
Mucosal Plasma Cell Activation and Proximity to Nerve Fibres Are Associated with Glycocalyx Reduction in Diarrhoea-Predominant Irritable Bowel Syndrome: Jejunal Barrier Alterations Underlying Clinical Manifestations

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants and Clinical Assessment
2.2. Experimental Design and Procedures
2.3. Collection of Biological Samples
2.4. Analytical Procedures
2.4.1. Gene Expression Analysis by RNAseq and Q-RT-PCR
2.4.2. Histological Assessment
2.4.3. Ultrastructural Analysis
- -
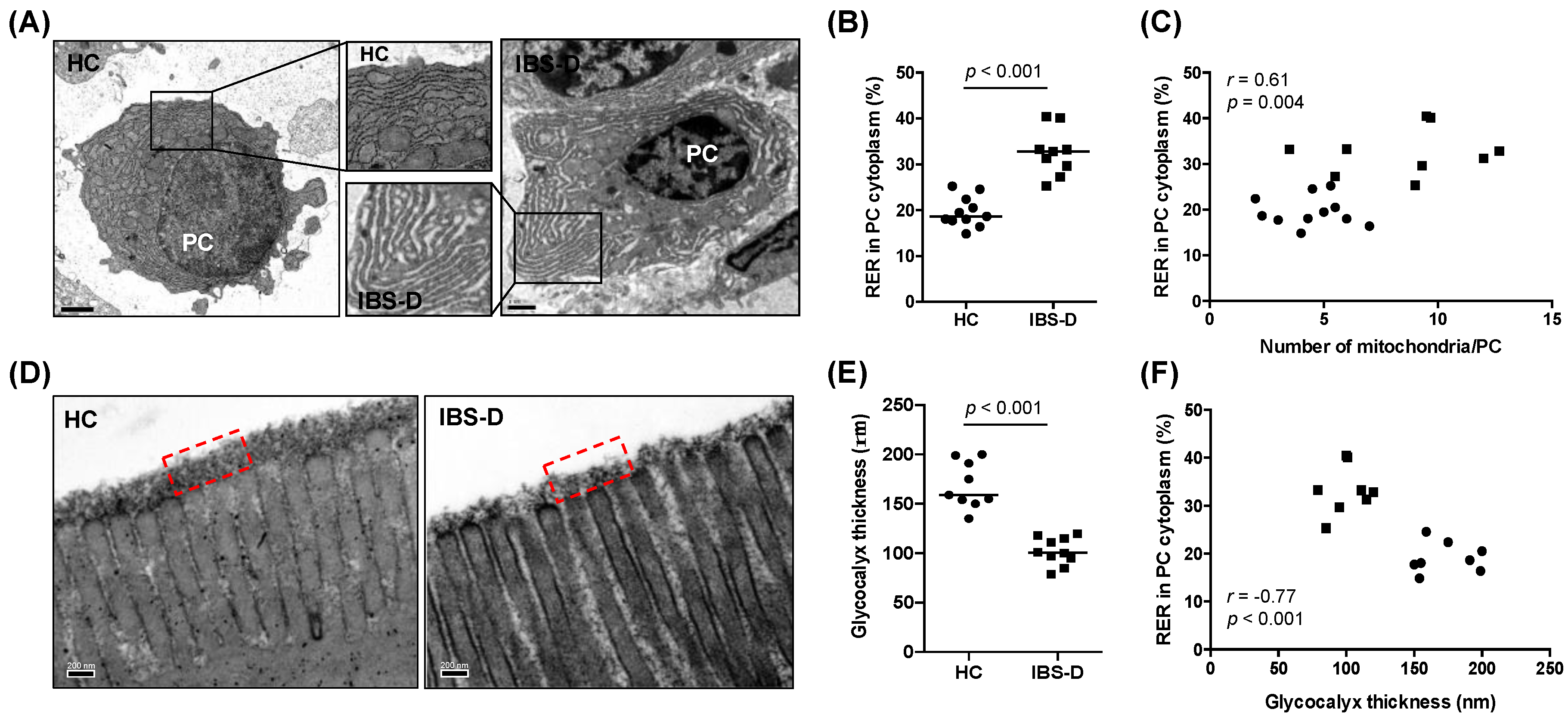
- Plasma cell activation: The rough endoplasmic reticulum (RER) and the mitochondria were quantitatively assessed as previously described [35] with modifications in 3–4 plasma cells per subject. Results are expressed as RER area (total [µm2] and %RER in the cytoplasm) and average number and area (µm2) of mitochondria per plasma cell. A detailed description of the process and validation of the method can be found in the supplementary material (Table S3).
