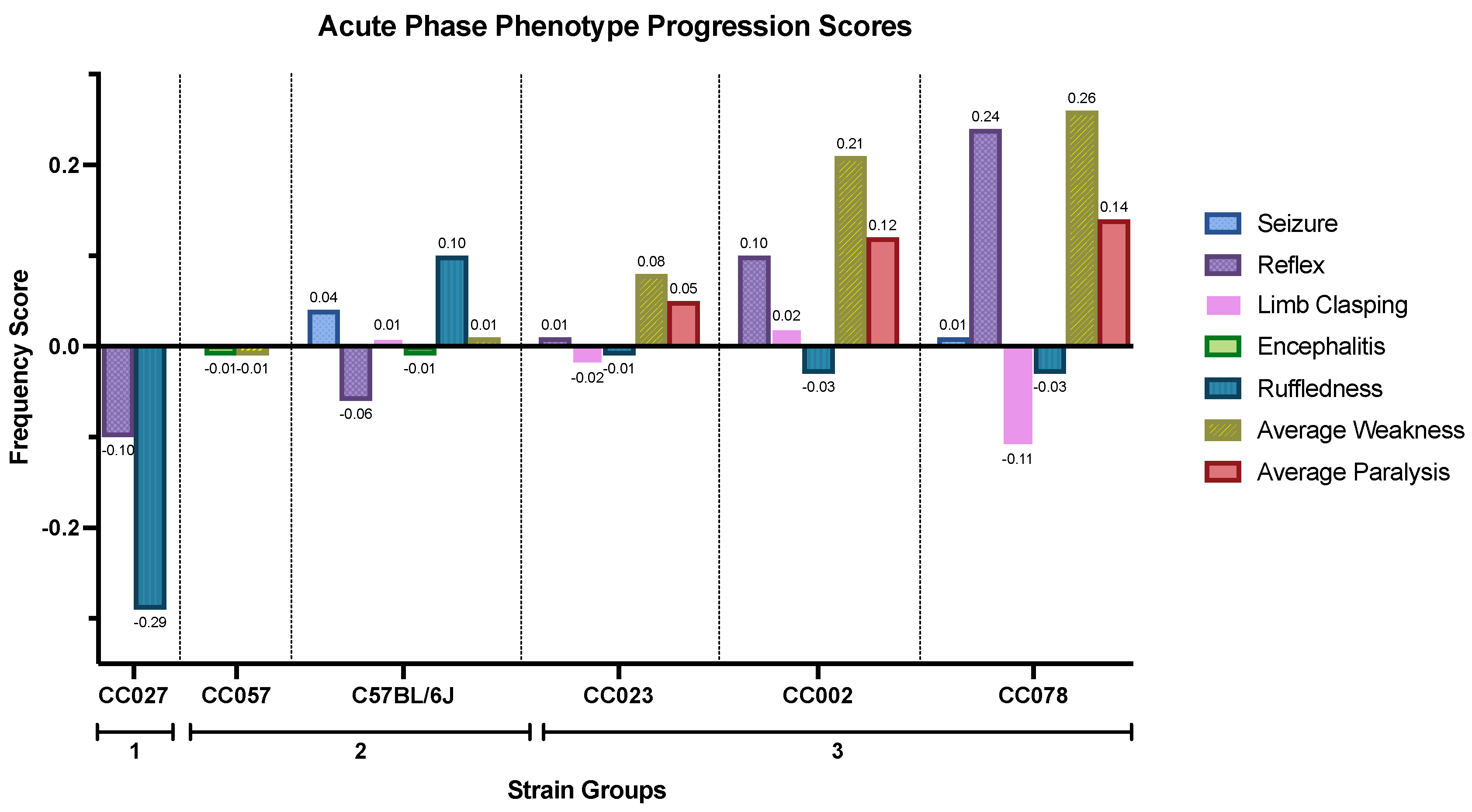
3.1. TMEV-Induced Phenotypes
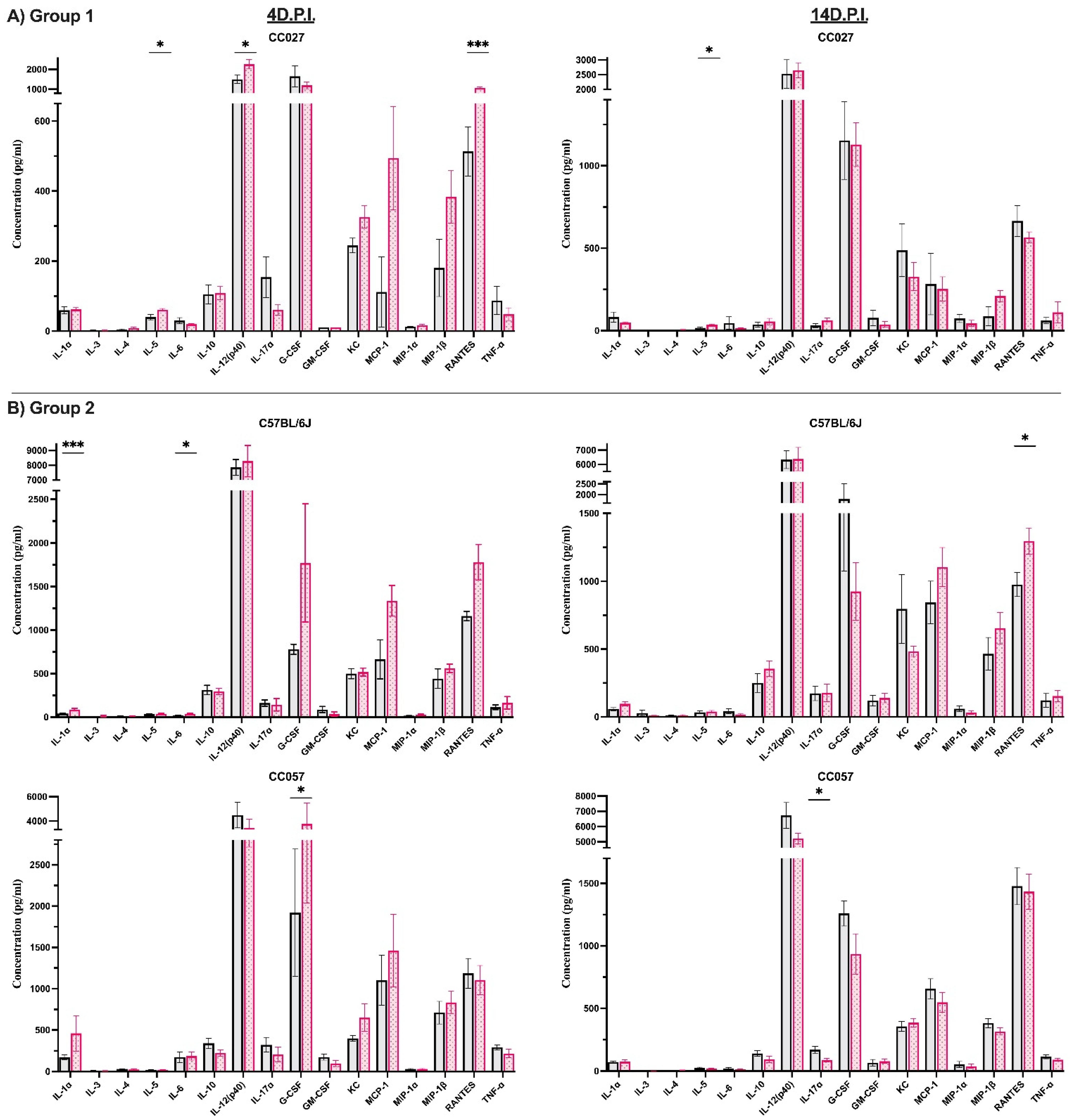
3.2. Identification of Immune Responses Influenced by Intracranial Injection
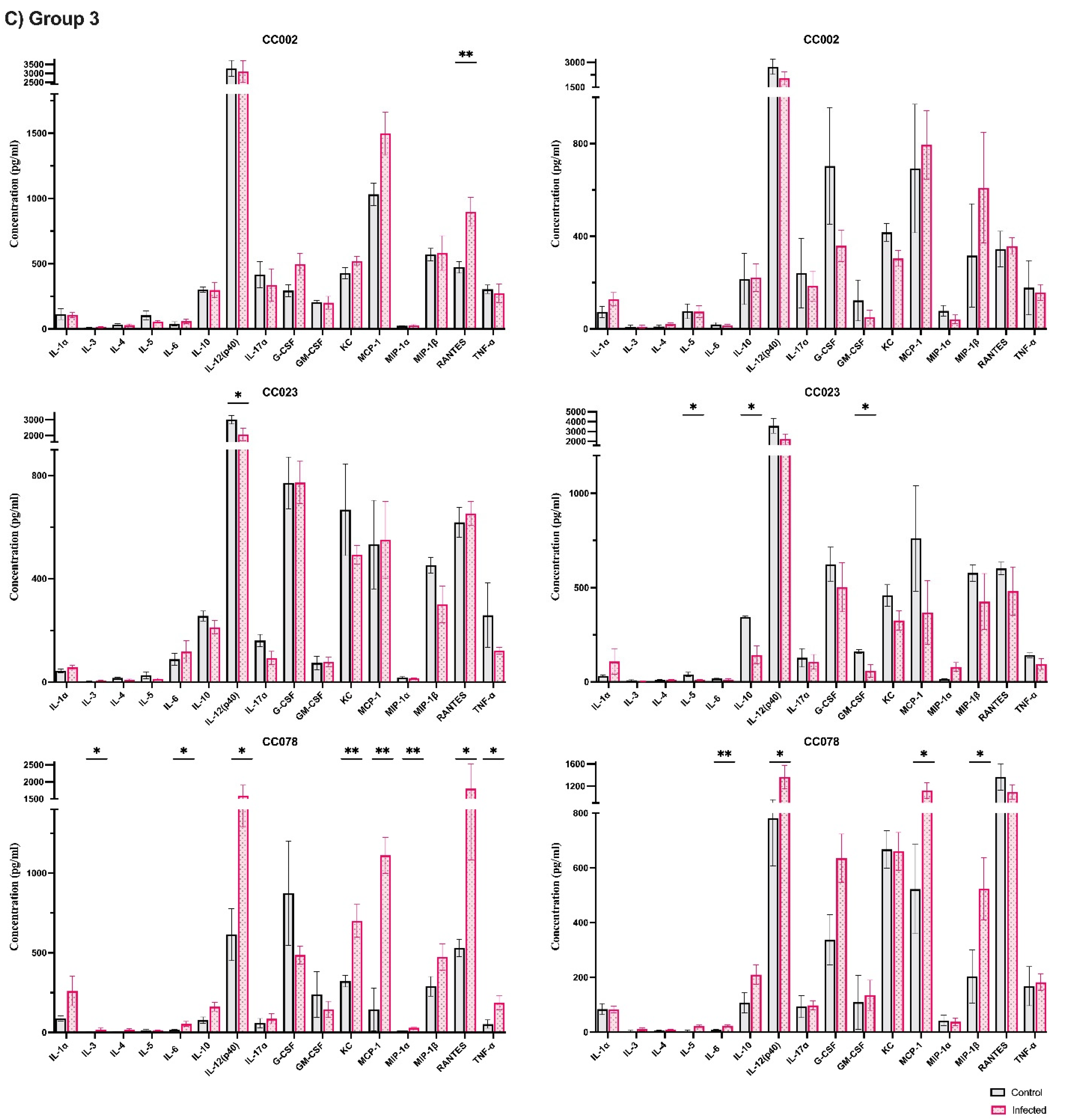
3.3. Characterization of Immune Responses Elicited Solely by TMEV Infection
3.4. Cytokine and Chemokine Influence on TMEV-Induced Phenotypes
3.5. Gender-Specific Differences Were Identified for Certain Cytokine and Chemokine Levels at 4 and 14 d.p.i.
3.6. Pre-Injection Serum Levels Serve as Predictive Markers for TMEV-Induced Phenotypes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, N.; Zuo, Y.; Jiang, L.; Peng, Y.; Huang, X.; Zuo, L. Epstein-Barr Virus and Neurological Diseases. Front. Mol. Biosci. 2022, 8, 1310. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, J.; Wen, Y. Lessons from Epstein-Barr virus DNA detection in cerebrospinal fluid as a diagnostic tool for EBV-induced central nervous system dysfunction among HIV-positive patients. Biomed. Pharmacother. 2022, 145, 112392. [Google Scholar] [CrossRef] [PubMed]
- Bello-Morales, R.; Andreu, S.; López-Guerrero, J.A. The Role of Herpes Simplex Virus Type 1 Infection in Demyelination of the Central Nervous System. Int. J. Mol. Sci. 2020, 21, 5026. [Google Scholar] [CrossRef] [PubMed]
- Jakhmola, S.; Upadhyay, A.; Jain, K.; Mishra, A.; Jha, H.C. Herpesviruses and the hidden links to Multiple Sclerosis neuropathology. J. Neuroimmunol. 2021, 358, 577636. [Google Scholar] [CrossRef] [PubMed]
- Ngarka, L.; Siewe Fodjo, J.N.; Aly, E.; Masocha, W.; Njamnshi, A.K. The Interplay Between Neuroinfections, the Immune System and Neurological Disorders: A Focus on Africa. Front. Immunol. 2022, 12, 5688. [Google Scholar] [CrossRef]
- Cheng, D.; Luo, Z.; Fu, X.; Stephenson, S.; Di Germanio, C.; Norris, P.J.; Fuchs, D.; Ndhlovu, L.C.; Li, Q.; Zetterberg, H.; et al. Elevated Cerebrospinal Fluid Anti-CD4 Autoantibody Levels in HIV Associate with Neuroinflammation. Microbiol. Spectr. 2022, 10, e01975-21. [Google Scholar] [CrossRef]
- Fontana, S.; Buttinelli, G.; Fiore, S.; Amato, C.; Pataracchia, M.; Kota, M.; Aćimović, J.; Blažević, M.; Mulaomerović, M.; Nikolaeva-Glomb, L.; et al. Retrospective Analysis of Six Years of Acute Flaccid Paralysis Surveillance and Polio Vaccine Coverage Reported by Italy, Serbia, Bosnia and Herzegovina, Montenegro, Bulgaria, Kosovo, Albania, North Macedonia, Malta, and Greece. Vaccines 2021, 10, 44. [Google Scholar] [CrossRef]
- Tseha, S.T. Polio: The Disease that Reemerged after Six Years in Ethiopia. Ethiop. J. Health Sci. 2021, 31, 897–902. [Google Scholar] [CrossRef]
- Xie, X.; Zeng, J. Neuroimmune Evasion of Zika Virus to Facilitate Viral Pathogenesis. Front. Cell. Infect. Microbiol. 2021, 11, 662447. [Google Scholar] [CrossRef]
- Lee, L.J.; Komarasamy, T.V.; Adnan, N.A.A.; James, W.; RMT Balasubramaniam, V. Hide and Seek: The Interplay Between Zika Virus and the Host Immune Response. Front. Immunol. 2021, 12, 4379. [Google Scholar] [CrossRef]
- Leon, L.L.; de Lima, R.G.; Boffi, L.C.; Bindilatti, R.N.; Garlipp, C.R.; Costa, S.C.B.; Bonon, S.H.A. Arbovirus, herpesvirus, and enterovirus associated with neurological syndromes in adult patients of a university hospital, 2017–2018. Rev. Soc. Bras. Med. Trop. 2021, 54, e0127. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.C.; Feuer, R.; Cashman, N.; Luo, H. Enteroviral Infection: The Forgotten Link to Amyotrophic Lateral Sclerosis? Front. Mol. Neurosci. 2018, 11, 63. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.; Ingre, C.; Roos, P.; Kamel, F.; Piehl, F. Risk factors for amyotrophic lateral sclerosis. Clin. Epidemiol. 2015, 7, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michael, B.D.; Solomon, T. Seizures and encephalitis: Clinical features, management, and potential pathophysiologic mechanisms. Epilepsia 2012, 53, 63–71. [Google Scholar] [CrossRef]
- Libbey, J.E.; Fujinami, R.S. Neurotropic viral infections leading to epilepsy: Focus on Theiler’s murine encephalomyelitis virus. Future Virol. 2011, 6, 1339–1350. [Google Scholar] [CrossRef] [Green Version]