- -
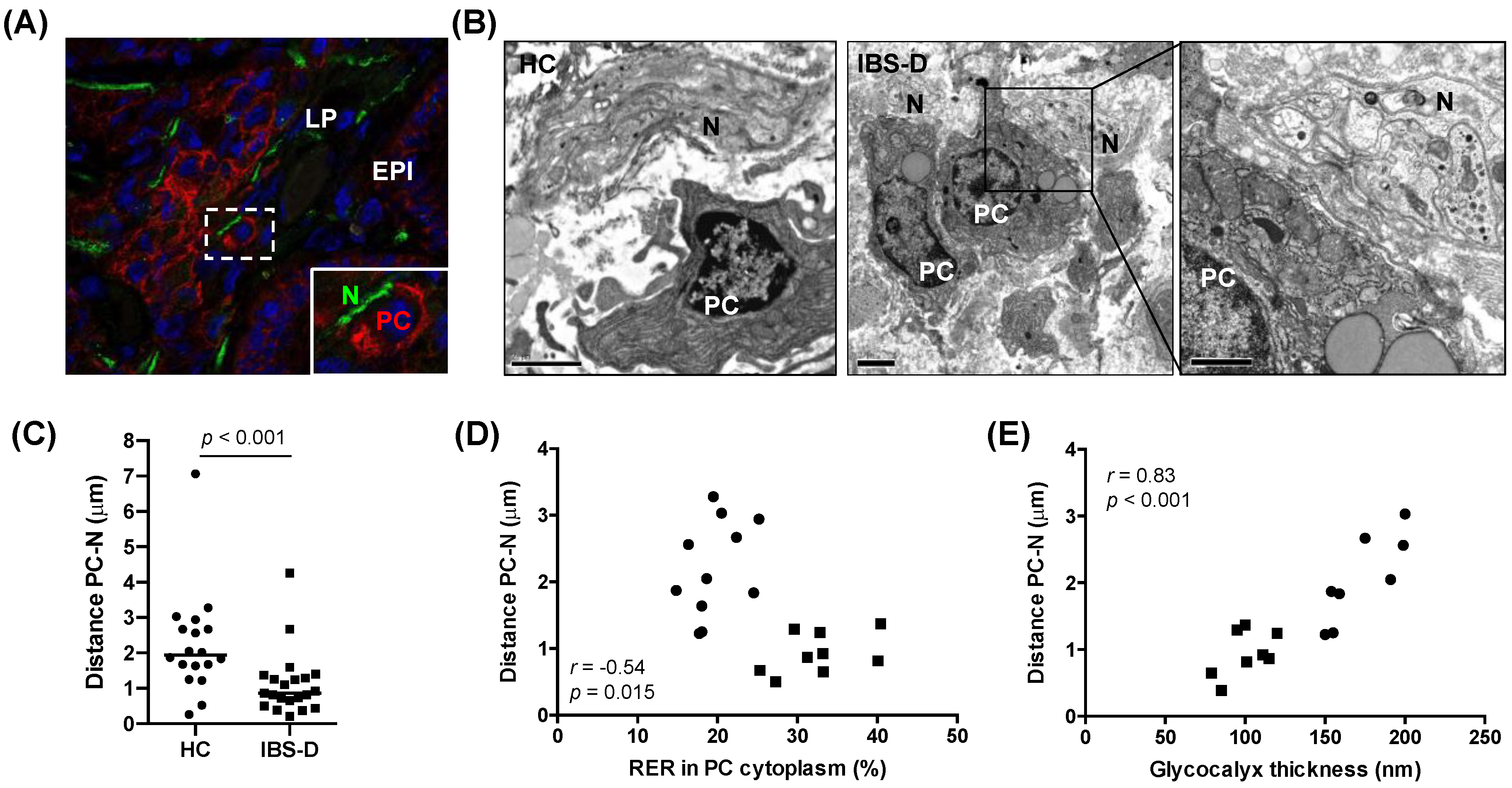
- Plasma cell proximity to nerve fibres: The distance between plasma cells and nerves (membrane to membrane) was measured (4–6 measurements per plasma cell) in 7–10 plasma cells per subject. Results are expressed as the average distance in µm.
- -
- Glycocalyx thickness: The thickness of the glycocalyx overlying the microvilli of the enterocytes was measured in ten non-overlapping fields per subject. Representative measurements (3–7 per field) that included the highest and lowest thickness were taken, and the results are expressed as the average thickness in nm.
2.4.4. Immunoglobulin Quantification
2.5. Statistical Analysis
3. Results
3.1. Study Population
Jejunal Histology and Immune Cell Counts
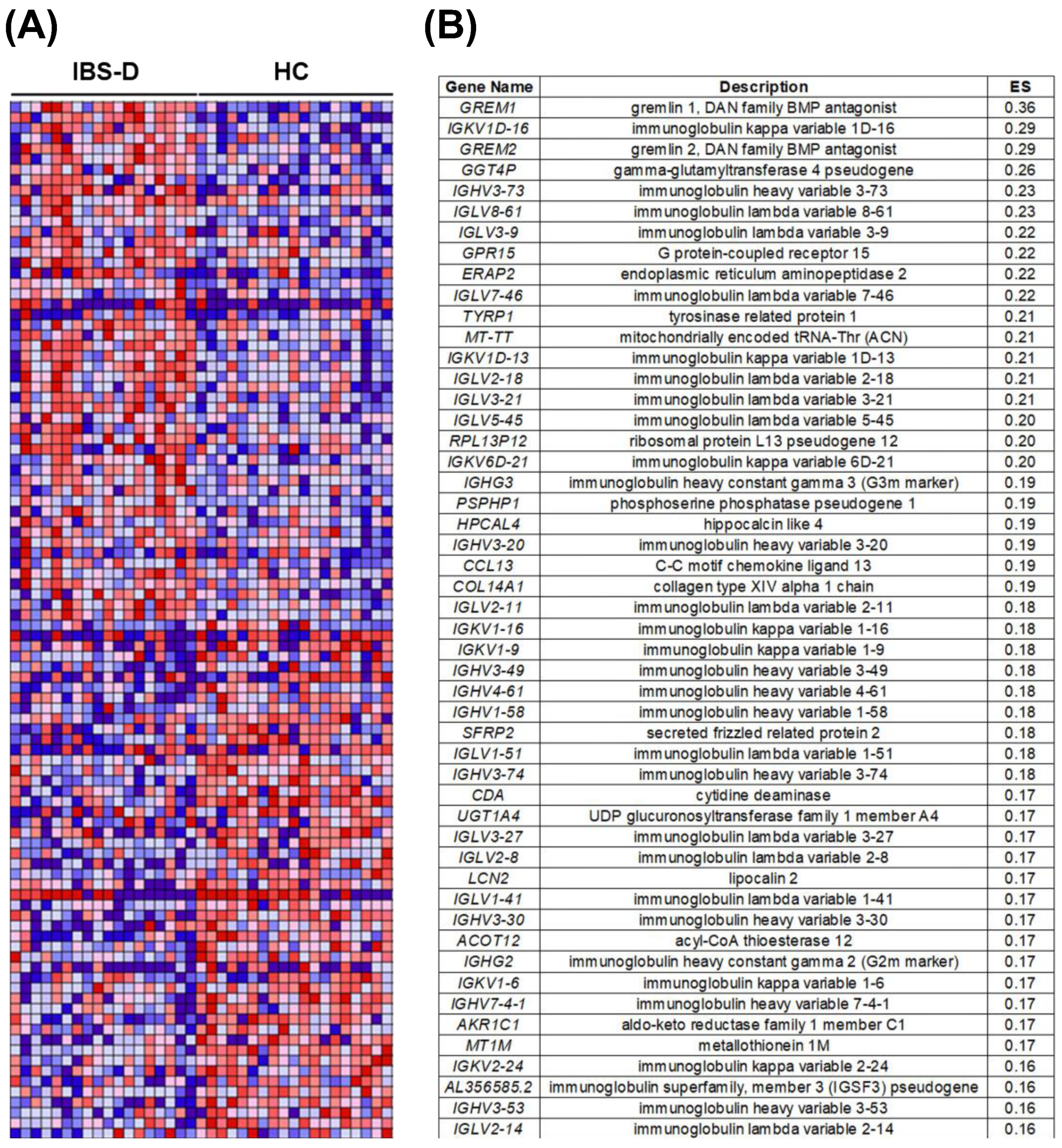
3.2. The Jejunal Mucosa Shows Higher Representation of Humoral Activity Pathways in IBS-D Compared to Health
3.3. Ultrastructural Assessment Shows Differential Plasma Cell Activation in the Jejunal Mucosa in IBS-D
3.4. Reduced Glycocalyx in IBS-D in Association with Plasma Cell Activation
3.5. Increased Mucosal Plasma Cell Proximity to Nerve Fibres in IBS-D, in Association with Plasma cell Activation and Glycocalyx Reduction
3.6. Higher Concentration of IgG in the Intestinal Lumen in IBS-D
3.7. Associations between Clinical, Histological, and Biological Variables
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Drossman, D.A. Functional Gastrointestinal Disorders: History, Pathophysiology, Clinical Features and Rome IV. Gastroenterology 2016, 150, 1262–1279.e2. [Google Scholar] [CrossRef] [PubMed]
- Barbara, G.; Cremon, C.; De Giorgio, R.; Dothel, G.; Zecchi, L.; Bellacosa, L.; Carini, G.; Stanghellini, V.; Corinaldesi, R. Mechanisms underlying visceral hypersensitivity in irritable bowel syndrome. Curr. Gastroenterol. Rep. 2011, 13, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Enck, P.; Aziz, Q.; Barbara, G.; Farmer, A.D.; Fukudo, S.; Mayer, E.A.; Niesler, B.; Quigley, E.M.; Rajilic-Stojanovic, M.; Schemann, M.; et al. Irritable bowel syndrome. Nat. Rev. Dis. Primers 2016, 2, 16014. [Google Scholar] [CrossRef] [PubMed]
- Ford, A.C.; Sperber, A.D.; Corsetti, M.; Camilleri, M. Irritable bowel syndrome. Lancet 2020, 396, 1675–1688. [Google Scholar] [CrossRef]
- Sibelli, A.; Chalder, T.; Everitt, H.; Workman, P.; Windgassen, S.; Moss-Morris, R. A systematic review with meta-analysis of the role of anxiety and depression in irritable bowel syndrome onset. Psychol. Med. 2016, 46, 3065–3080. [Google Scholar] [CrossRef]
- Bertiaux-Vandaele, N.; Youmba, S.B.; Belmonte, L.; Lecleire, S.; Antonietti, M.; Gourcerol, G.; Leroi, A.M.; Dechelotte, P.; Menard, J.F.; Ducrotte, P.; et al. The expression and the cellular distribution of the tight junction proteins are altered in irritable bowel syndrome patients with differences according to the disease subtype. Am. J. Gastroenterol. 2011, 106, 2165–2173. [Google Scholar] [CrossRef]
- Piche, T.; Barbara, G.; Aubert, P.; Bruley des Varannes, S.; Dainese, R.; Nano, J.L.; Cremon, C.; Stanghellini, V.; De Giorgio, R.; Galmiche, J.P.; et al. Impaired intestinal barrier integrity in the colon of patients with irritable bowel syndrome: Involvement of soluble mediators. Gut 2009, 58, 196–201. [Google Scholar] [CrossRef]
- Martinez, C.; Lobo, B.; Pigrau, M.; Ramos, L.; Gonzalez-Castro, A.M.; Alonso, C.; Guilarte, M.; Guila, M.; de Torres, I.; Azpiroz, F.; et al. Diarrhoea-predominant irritable bowel syndrome: An organic disorder with structural abnormalities in the jejunal epithelial barrier. Gut 2013, 62, 1160–1168. [Google Scholar] [CrossRef]
- Zhou, Q.; Zhang, B.; Verne, G.N. Intestinal membrane permeability and hypersensitivity in the irritable bowel syndrome. Pain 2009, 146, 41–46. [Google Scholar] [CrossRef]
- Farre, R.; Vicario, M. Abnormal Barrier Function in Gastrointestinal Disorders. Handb. Exp. Pharmacol. 2017, 239, 193–217. [Google Scholar] [CrossRef]
- Bashashati, M.; Moossavi, S.; Cremon, C.; Barbaro, M.R.; Moraveji, S.; Talmon, G.; Rezaei, N.; Hughes, P.A.; Bian, Z.X.; Choi, C.H.; et al. Colonic immune cells in irritable bowel syndrome: A systematic review and meta-analysis. Neurogastroenterol. Motil. 2018, 30, e13192. [Google Scholar] [CrossRef]
- Robles, A.; Perez Ingles, D.; Myneedu, K.; Deoker, A.; Sarosiek, I.; Zuckerman, M.J.; Schmulson, M.J.; Bashashati, M. Mast cells are increased in the small intestinal mucosa of patients with irritable bowel syndrome: A systematic review and meta-analysis. Neurogastroenterol. Motil. 2019, 31, e13718. [Google Scholar] [CrossRef]
- Vicario, M.; Gonzalez-Castro, A.M.; Martinez, C.; Lobo, B.; Pigrau, M.; Guilarte, M.; de Torres, I.; Mosquera, J.L.; Fortea, M.; Sevillano-Aguilera, C.; et al. Increased humoral immunity in the jejunum of diarrhoea-predominant irritable bowel syndrome associated with clinical manifestations. Gut 2015, 64, 1379–1388. [Google Scholar] [CrossRef]
- Liu, Y.; Yuan, X.; Li, L.; Lin, L.; Zuo, X.; Cong, Y.; Li, Y. Increased Ileal Immunoglobulin A Production and Immunoglobulin A-Coated Bacteria in Diarrhea-Predominant Irritable Bowel Syndrome. Clin. Transl. Gastroenterol. 2020, 11, e00146. [Google Scholar] [CrossRef]
- Dlugosz, A.; Nowak, P.; D’Amato, M.; Mohammadian Kermani, G.; Nystrom, J.; Abdurahman, S.; Lindberg, G. Increased serum levels of lipopolysaccharide and antiflagellin antibodies in patients with diarrhea-predominant irritable bowel syndrome. Neurogastroenterol. Motil. 2015, 27, 1747–1754. [Google Scholar] [CrossRef]
- Martinez, C.; Vicario, M.; Ramos, L.; Lobo, B.; Mosquera, J.L.; Alonso, C.; Sanchez, A.; Guilarte, M.; Antolin, M.; de Torres, I.; et al. The jejunum of diarrhea-predominant irritable bowel syndrome shows molecular alterations in the tight junction signaling pathway that are associated with mucosal pathobiology and clinical manifestations. Am. J. Gastroenterol. 2012, 107, 736–746. [Google Scholar] [CrossRef]
- Rodino-Janeiro, B.K.; Martinez, C.; Fortea, M.; Lobo, B.; Pigrau, M.; Nieto, A.; Gonzalez-Castro, A.M.; Salvo-Romero, E.; Guagnozzi, D.; Pardo-Camacho, C.; et al. Decreased TESK1-mediated cofilin 1 phosphorylation in the jejunum of IBS-D patients may explain increased female predisposition to epithelial dysfunction. Sci. Rep. 2018, 8, 2255. [Google Scholar] [CrossRef]
- Hanning, N.; Edwinson, A.L.; Ceuleers, H.; Peters, S.A.; De Man, J.G.; Hassett, L.C.; De Winter, B.Y.; Grover, M. Intestinal barrier dysfunction in irritable bowel syndrome: A systematic review. Ther. Adv. Gastroenterol. 2021, 14, 1–31. [Google Scholar] [CrossRef]
- Pelaseyed, T.; Bergstrom, J.H.; Gustafsson, J.K.; Ermund, A.; Birchenough, G.M.; Schutte, A.; van der Post, S.; Svensson, F.; Rodriguez-Pineiro, A.M.; Nystrom, E.E.; et al. The mucus and mucins of the goblet cells and enterocytes provide the first defense line of the gastrointestinal tract and interact with the immune system. Immunol. Rev. 2014, 260, 8–20. [Google Scholar] [CrossRef]
- Grondin, J.A.; Kwon, Y.H.; Far, P.M.; Haq, S.; Khan, W.I. Mucins in Intestinal Mucosal Defense and Inflammation: Learning From Clinical and Experimental Studies. Front. Immunol. 2020, 11, 2054. [Google Scholar] [CrossRef]
- Basivireddy, J.; Jacob, M.; Ramamoorthy, P.; Balasubramanian, K.A. Alterations in the intestinal glycocalyx and bacterial flora in response to oral indomethacin. Int. J. Biochem. Cell Biol. 2005, 37, 2321–2332. [Google Scholar] [CrossRef] [PubMed]