- Bjornevik, K.; Cortese, M.; Healy, B.C.; Kuhle, J.; Mina, M.J.; Leng, Y.; Elledge, S.J.; Niebuhr, D.W.; Scher, A.I.; Munger, K.L.; et al. Longitudinal analysis reveals high prevalence of Epstein-Barr virus associated with multiple sclerosis. Science 2022, 375, 296–301. [Google Scholar] [CrossRef]
- Steelman, A.J. Infection as an Environmental Trigger of Multiple Sclerosis Disease Exacerbation. Front. Immunol. 2015, 6, 520. [Google Scholar] [CrossRef] [Green Version]
- Caggiu, E.; Arru, G.; Hosseini, S.; Niegowska, M.; Sechi, G.; Zarbo, I.R.; Sechi, L.A. Inflammation, Infectious Triggers, and Parkinson’s Disease. Front. Neurol. 2019, 10, 122. [Google Scholar] [CrossRef] [Green Version]
- Woulfe, J.M.; Gray, M.T.; Gray, D.A.; Munoz, D.G.; Middeldorp, J.M. Hypothesis: A role for EBV-induced molecular mimicry in Parkinson’s disease. Parkinsonism Relat. Disord. 2014, 20, 685–694. [Google Scholar] [CrossRef]
- Lipton, H.L. Theiler’s virus infection in mice: An unusual biphasic disease process leading to demyelination. Infect. Immun. 1975, 11, 1147–1155. [Google Scholar] [CrossRef] [Green Version]
- Lipton, H.L. Persistent Theiler’s Murine Encephalomyelitis Virus Infection in Mice depends on Plaque Size. J. Gen. Virol. 1980, 46, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Libbey, J.E.; Fujinami, R.S. Viral mouse models used to study multiple sclerosis: Past and present. Arch. Virol. 2021, 166, 1015–1033. [Google Scholar] [CrossRef] [PubMed]
- DePaula-Silva, A.B.; Hanak, T.J.; Libbey, J.E.; Fujinami, R.S. Theiler’s murine encephalomyelitis virus infection of SJL/J and C57BL/6J mice: Models for multiple sclerosis and epilepsy. J. Neuroimmunol. 2017, 308, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Gerhauser, I.; Hansmann, F.; Ciurkiewicz, M.; Löscher, W.; Beineke, A. Facets of Theiler’s Murine Encephalomyelitis Virus-Induced Diseases: An Update. Int. J. Mol. Sci. 2019, 20, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez Gomez, A.A.; Karmakar, M.; Carroll, R.J.; Lawley, K.S.; Amstalden, K.; Young, C.R.; Threadgill, D.W.; Welsh, C.J.; Brinkmeyer-Langford, C. Genetic and immunological contributors to virus-induced paralysis. Brain Behav. Immun. Health 2021, 18, 100395. [Google Scholar] [CrossRef]
- Kim, B.S. Excessive Innate Immunity Steers Pathogenic Adaptive Immunity in the Development of Theiler’s Virus-Induced Demyelinating Disease. Int. J. Mol. Sci. 2021, 22, 5254. [Google Scholar] [CrossRef]
- Graves, M.C.; Bologa, L.; Siegel, L.; Londe, H. Theiler’s virus in brain cell cultures: Lysis of neurons and oligodendrocytes and persistence in astrocytes and macrophages. J. Neurosci. Res. 1986, 15, 491–501. [Google Scholar] [CrossRef]
- Welsh, C.J.; Sapatino, B.V.; Rosenbaum, B.A.; Smith, R. Characteristics of cloned cerebrovascular endothelial cells following infection with Theiler’s virus. I. Acute infection. J. Neuroimmunol. 1995, 62, 119–125. [Google Scholar] [CrossRef]
- Aubert, C.; Chamorro, M.; Brahic, M. Identification of Theiler’s virus infected cells in the central nervous system of the mouse during demyelinating disease. Microb. Pathog. 1987, 3, 319–326. [Google Scholar] [CrossRef]
- Lipton, H.L.; Twaddle, G.; Jelachich, M.L. The predominant virus antigen burden is present in macrophages in Theiler’s murine encephalomyelitis virus-induced demyelinating disease. J. Virol. 1995, 69, 2525–2533. [Google Scholar] [CrossRef] [Green Version]
- Oleszak, E.L.; Chang, J.R.; Friedman, H.; Katsetos, C.D.; Platsoucas, C.D. Theiler’s Virus Infection: A Model for Multiple Sclerosis. Clin. Microbiol. Rev. 2004, 17, 174–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trottier, M.; Schlitt, B.P.; Kung, A.Y.; Lipton, H.L. Transition from acute to persistent Theiler’s virus infection requires active viral replication that drives proinflammatory cytokine expression and chronic demyelinating disease. J. Virol. 2004, 78, 12480–12488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mecha, M.; Carrillo-Salinas, F.J.; Mestre, L.; Feliú, A.; Guaza, C. Viral models of multiple sclerosis: Neurodegeneration and demyelination in mice infected with Theiler’s virus. Prog. Neurobiol. 2013, 101–102, 46–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Churchill, G.A.; Airey, D.C.; Allayee, H.; Angel, J.M.; Attie, A.D.; Beatty, J.; Beavis, W.D.; Belknap, J.K.; Bennett, B.; Berrettini, W.; et al. The Collaborative Cross, a community resource for the genetic analysis of complex traits. Nat. Genet. 2004, 36, 1133–1137. [Google Scholar] [CrossRef] [PubMed]