- Layunta, E.; Javerfelt, S.; Dolan, B.; Arike, L.; Pelaseyed, T. IL-22 promotes the formation of a MUC17 glycocalyx barrier in the postnatal small intestine during weaning. Cell Rep. 2021, 34, 108757. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Yin, Y.; Qin, M.; Li, K.; Liu, F.; Zhou, X.; Song, X.; Li, B. Vagus Nerve Stimulation Protects Enterocyte Glycocalyx After Hemorrhagic Shock Via the Cholinergic Anti-Inflammatory Pathway. Shock 2021, 56, 832–839. [Google Scholar] [CrossRef] [PubMed]
- Drossman, D.A. The functional gastrointestinal disorders and the Rome III process. Gastroenterology 2006, 130, 1377–1390. [Google Scholar] [CrossRef]
- Francis, C.Y.; Morris, J.; Whorwell, P.J. The irritable bowel severity scoring system: A simple method of monitoring irritable bowel syndrome and its progress. Aliment. Pharmacol. Ther. 1997, 11, 395–402. [Google Scholar] [CrossRef]
- Lewis, S.J.; Heaton, K.W. Stool form scale as a useful guide to intestinal transit time. Scand. J. Gastroenterol. 1997, 32, 920–924. [Google Scholar] [CrossRef]
- Beck, A.T.; Ward, C.H.; Mendelson, M.; Mock, J.; Erbaugh, J. An inventory for measuring depression. Arch. Gen. Psychiatry 1961, 4, 561–571. [Google Scholar] [CrossRef]
- Holmes, T.H.; Rahe, R.H. The Social Readjustment Rating Scale. J. Psychosom. Res. 1967, 11, 213–218. [Google Scholar] [CrossRef]
- Cohen, S.; Kamarck, T.; Mermelstein, R. A global measure of perceived stress. J. Health Soc. Behav. 1983, 24, 385–396. [Google Scholar] [CrossRef]
- Guilarte, M.; Santos, J.; de Torres, I.; Alonso, C.; Vicario, M.; Ramos, L.; Martinez, C.; Casellas, F.; Saperas, E.; Malagelada, J.R. Diarrhoea-predominant IBS patients show mast cell activation and hyperplasia in the jejunum. Gut 2007, 56, 203–209. [Google Scholar] [CrossRef]
- Martinez, C.; Rodino-Janeiro, B.K.; Lobo, B.; Stanifer, M.L.; Klaus, B.; Granzow, M.; Gonzalez-Castro, A.M.; Salvo-Romero, E.; Alonso-Cotoner, C.; Pigrau, M.; et al. miR-16 and miR-125b are involved in barrier function dysregulation through the modulation of claudin-2 and cingulin expression in the jejunum in IBS with diarrhoea. Gut 2017, 66, 1537–1538. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef]
- Ferguson, A.; Murray, D. Quantitation of intraepithelial lymphocytes in human jejunum. Gut 1971, 12, 988–994. [Google Scholar] [CrossRef]
- Kirk, S.J.; Cliff, J.M.; Thomas, J.A.; Ward, T.H. Biogenesis of secretory organelles during B cell differentiation. J. Leukoc. Biol. 2010, 87, 245–255. [Google Scholar] [CrossRef]
- Vivinus-Nebot, M.; Dainese, R.; Anty, R.; Saint-Paul, M.C.; Nano, J.L.; Gonthier, N.; Marjoux, S.; Frin-Mathy, G.; Bernard, G.; Hebuterne, X.; et al. Combination of allergic factors can worsen diarrheic irritable bowel syndrome: Role of barrier defects and mast cells. Am. J. Gastroenterol. 2012, 107, 75–81. [Google Scholar] [CrossRef]
- Sun, W.W.; Krystofiak, E.S.; Leo-Macias, A.; Cui, R.; Sesso, A.; Weigert, R.; Ebrahim, S.; Kachar, B. Nanoarchitecture and dynamics of the mouse enteric glycocalyx examined by freeze-etching electron tomography and intravital microscopy. Commun. Biol. 2020, 3, 5. [Google Scholar] [CrossRef]
- Fang, J.; Wang, H.; Zhou, Y.; Zhang, H.; Zhou, H.; Zhang, X. Slimy partners: The mucus barrier and gut microbiome in ulcerative colitis. Exp. Mol. Med. 2021, 53, 772–787. [Google Scholar] [CrossRef]
- Qin, X. Damage of the Mucus Layer: The Possible Shared Critical Common Cause for Both Inflammatory Bowel Disease (IBD) and Irritable Bowel Syndrome (IBS). Inflamm. Bowel Dis. 2017, 23, E11–E12. [Google Scholar] [CrossRef]
- Aerssens, J.; Camilleri, M.; Talloen, W.; Thielemans, L.; Gohlmann, H.W.; Van Den Wyngaert, I.; Thielemans, T.; De Hoogt, R.; Andrews, C.N.; Bharucha, A.E.; et al. Alterations in mucosal immunity identified in the colon of patients with irritable bowel syndrome. Clin. Gastroenterol. Hepatol. 2008, 6, 194–205. [Google Scholar] [CrossRef]
- Jabbar, K.S.; Dolan, B.; Eklund, L.; Wising, C.; Ermund, A.; Johansson, A.; Tornblom, H.; Simren, M.; Hansson, G.C. Association between Brachyspira and irritable bowel syndrome with diarrhoea. Gut 2021, 70, 1117–1129. [Google Scholar] [CrossRef]
- Wang, S.H.; Dong, L.; Luo, J.Y.; Gong, J.; Li, L.; Lu, X.L.; Han, S.P. Decreased expression of serotonin in the jejunum and increased numbers of mast cells in the terminal ileum in patients with irritable bowel syndrome. World J. Gastroenterol. 2007, 13, 6041–6047. [Google Scholar] [CrossRef]
- Ford, A.C.; Lacy, B.E.; Harris, L.A.; Quigley, E.M.M.; Moayyedi, P. Effect of Antidepressants and Psychological Therapies in Irritable Bowel Syndrome: An Updated Systematic Review and Meta-Analysis. Am. J. Gastroenterol. 2019, 114, 21–39. [Google Scholar] [CrossRef]
- Hansen, M.B.; Witte, A.B. The role of serotonin in intestinal luminal sensing and secretion. Acta Physiol. 2008, 193, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Arabyan, N.; Park, D.; Foutouhi, S.; Weis, A.M.; Huang, B.C.; Williams, C.C.; Desai, P.; Shah, J.; Jeannotte, R.; Kong, N.; et al. Salmonella Degrades the Host Glycocalyx Leading to Altered Infection and Glycan Remodeling. Sci. Rep. 2016, 6, 29525. [Google Scholar] [CrossRef] [PubMed]
- Bednarska, O.; Walter, S.A.; Casado-Bedmar, M.; Strom, M.; Salvo-Romero, E.; Vicario, M.; Mayer, E.A.; Keita, A.V. Vasoactive Intestinal Polypeptide and Mast Cells Regulate Increased Passage of Colonic Bacteria in Patients With Irritable Bowel Syndrome. Gastroenterology 2017, 153, 948–960 e943. [Google Scholar] [CrossRef] [PubMed]
- Martino, C.; Kellman, B.P.; Sandoval, D.R.; Clausen, T.M.; Marotz, C.A.; Song, S.J.; Wandro, S.; Zaramela, L.S.; Salido Benitez, R.A.; Zhu, Q.; et al. Bacterial modification of the host glycosaminoglycan heparan sulfate modulates SARS-CoV-2 infectivity. bioRxiv 2020. [Google Scholar] [CrossRef]
- Wang, L.; Alammar, N.; Singh, R.; Nanavati, J.; Song, Y.; Chaudhary, R.; Mullin, G.E. Gut Microbial Dysbiosis in the Irritable Bowel Syndrome: A Systematic Review and Meta-Analysis of Case-Control Studies. J. Acad. Nutr. Diet. 2020, 120, 565–586. [Google Scholar] [CrossRef]
- Saffouri, G.B.; Shields-Cutler, R.R.; Chen, J.; Yang, Y.; Lekatz, H.R.; Hale, V.L.; Cho, J.M.; Battaglioli, E.J.; Bhattarai, Y.; Thompson, K.J.; et al. Small intestinal microbial dysbiosis underlies symptoms associated with functional gastrointestinal disorders. Nat. Commun. 2019, 10, 2012. [Google Scholar] [CrossRef]
- Qin, H.Y.; Cheng, C.W.; Tang, X.D.; Bian, Z.X. Impact of psychological stress on irritable bowel syndrome. World J. Gastroenterol. 2014, 20, 14126–14131. [Google Scholar] [CrossRef]
- Rengarajan, S.; Knoop, K.A.; Rengarajan, A.; Chai, J.N.; Grajales-Reyes, J.G.; Samineni, V.K.; Russler-Germain, E.V.; Ranganathan, P.; Fasano, A.; Sayuk, G.S.; et al. A Potential Role for Stress-Induced Microbial Alterations in IgA-Associated Irritable Bowel Syndrome with Diarrhea. Cell Rep. Med. 2020, 1, 100124. [Google Scholar] [CrossRef]
- Vicario, M.; Guilarte, M.; Alonso, C.; Yang, P.; Martinez, C.; Ramos, L.; Lobo, B.; Gonzalez, A.; Guila, M.; Pigrau, M.; et al. Chronological assessment of mast cell-mediated gut dysfunction and mucosal inflammation in a rat model of chronic psychosocial stress. Brain Behav. Immun. 2010, 24, 1166–1175. [Google Scholar] [CrossRef]
- Da Silva, S.; Robbe-Masselot, C.; Ait-Belgnaoui, A.; Mancuso, A.; Mercade-Loubiere, M.; Salvador-Cartier, C.; Gillet, M.; Ferrier, L.; Loubiere, P.; Dague, E.; et al. Stress disrupts intestinal mucus barrier in rats via mucin O-glycosylation shift: Prevention by a probiotic treatment. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G420–G429. [Google Scholar] [CrossRef]
- Chassaing, B.; Koren, O.; Goodrich, J.K.; Poole, A.C.; Srinivasan, S.; Ley, R.E.; Gewirtz, A.T. Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syndrome. Nature 2015, 519, 92–96. [Google Scholar] [CrossRef]