- Threadgill, D.W.; Miller, D.R.; Churchill, G.A.; de Villena, F.P.-M. The Collaborative Cross: A Recombinant Inbred Mouse Population for the Systems Genetic Era. ILAR J. 2011, 52, 24–31. [Google Scholar] [CrossRef]
- Threadgill, D.W.; Churchill, G.A. Ten years of the Collaborative Cross. Genetics 2012, 190, 291–294. [Google Scholar] [CrossRef] [Green Version]
- Welsh, C.J.R.; Tonks, P.; Nash, A.A.; Blakemore, W.F. The Effect of L3T4 T Cell Depletion on the Pathogenesis of Theiler’s Murine Encephalomyelitis Virus Infection in CBA Mice. J. Gen. Virol. 1987, 68, 1659–1667. [Google Scholar] [CrossRef]
- Eldridge, R.; Osorio, D.; Amstalden, K.; Edwards, C.; Young, C.R.; Cai, J.J.; Konganti, K.; Hillhouse, A.; Threadgill, D.W.; Welsh, C.J.; et al. Antecedent presentation of neurological phenotypes in the Collaborative Cross reveals four classes with complex sex-dependencies. Sci. Rep. 2020, 10, 7918. [Google Scholar] [CrossRef]
- Lawley, K.S.; Rech, R.R.; Elenwa, F.; Han, G.; Perez Gomez, A.A.; Amstalden, K.; Welsh, C.J.; Young, C.R.; Threadgill, D.W.; Brinkmeyer-Langford, C.L. Host genetic diversity drives variable central nervous system lesion distribution in chronic phase of Theiler’s Murine Encephalomyelitis Virus (TMEV) infection. PLoS ONE 2021, 16, e0256370. [Google Scholar] [CrossRef]
- Brinkmeyer-Langford, C.L.; Rech, R.; Amstalden, K.; Kochan, K.J.; Hillhouse, A.E.; Young, C.; Welsh, C.J.; Threadgill, D.W. Host genetic background influences diverse neurological responses to viral infection in mice. Sci. Rep. 2017, 7, 12194. [Google Scholar] [CrossRef] [Green Version]
- Sieve, A.N.; Steelman, A.J.; Young, C.R.; Storts, R.; Welsh, T.H.; Welsh, C.J.R.; Meagher, M.W. Chronic restraint stress during early Theiler’s virus infection exacerbates the subsequent demyelinating disease in SJL mice. J. Neuroimmunol. 2004, 155, 103–118. [Google Scholar] [CrossRef] [PubMed]
- Campbell, T.; Meagher, M.W.; Sieve, A.; Scott, B.; Storts, R.; Welsh, T.H.; Welsh, C.J.R. The Effects of Restraint Stress on the Neuropathogenesis of Theiler’s Virus Infection: I. Acute Disease. Brain Behav. Immun. 2001, 15, 235–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meagher, M.W.; Sieve, A.N.; Johnson, R.R.; Satterlee, D.; Belyavskyi, M.; Mi, W.; Prentice, T.W.; Welsh, T.H.; Welsh, C.J.R. Neonatal Maternal Separation Alters Immune, Endocrine, and Behavioral Responses to Acute Theiler’s Virus Infection in Adult Mice. Behav. Genet. 2010, 40, 233–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welsh, C.J.R.; Bustamante, L.; Nayak, M.; Welsh, T.H.; Dean, D.D.; Meagher, M.W. The effects of restraint stress on the neuropathogenesis of Theiler’s virus infection II: NK cell function and cytokine levels in acute disease. Brain Behav. Immun. 2004, 18, 166–174. [Google Scholar] [CrossRef]
- Pappas, S.S.; Darr, K.; Holley, S.M.; Cepeda, C.; Mabrouk, O.S.; Wong, J.-M.T.; LeWitt, T.M.; Paudel, R.; Houlden, H.; Kennedy, R.T.; et al. Forebrain deletion of the dystonia protein torsinA causes dystonic-like movements and loss of striatal cholinergic neurons. eLife 2015, 4, e08352. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Huo, X.; Cao, Y.-F.; Li, S.-N.; Du, Z.; Shao, P.; Leng, J.; Zhang, C.; Sun, X.-Y.; Ma, R.C.W.; et al. Bile acid metabolites in early pregnancy and risk of gestational diabetes in Chinese women: A nested case-control study. EBioMedicine 2018, 35, 317–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkmeyer-Langford, C.; Amstalden, K.; Konganti, K.; Hillhouse, A.; Lawley, K.; Perez-Gomez, A.; Young, C.R.; Welsh, C.J.; Threadgill, D.W. Resilience in Long-Term Viral Infection: Genetic Determinants and Interactions. Int. J. Mol. Sci. 2021, 22, 11379. [Google Scholar] [CrossRef]
- Lipton, H.L.; Melvold, R. Genetic analysis of susceptibility to Theiler’s virus-induced demyelinating disease in mice. J. Immunol. 1984, 132, 1821–1825. [Google Scholar]
- Clatch, R.J.; Melvold, R.W.; Miller, S.D.; Lipton, H.L. Theiler’s murine encephalomyelitis virus (TMEV)-induced demyelinating disease in mice is influenced by the H-2D region: Correlation with TEMV-specific delayed-type hypersensitivity. J. Immunol. 1985, 135, 1408–1414. [Google Scholar]
- Woodcock, T.; Morganti-Kossmann, M.C. The Role of Markers of Inflammation in Traumatic Brain Injury. Front. Neurol. 2013, 4, 18. [Google Scholar] [CrossRef] [Green Version]
- Arvin, B.; Neville, L.F.; Barone, F.C.; Feuerstein, G.Z. The Role of Inflammation and Cytokines in Brain Injury. Neurosci. Biobehav. Rev. 1996, 20, 445–452. [Google Scholar] [CrossRef]
- Janicki-Deverts, D.; Cohen, S.; Doyle, W.J.; Turner, R.B.; Treanor, J.J. Infection-induced proinflammatory cytokines are associated with decreases in positive affect, but not increases in negative affect. Brain Behav. Immun. 2007, 21, 301–307. [Google Scholar] [CrossRef] [Green Version]
- Ngcobo, S.; Molatlhegi, R.P.; Osman, F.; Ngcapu, S.; Samsunder, N.; Garrett, N.J.; Abdool Karim, S.S.; Abdool Karim, Q.; McKinnon, L.R.; Sivro, A. Pre-infection plasma cytokines and chemokines as predictors of HIV disease progression. Sci. Rep. 2022, 12, 2437. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lipton, H.L.; Perelson, A.S.; Dahari, H. Modeling the acute and chronic phases of Theiler murine encephalomyelitis virus infection. J. Virol. 2013, 87, 4052–4059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, J.R.; Zaczynska, E.; Katsetos, C.D.; Platsoucas, C.D.; Oleszak, E.L. Differential Expression of TGF-β, IL-2, and Other Cytokines in the CNS of Theiler’s Murine Encephalomyelitis Virus-Infected Susceptible and Resistant Strains of Mice. Virology 2000, 278, 346–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pachner, A.R.; Li, L.; Gilli, F. Chemokine biomarkers in central nervous system tissue and cerebrospinal fluid in the Theiler’s virus model mirror those in multiple sclerosis. Cytokine 2015, 76, 577–580. [Google Scholar] [CrossRef]
- Hale, K.D.; Ghanta, V.K.; Gauthier, D.K.; Hiramoto, R.N. Effects of Rotational Stress of Different Duration on NK Cell Activity, Proinflammatory Cytokines, and POMC-Derived Peptides in Mice. Neuroimmunomodulation 2001, 9, 34–40. [Google Scholar] [CrossRef]
- Kim, B.S.; Jin, Y.-H.; Meng, L.; Hou, W.; Kang, H.S.; Park, H.S.; Koh, C.-S. IL-1 signal affects both protection and pathogenesis of virus-induced chronic CNS demyelinating disease. J. Neuroinflamm. 2012, 9, 217. [Google Scholar] [CrossRef] [Green Version]
- Takatsu, K. Interleukin-5 and IL-5 receptor in health and diseases. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2011, 87, 463–485. [Google Scholar] [CrossRef] [Green Version]
- Cooper, A.M.; Khader, S.A. IL-12p40: An inherently agonistic cytokine. Trends Immunol. 2007, 28, 33–38. [Google Scholar] [CrossRef]
- Mikolajczyk, T.P.; Nosalski, R.; Szczepaniak, P.; Budzyn, K.; Osmenda, G.; Skiba, D.; Sagan, A.; Wu, J.; Vinh, A.; Marvar, P.J.; et al. Role of chemokine RANTES in the regulation of perivascular inflammation, T-cell accumulation, and vascular dysfunction in hypertension. FASEB J. 2016, 30, 1987–1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Chow, J.M.; Gri, G.; Carra, G.; Gerosa, F.; Wolf, S.F.; Dzialo, R.; Trinchieri, G. The interleukin 12 p40 gene promoter is primed by interferon gamma in monocytic cells. J. Exp. Med. 1996, 183, 147–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapil, P.; Atkinson, R.; Ramakrishna, C.; Cua, D.J.; Bergmann, C.C.; Stohlman, S.A. Interleukin-12 (IL-12), but not IL-23, deficiency ameliorates viral encephalitis without affecting viral control. J. Virol. 2009, 83, 5978–5986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glass, W.G.; Lim, J.K.; Cholera, R.; Pletnev, A.G.; Gao, J.-L.; Murphy, P.M. Chemokine receptor CCR5 promotes leukocyte trafficking to the brain and survival in West Nile virus infection. J. Exp. Med. 2005, 202, 1087–1098. [Google Scholar] [CrossRef] [Green Version]