- Jakobsson, H.E.; Rodriguez-Pineiro, A.M.; Schutte, A.; Ermund, A.; Boysen, P.; Bemark, M.; Sommer, F.; Backhed, F.; Hansson, G.C.; Johansson, M.E. The composition of the gut microbiota shapes the colon mucus barrier. EMBO Rep. 2015, 16, 164–177. [Google Scholar] [CrossRef]
- Fenton, C.G.; Taman, H.; Florholmen, J.; Sorbye, S.W.; Paulssen, R.H. Transcriptional Signatures That Define Ulcerative Colitis in Remission. Inflamm. Bowel Dis. 2021, 27, 94–105. [Google Scholar] [CrossRef]
- Mockl, L. The Emerging Role of the Mammalian Glycocalyx in Functional Membrane Organization and Immune System Regulation. Front. Cell Dev. Biol. 2020, 8, 253. [Google Scholar] [CrossRef]
- Mole, C.M.; Bene, M.C.; Montagne, P.M.; Seilles, E.; Faure, G.C. Light chains of immunoglobulins in human secretions. Clin. Chim. Acta 1994, 224, 191–197. [Google Scholar] [CrossRef]
- Dlugosz, A.; Tornblom, H.; Mohammadian, G.; Morgan, G.; Veress, B.; Edvinsson, B.; Sandstrom, G.; Lindberg, G. Chlamydia trachomatis antigens in enteroendocrine cells and macrophages of the small bowel in patients with severe irritable bowel syndrome. BMC Gastroenterol. 2010, 10, 19. [Google Scholar] [CrossRef]
- Lobo, B.; Ramos, L.; Martinez, C.; Guilarte, M.; Gonzalez-Castro, A.M.; Alonso-Cotoner, C.; Pigrau, M.; de Torres, I.; Rodino-Janeiro, B.K.; Salvo-Romero, E.; et al. Downregulation of mucosal mast cell activation and immune response in diarrhoea-irritable bowel syndrome by oral disodium cromoglycate: A pilot study. United Eur. Gastroenterol. J. 2017, 5, 887–897. [Google Scholar] [CrossRef]
- Nozu, T.; Miyagishi, S.; Nozu, R.; Takakusaki, K.; Okumura, T. Lipopolysaccharide induces visceral hypersensitivity: Role of interleukin-1, interleukin-6, and peripheral corticotropin-releasing factor in rats. J. Gastroenterol. 2017, 52, 72–80. [Google Scholar] [CrossRef]
- Ferrante, A.; Beard, L.J.; Feldman, R.G. IgG subclass distribution of antibodies to bacterial and viral antigens. Pediatr. Infect. Dis. J. 1990, 9, S16–S24. [Google Scholar] [CrossRef]
- Goldenberg, H.B.; McCool, T.L.; Weiser, J.N. Cross-reactivity of human immunoglobulin G2 recognizing phosphorylcholine and evidence for protection against major bacterial pathogens of the human respiratory tract. J. Infect. Dis. 2004, 190, 1254–1263. [Google Scholar] [CrossRef]
- Kluber, P.; Meurer, S.K.; Lambertz, J.; Schwarz, R.; Zechel-Gran, S.; Braunschweig, T.; Hurka, S.; Domann, E.; Weiskirchen, R. Depletion of Lipocalin 2 (LCN2) in Mice Leads to Dysbiosis and Persistent Colonization with Segmented Filamentous Bacteria. Int. J. Mol. Sci. 2021, 22, 13156. [Google Scholar] [CrossRef]
- Waters, L.R.; Ahsan, F.M.; Wolf, D.M.; Shirihai, O.; Teitell, M.A. Initial B Cell Activation Induces Metabolic Reprogramming and Mitochondrial Remodeling. iScience 2018, 5, 99–109. [Google Scholar] [CrossRef]
- Kimata, H.; Yoshida, A.; Ishioka, C.; Fujimoto, M.; Furusho, K. Vasoactive intestinal peptide enhances immunoglobulin production and growth in human plasma cells via mechanisms that may involve protein kinase C. J. Clin. Endocrinol. Metab. 1996, 81, 3024–3032. [Google Scholar] [CrossRef]
- Scicchitano, R.; Biennenstock, J.; Stanisz, A.M. In vivo immunomodulation by the neuropeptide substance P. Immunology 1988, 63, 733–735. [Google Scholar]
- Xu, X.J.; Zhang, Y.L.; Liu, L.; Pan, L.; Yao, S.K. Increased expression of nerve growth factor correlates with visceral hypersensitivity and impaired gut barrier function in diarrhoea-predominant irritable bowel syndrome: A preliminary explorative study. Aliment. Pharmacol. Ther. 2017, 45, 100–114. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lei, B.; Yuan, Y.; Zhang, L.; Hu, L.; Jin, S.; Kang, B.; Liao, X.; Sun, W.; Xu, F.; et al. Brain control of humoral immune responses amenable to behavioural modulation. Nature 2020, 581, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Lakhan, S.E.; Kirchgessner, A. Neuroinflammation in inflammatory bowel disease. J. Neuroinflamm. 2010, 7, 37. [Google Scholar] [CrossRef] [PubMed]