- Kouro, T.; Takatsu, K. IL-5-and eosinophil-mediated inflammation: From discovery to therapy. Int. Immunol. 2009, 21, 1303–1309. [Google Scholar] [CrossRef] [Green Version]
- Gadani, S.P.; Cronk, J.C.; Norris, G.T.; Kipnis, J. IL-4 in the brain: A cytokine to remember. J. Immunol. 2012, 189, 4213–4219. [Google Scholar] [CrossRef]
- Lin, S.-J.; Shu, P.-Y.; Chang, C.; Ng, A.-K.; Hu, C. IL-4 suppresses the expression and the replication of hepatitis B virus in the hepatocellular carcinoma cell line Hep3B. J. Immunol. 2003, 171, 4708–4716. [Google Scholar] [CrossRef]
- Gorski, S.A.; Hahn, Y.S.; Braciale, T.J. Group 2 innate lymphoid cell production of IL-5 is regulated by NKT cells during influenza virus infection. PLoS Pathog. 2013, 9, e1003615. [Google Scholar] [CrossRef]
- Butti, E.; Bergami, A.; Recchia, A.; Brambilla, E.; Del Carro, U.; Amadio, S.; Cattalini, A.; Esposito, M.; Stornaiuolo, A.; Comi, G.; et al. IL4 gene delivery to the CNS recruits regulatory T cells and induces clinical recovery in mouse models of multiple sclerosis. Gene Ther. 2008, 15, 504–515. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, J.; Gong, Y.; Gu, Y.; Xiang, Q.; Tang, L.-L. Interleukin-6 and granulocyte colony-stimulating factor as predictors of the prognosis of influenza-associated pneumonia. BMC Infect. Dis. 2022, 22, 343. [Google Scholar] [CrossRef]
- Semerad, C.L.; Liu, F.; Gregory, A.D.; Stumpf, K.; Link, D.C. G-CSF is an essential regulator of neutrophil trafficking from the bone marrow to the blood. Immunity 2002, 17, 413–423. [Google Scholar] [CrossRef] [Green Version]
- Veldhoen, M. Interleukin 17 is a chief orchestrator of immunity. Nat. Immunol. 2017, 18, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Sahu, U.; Biswas, D.; Prajapati, V.K.; Singh, A.K.; Samant, M.; Khare, P. Interleukin-17-A multifaceted cytokine in viral infections. J. Cell. Physiol. 2021, 236, 8000–8019. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.-T.; Yao, X.-T.; Peng, Q.; Chen, D.-K. The protective and pathogenic roles of IL-17 in viral infections: Friend or foe? Open Biol. 2019, 9, 190109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedrossian, N.; Haidar, M.; Fares, J.; Kobeissy, F.H.; Fares, Y. Inflammation and Elevation of Interleukin-12p40 in Patients with Schizophrenia. Front. Mol. Neurosci. 2016, 9, 16. [Google Scholar] [CrossRef]
- Eede, P.; Obst, J.; Benke, E.; Yvon-Durocher, G.; Richard, B.C.; Gimber, N.; Schmoranzer, J.; Böddrich, A.; Wanker, E.E.; Prokop, S.; et al. Interleukin-12/23 deficiency differentially affects pathology in male and female Alzheimer’s disease-like mice. EMBO Rep. 2020, 21, e48530. [Google Scholar] [CrossRef]
- Erickson, M.A.; Liang, W.S.; Fernandez, E.G.; Bullock, K.M.; Thysell, J.A.; Banks, W.A. Genetics and sex influence peripheral and central innate immune responses and blood-brain barrier integrity. PLoS ONE 2018, 13, e0205769. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, K.W.; Lee, J.J.; Foster, G.A.; Krysztof, D.; Stramer, S.L.; Lim, J.K. Sex differences in cytokine production following West Nile virus infection: Implications for symptom manifestation. Pathog. Dis. 2019, 77, ftz016. [Google Scholar] [CrossRef] [Green Version]
- Targowski, T.; Jahnz-Rózyk, K.; Plusa, T.; Glodzinska-Wyszogrodzka, E. Influence of age and gender on serum eotaxin concentration in healthy and allergic people. J. Investig. Allergol. Clin. Immunol. 2005, 15, 277–282. [Google Scholar]
- Pokorna Formanova, P.; Palus, M.; Salat, J.; Hönig, V.; Stefanik, M.; Svoboda, P.; Ruzek, D. Changes in cytokine and chemokine profiles in mouse serum and brain, and in human neural cells, upon tick-borne encephalitis virus infection. J. Neuroinflamm. 2019, 16, 205. [Google Scholar] [CrossRef] [Green Version]
- Pavelko, K.D.; Howe, C.L.; Drescher, K.M.; Gamez, J.D.; Johnson, A.J.; Wei, T.; Ransohoff, R.M.; Rodriguez, M. Interleukin-6 protects anterior horn neurons from lethal virus-induced injury. J. Neurosci. 2003, 23, 481–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinman, L. Assessment of Animal Models Minireview for MS and Demyelinating Disease in the Design of Rational Therapy. Neuron 1999, 24, 511–514. [Google Scholar] [CrossRef] [Green Version]
- Meyaard, L.; Hovenkamp, E.; Pakker, N.; van der Pouw Kraan, T.C.T.M.; Miedema, F. Interleukin-12 (IL-12) Production in Whole Blood Cultures from Human Immunodeficiency Virus-Infected Individuals Studied in Relation to IL-10 and Prostaglandin E2 Production. Blood 1997, 89, 570–576. [Google Scholar] [CrossRef] [PubMed]
- Diagbouga, S.; Aldebert, D.; Fumoux, F.; Capron, M.; Ledru, E. Relationship Between Interleukin-5 Production and Variations in Eosinophil Counts During HIV Infection in West Africa: Influence of Mycobacterium tuberculosis Infection. Scand. J. Immunol. 1999, 49, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Resino, S.; Sanchez-Ramon, S.; Bellon, J.M.; Jimenez, J.L.; Munoz-Fernandez, M.A. Impaired Interleukin-5 Production by T Cells as a Prognostic Marker of Disease Progression in HIV-1-Infected Children; John Libbey Eurotext Ltd.: Arcueil, France, 2001; Volume 12. [Google Scholar]
- Shi, Y.; Liu, C.H.; Roberts, A.I.; Das, J.; Xu, G.; Ren, G.; Zhang, Y.; Zhang, L.; Yuan, Z.R.; Tan, H.S.W.; et al. Granulocyte-macrophage colony-stimulating factor (GM-CSF) and T-cell responses: What we do and don’t know. Cell Res. 2006, 16, 126–133. [Google Scholar] [CrossRef]
- Tasca, K.I.; Calvi, S.A.; de Souza, L.D.R. Immunovirological parameters and cytokines in HIV infection. Rev. Soc. Bras. Med. Trop. 2012, 45, 663–669. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, S.J.; Nelson, G.W. Human genes that limit AIDS. Nat. Genet. 2004, 36, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Modi, W.S.; Goedert, J.J.; Strathdee, S.; Buchbinder, S.; Detels, R.; Donfield, S.; O’Brien, S.J.; Winkler, C. MCP-1-MCP-3-Eotaxin gene cluster influences HIV-1 transmission. AIDS 2003, 17, 2357–2365. [Google Scholar] [CrossRef]
- Lee, P.W.; Xin, M.K.; Pei, W.; Yang, Y.; Lovett-Racke, A.E. IL-3 Is a Marker of Encephalitogenic T Cells, but Not Essential for CNS Autoimmunity. Front. Immunol. 2018, 9, 1255. [Google Scholar] [CrossRef]
- Renner, K.; Hellerbrand, S.; Hermann, F.; Riedhammer, C.; Talke, Y.; Schiechl, G.; Gomez, M.R.; Kutzi, S.; Halbritter, D.; Goebel, N.; et al. IL-3 promotes the development of experimental autoimmune encephalitis. JCI Insight 2016, 1, e87157. [Google Scholar] [CrossRef] [Green Version]
- Cua, D.J.; Sherlock, J.; Chen, Y.; Murphy, C.A.; Joyce, B.; Seymour, B.; Lucian, L.; To, W.; Kwan, S.; Churakova, T.; et al. Interleukin-23 rather than interleukin-12 is the critical cytokine for autoimmune inflammation of the brain. Nature 2003, 421, 744–748. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; He, C.; Nair, L.; Yeung, J.; Egwuagu, C.E. Interleukin 12 (IL-12) family cytokines: Role in immune pathogenesis and treatment of CNS autoimmune disease. Cytokine 2015, 75, 249–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michael, B.D.; Bricio-Moreno, L.; Sorensen, E.W.; Miyabe, Y.; Lian, J.; Solomon, T.; Kurt-Jones, E.A.; Luster, A.D. Astrocyte-and Neuron-Derived CXCL1 Drives Neutrophil Transmigration and Blood-Brain Barrier Permeability in Viral Encephalitis. Cell Rep. 2020, 32, 108150. [Google Scholar] [CrossRef] [PubMed]
- Alkhatib, G.; Locati, M.; Kennedy, P.E.; Murphy, P.M.; Berger, E.A. HIV-1 coreceptor activity of CCR5 and its inhibition by chemokines: Independence from G protein signaling and importance of coreceptor downmodulation. Virology 1997, 234, 340–348. [Google Scholar] [CrossRef] [Green Version]
- Cocchi, F.; DeVico, A.L.; Yarchoan, R.; Redfield, R.; Cleghorn, F.; Blattner, W.A.; Garzino-Demo, A.; Colombini-Hatch, S.; Margolis, D.; Gallo, R.C. Higher macrophage inflammatory protein (MIP)-1alpha and MIP-1beta levels from CD8+ T cells are associated with asymptomatic HIV-1 infection. Proc. Natl. Acad. Sci. USA 2000, 97, 13812–13817. [Google Scholar] [CrossRef] [Green Version]