- Akbar, A.; Yiangou, Y.; Facer, P.; Walters, J.R.; Anand, P.; Ghosh, S. Increased capsaicin receptor TRPV1-expressing sensory fibres in irritable bowel syndrome and their correlation with abdominal pain. Gut 2008, 57, 923–929. [Google Scholar] [CrossRef]
- Sohn, W.; Lee, O.Y.; Lee, S.P.; Lee, K.N.; Jun, D.W.; Lee, H.L.; Yoon, B.C.; Choi, H.S.; Sim, J.; Jang, K.S. Mast cell number, substance P and vasoactive intestinal peptide in irritable bowel syndrome with diarrhea. Scand. J. Gastroenterol. 2014, 49, 43–51. [Google Scholar] [CrossRef]
- Liu, S. Neurotrophic factors in enteric physiology and pathophysiology. Neurogastroenterol. Motil. 2018, 30, e13446. [Google Scholar] [CrossRef]
- Saruta, M.; Takahashi, K.; Suzuki, T.; Torii, A.; Kawakami, M.; Sasano, H. Urocortin 1 in colonic mucosa in patients with ulcerative colitis. J. Clin. Endocrinol. Metab. 2004, 89, 5352–5361. [Google Scholar] [CrossRef]
- Yamato, S.; Kurematsu, A.; Amano, T.; Ariga, H.; Ando, T.; Komaki, G.; Wada, K. Urocortin 1: A putative excitatory neurotransmitter in the enteric nervous system. Neurogastroenterol. Motil. 2020, 32, e13842. [Google Scholar] [CrossRef]
- Brandt, R.; Timm, S.; Gorenflos Lopez, J.L.; Kwame Abledu, J.; Kuebler, W.M.; Hackenberger, C.P.R.; Ochs, M.; Lopez-Rodriguez, E. Metabolic Glycoengineering Enables the Ultrastructural Visualization of Sialic Acids in the Glycocalyx of the Alveolar Epithelial Cell Line hAELVi. Front. Bioeng. Biotechnol. 2020, 8, 614357. [Google Scholar] [CrossRef]
- Alonso-Cotoner, C.; Abril-Gil, M.; Albert-Bayo, M.; Mall, J.G.; Exposito, E.; Gonzalez-Castro, A.M.; Lobo, B.; Santos, J. The Role of Purported Mucoprotectants in Dealing with Irritable Bowel Syndrome, Functional Diarrhea, and Other Chronic Diarrheal Disorders in Adults. Adv. Ther. 2021, 38, 2054–2076. [Google Scholar] [CrossRef]
- Azzini, E.; Maiani, G.; Garaguso, I.; Polito, A.; Foddai, M.S.; Venneria, E.; Durazzo, A.; Intorre, F.; Palomba, L.; Rauseo, M.L.; et al. The Potential Health Benefits of Polyphenol-Rich Extracts from Cichorium intybus L. Studied on Caco-2 Cells Model. Oxidative Med. Cell. Longev. 2016, 2016, 1594616. [Google Scholar] [CrossRef]
- Kong, C.; Elderman, M.; Cheng, L.; de Haan, B.J.; Nauta, A.; de Vos, P. Modulation of Intestinal Epithelial Glycocalyx Development by Human Milk Oligosaccharides and Non-Digestible Carbohydrates. Mol. Nutr. Food Res. 2019, 63, e1900303. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| HC | IBS-D | ||
|---|---|---|---|
| (n = 63) | (n = 71) | p | |
| Sex, F:M | 29:34 | 50:21 | 0.005 |
| Age, years (range) | 25 (18–54) | 36 (18–64) | <0.0001 |
| Intensity of abdominal pain, score | - | 44.5 (2–100) | - |
| Frequency of abdominal pain, number of days | - | 5 (1–10) | - |
| Bowel movements, number/day | 1.5 (0.5–2.5) | 3.0 (0.5–12) | <0.0001 |
| Stool form, Bristol score | 3.5 (2–4.5) | 5.5 (3–7) | <0.0001 |
| Dyspepsia (Yes/No) | - | 26/41 | - |
| IBS-SSS | - | 268.5 ± 91.9 | - |
| Dyspeptic | - | 291.4 ± 95.2 | 0.019 * |
| Non-dyspeptic | - | 236.1 ± 77.4 | |
| Holmes-Rahe scale, score | 88 (0–357) | 111 (0–889) | 0.145 |
| Cohen scale, score | 14 (4–37) | 24 (1–41) | <0.0001 |
| Beck’s Depression Inventory, score | 0 (0–22) | 8 (1–31) | <0.0001 |
| CD117+ (cells/hpf) | 20.1 (2.0–59) | 25.4 (5.6–59) | 0.185 |
| CD3+ (cells/100 enterocytes) | 17 (7.0–40) | 16 (1.6–37) | 0.746 |
| Eosin+ (cells/hpf) | 1.5 (0–5.6) | 1.7 (0–18) | 0.213 |
| Reactome Pathway | NES | p | FDR |
|---|---|---|---|
| PTEN regulation | 1.88 | 0.027 | 0.14 |
| Regulation of apoptosis | 1.73 | 0.010 | 0.14 |
| Interleukin 1 signalling | 1.90 | 0.008 | 0.15 |
| Activation of NF-KB in B cells | 1.91 | 0.004 | 0.15 |