- Fresegna, D.; Bullitta, S.; Musella, A.; Rizzo, F.R.; De Vito, F.; Guadalupi, L.; Caioli, S.; Balletta, S.; Sanna, K.; Dolcetti, E.; et al. Re-Examining the Role of TNF in MS Pathogenesis and Therapy. Cells 2020, 9, 2290. [Google Scholar] [CrossRef]
- Jin, Y.-H.; Mohindru, M.; Kang, M.H.; Fuller, A.C.; Kang, B.; Gallo, D.; Kim, B.S. Differential virus replication, cytokine production, and antigen-presenting function by microglia from susceptible and resistant mice infected with Theiler’s virus. J. Virol. 2007, 81, 11690–11702. [Google Scholar] [CrossRef] [Green Version]
- Moore, T.C.; Bush, K.L.; Cody, L.; Brown, D.M.; Petro, T.M. Control of early Theiler’s murine encephalomyelitis virus replication in macrophages by interleukin-6 occurs in conjunction with STAT1 activation and nitric oxide production. J. Virol. 2012, 86, 10841–10851. [Google Scholar] [CrossRef] [Green Version]
- Koper-Lenkiewicz, O.M.; Kamińska, J.; Lisowska, A.; Milewska, A.; Hirnle, T.; Dymicka-Piekarska, V. Factors Associated with RANTES Concentration in Cardiovascular Disease Patients. Biomed Res. Int. 2019, 2019, 3026453. [Google Scholar] [CrossRef]
- Fonteh, C.N.; Palestine, A.G.; Wagner, B.D.; Patnaik, J.L.; Mathias, M.T.; Mandava, N.; Baldermann, R.; Lynch, A.M.; University of Colorado Retina Research Group. Sex Differences in RANTES (CCL5) in Patients with Intermediate Age-Related Macular Degeneration (AMD) and Controls With no AMD. Transl. Vis. Sci. Technol. 2022, 11, 12. [Google Scholar] [CrossRef]
- Mulchandani, R.; Lyngdoh, T.; Kakkar, A.K. Deciphering the COVID-19 cytokine storm: Systematic review and meta-analysis. Eur. J. Clin. Investig. 2021, 51, e13429. [Google Scholar] [CrossRef] [PubMed]
- Ragab, D.; Salah Eldin, H.; Taeimah, M.; Khattab, R.; Salem, R. The COVID-19 Cytokine Storm; What We Know So Far. Front. Immunol. 2020, 11, 1446. [Google Scholar] [CrossRef] [PubMed]
- Stacey, A.R.; Norris, P.J.; Qin, L.; Haygreen, E.A.; Taylor, E.; Heitman, J.; Lebedeva, M.; DeCamp, A.; Li, D.; Grove, D.; et al. Induction of a Striking Systemic Cytokine Cascade prior to Peak Viremia in Acute Human Immunodeficiency Virus Type 1 Infection, in Contrast to More Modest and Delayed Responses in Acute Hepatitis B and C Virus Infections. J. Virol. 2009, 83, 3719–3733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muema, D.M.; Akilimali, N.A.; Ndumnego, O.C.; Rasehlo, S.S.; Durgiah, R.; Ojwach, D.B.A.; Ismail, N.; Dong, M.; Moodley, A.; Dong, K.L.; et al. Association between the cytokine storm, immune cell dynamics, and viral replicative capacity in hyperacute HIV infection. BMC Med. 2020, 18, 81. [Google Scholar] [CrossRef] [Green Version]
- Noelle, R.J.; Nowak, E.C. Cellular sources and immune functions of interleukin-9. Nat. Rev. Immunol. 2010, 10, 683–687. [Google Scholar] [CrossRef]
- Ding, X.; Cao, F.; Cui, L.; Ciric, B.; Zhang, G.-X.; Rostami, A. IL-9 signaling affects central nervous system resident cells during inflammatory stimuli. Exp. Mol. Pathol. 2015, 99, 570–574. [Google Scholar] [CrossRef]
- Costanza, M.; Colombo, M.P.; Pedotti, R. Mast cells in the pathogenesis of multiple sclerosis and experimental autoimmune encephalomyelitis. Int. J. Mol. Sci. 2012, 13, 15107–15125. [Google Scholar] [CrossRef] [Green Version]
- Conti, P.; Kempuraj, D. Important role of mast cells in multiple sclerosis. Mult. Scler. Relat. Disord. 2016, 5, 77–80. [Google Scholar] [CrossRef]
- Hendriksen, E.; van Bergeijk, D.; Oosting, R.S.; Redegeld, F.A. Mast cells in neuroinflammation and brain disorders. Neurosci. Biobehav. Rev. 2017, 79, 119–133. [Google Scholar] [CrossRef]
- Monastero, R.N.; Pentyala, S. Cytokines as Biomarkers and Their Respective Clinical Cutoff Levels. Int. J. Inflam. 2017, 2017, 4309485. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Complete Mouse List | ||||||
|---|---|---|---|---|---|---|
| Strain | Gender | Cohort A | Cohort B | Total | ||
| PBS | TMEV | PBS | TMEV | |||
| C57BL/6J | F | 3 | 4 | 4 | 4 | 15 |
| M | 3 | 4 | 4 | 4 | 15 | |
| CC002 | F | 2 | 4 | 3 | 4 | 13 |
| M | 3 | 4 | 3 | 4 | 14 | |
| CC023 | F | 4 | 4 | 3 | 4 | 15 |
| M | 9 | 6 | 3 | 4 | 22 | |
| CC027 | F | 3 | 4 | 3 | 4 | 14 |
| M | 3 | 4 | 3 | 4 | 14 | |
| CC057 | F | 4 | 4 | 3 | 4 | 15 |
| M | 3 | 4 | 3 | 5 | 15 | |
| CC078 | F | 3 | 3 | 3 | 7 | 16 |
| M | 3 | 4 | 3 | 4 | 14 | |
| Total | 43 | 49 | 38 | 52 | 182 | |
| Cytokines and Chemokines Affected by Intracranial Injection throughout Post-Injection Timepoints | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Strain | D.P.I. | IL-1α | IL-1β | IL-2 | IL-3 | IL-4 | IL-5 | IL-6 | IL-9 | IL-10 | IL-12(p40) | IL-12(p70) | IL-13 | IL-17α | Eotaxin | G-CSF | GM-CSF | IFNɣ | KC | MCP-1 | MIP-1α | MIP-1β | RANTES | TNF-α |
| C57BL/6J | 4 | |||||||||||||||||||||||
| 14 | ||||||||||||||||||||||||
| CC002 | 4 | |||||||||||||||||||||||
| 14 | ||||||||||||||||||||||||
| CC023 | 4 | |||||||||||||||||||||||
| 14 | ||||||||||||||||||||||||
| CC027 | 4 | |||||||||||||||||||||||
| 14 | ||||||||||||||||||||||||
| CC057 | 4 | |||||||||||||||||||||||
| 14 | ||||||||||||||||||||||||
| CC078 | 4 | |||||||||||||||||||||||
| 14 | ||||||||||||||||||||||||
| Cytokine and Chemokine Changes throughout 4 d.p.i. and 14 d.p.i. | ||
|---|---|---|
| Strain | Control | Infected |
| C57BL/6J | Eotaxin | IL-5 Eotaxin GM-CSF |
| CC002 | IL-6 IL-12(p40) KC MIP-1β RANTES | IL-3 IL-6 IL-12(p40) IL-17α G-CSF RANTES |
| CC023 | IL-12(p70) | IL-5 IL-6 IL-13 KC MIP-1α |
| CC027 | IL-4 IL-17α GM-CSF IFN-γ | IL-3 IL-10 IL-17α G-CSF GM-CSF KC RANTES |
| CC057 | IL-6 IL-12(p40) | IL-6 IL-12(p40) MIP-1β |
| CC078 | GM-CSF KC MCP-1 MIP-1α RANTES | IL-12(p40) |
| Gender-Specific Differences among Cytokine and Chemokine Levels across the Acute Phase of Infection | ||||||
|---|---|---|---|---|---|---|
| Strain | Female | Male | p-Value | |||
| 4 d.p.i. | 14 d.p.i. | 4 d.p.i. | 14 d.p.i. | |||
| C57BL/6J | Eotaxin | 0.05 | * | |||
| CC002 | Eotaxin s | 0.01 | ** | |||
| CC023 | Eotaxin | Eotaxin s | Eotaxin s | 0.001 | *** | |
| CC027 | IL-12(p40) | Eotaxin | Eotaxin | G-CSF s | 0.0001 | **** |
| G-CSF s | ||||||
| CC057 | IL-12(p40) s Eotaxin s | |||||
| CC078 | RANTES | |||||
| Stepwise Regression Analysis Output per Phenotype | |||||
|---|---|---|---|---|---|
| Phenotype | Variables | Estimate | Std. Error | p-Value | Significance |
| Reflex | TNF-α | −0.078 | 0.015 | 1.28 × 10−6 | *** |
| IL-1β | 0.035 | 0.01 | 0.0015 | ** | |
| Limb Paralysis | IL-9 | −0.060 | 0.027 | 0.0305 | * |
| TMEV infection | −0.0120 | 0.028 | 1.47 × 10−4 | *** | |
| Limb Weakness | TNF-α | −0.057 | 0.014 | 7.31 × 10−5 | *** |
| IL-1β | 0.031 | 0.010 | 0.0021 | ** | |
| MIP-1β | −0.020 | 0.008 | 0.0114 | * | |
| TMEV infection | −0.073 | 0.026 | 0.0053 | ** | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez Gómez, A.A.; Karmakar, M.; Carroll, R.J.; Lawley, K.S.; Amstalden, K.; Young, C.R.; Threadgill, D.W.; Welsh, C.J.; Brinkmeyer-Langford, C. Serum Cytokines Predict Neurological Damage in Genetically Diverse Mouse Models. Cells 2022, 11, 2044. https://doi.org/10.3390/cells11132044
Pérez Gómez AA, Karmakar M, Carroll RJ, Lawley KS, Amstalden K, Young CR, Threadgill DW, Welsh CJ, Brinkmeyer-Langford C. Serum Cytokines Predict Neurological Damage in Genetically Diverse Mouse Models. Cells. 2022; 11(13):2044. https://doi.org/10.3390/cells11132044
Chicago/Turabian StylePérez Gómez, Aracely A., Moumita Karmakar, Raymond J. Carroll, Koedi S. Lawley, Katia Amstalden, Colin R. Young, David W. Threadgill, C. Jane Welsh, and Candice Brinkmeyer-Langford. 2022. "Serum Cytokines Predict Neurological Damage in Genetically Diverse Mouse Models" Cells 11, no. 13: 2044. https://doi.org/10.3390/cells11132044
APA StylePérez Gómez, A. A., Karmakar, M., Carroll, R. J., Lawley, K. S., Amstalden, K., Young, C. R., Threadgill, D. W., Welsh, C. J., & Brinkmeyer-Langford, C. (2022). Serum Cytokines Predict Neurological Damage in Genetically Diverse Mouse Models. Cells, 11(13), 2044. https://doi.org/10.3390/cells11132044