| Cellular responses to stress | 1.75 | 0.006 | 0.15 |
| Protein ubiquitination | 1.78 | 0.033 | 0.15 |
| Downstream signalling events of B cell receptor | 1.97 | 0.002 | 0.16 |
| Dectin 1 mediated non canonical NF-KB signalling | 1.69 | 0.032 | 0.17 |
| TNFR2 non-canonical NF-KB pathway | 1.65 | 0.025 | 0.18 |
| Cell extracellular matrix interactions | 1.60 | 0.073 | 0.19 |
| Interleukin 1 family signalling | 1.59 | 0.047 | 0.19 |
| Signalling by the B cell receptor | 1.58 | 0.042 | 0.19 |
| Antigen processing cross presentation | 1.63 | 0.060 | 0.19 |
| Programmed cell death | 2.04 | 0.0001 | 0.20 |
| Regulation of actin dynamics for phagocytic cup formation | 1.56 | 0.034 | 0.21 |
| Antigen activates B cell receptor leading to generation of second messengers | 1.55 | 0.019 | 0.21 |
| FCERI-mediated NF-KB activation | 1.54 | 0.049 | 0.22 |
| Cross presentation of soluble exogenous antigens endosomes | 1.54 | 0.071 | 0.22 |
| Complement cascade | 1.52 | 0.021 | 0.22 |
| Neutrophil degranulation | 1.49 | 0.058 | 0.24 |
| Class I MHC mediated antigen processing presentation | 1.50 | 0.033 | 0.24 |
| Gap junction assembly | 1.50 | 0.063 | 0.24 |
| Gap junction trafficking and regulation | 1.50 | 0.062 | 0.24 |
| Tight junction interactions | 1.48 | 0.059 | 0.24 |
| Immunologic Signature | NES | p | FDR |
|---|---|---|---|
| Plasma cell activity | |||
| Down-regulated genes in naive B cells vs. plasma cells from bone marrow and blood | 1.86 | 0.01 | 0.20 |
| Down-regulated genes in memory IgM B cells vs. plasma cells from bone marrow and blood | 1.67 | 0.04 | 0.23 |
| Down-regulated genes in B lymphocytes from peritoneal cavity vs. spleen | 1.63 | 0.07 | 0.23 |
| Up-regulated genes in plasma cells vs. memory B cells | 1.75 | 0.01 | 0.23 |
| Germinal centre activity | |||
| Upregulated genes in IgD-peripheral blood B cells vs. dark zone germinal centre B cells | 1.87 | 0.01 | 0.22 |
| Down-regulated genes in naive B cell vs. dark zone germinal centre B cells | 1.83 | 0.01 | 0.23 |
| Down-regulated genes in IgD+ peripheral blood B cells vs. dark zone germinal centre B cells | 1.66 | 0.03 | 0.23 |
| Down-regulated genes in pre-germinal centre B cells vs. dark zone germinal centre B cells | 1.81 | 0.01 | 0.23 |
| Down-regulated genes in naive follicular B cells vs. early germinal centre B cells | 1.63 | 0.06 | 0.24 |
| Upregulated genes in germinal centre B cells vs. naive B cells | 1.61 | 0.04 | 0.24 |
| Predominance of B cells | |||
| Down-regulated genes in B cells vs. plasmacytoid dendritic cells | 1.68 | 0.03 | 0.22 |
| Down-regulated genes in Neutrophils vs. B cells | 1.70 | 0.04 | 0.22 |
| Down-regulated genes in naive B cells vs. day 0 monocytes | 1.61 | 0.07 | 0.24 |
| Down-regulated genes in naive B cells vs. plasma cells | 1.58 | 0.02 | 0.25 |
| Up-regulated genes in B cells vs. monocyte macrophages | 1.60 | 0.03 | 0.25 |
| Down-regulated genes in naive CD4 T cells vs. naive B cells | 1.61 | 0.06 | 0.25 |
| Ig (ng/mg Protein) | HC (n = 17) | IBS-D (n = 19) | p |
|---|---|---|---|
| sIgA | 11.55 × 103 (1.34–71.30) | 26.78 × 103 (4.02–221) | 0.099 |
| IgM | 62.54 (3.44–1074) | 109.1 (4.75–1854) | 0.128 |
| IgG | 3.68 (0.61–35.34) | 11.38 (1.5–45.82) | 0.041 |
| IgG1 | 0.15 (0.003–0.82) | 0.07 (0.002–96) | 0.274 |
| IgG2 | 0.40 ± 0.29 | 0.71 ± 0.53 | 0.039 |
| IgG3 | 0.91 (0.19–1.83) | 1.05 (0.28–3.65) | 0.079 |
| IgG4 | 0.028 ± 0.004 | 0.021 ± 0.004 | 0.184 |
| IgE | 0.090 (0–0.472) | 0.159 (0.012–2.590) | 0.104 |
| Glycocalyx Thickness | %RER in PC Cytoplasm | PC-N Distance | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Variables | rs | p | FDR | rs | p | FDR | rs | p | FDR |
| Age | −0.23 | 0.336 | 0.56 | 0.2 | 0.397 | 0.662 | 0.13 | 0.443 | 0.633 |
| Intensity of abdominal pain | 0.34 | 0.365 | 0.521 | 0.55 | 0.196 | 0.57 | 0.18 | 0.45 | 0.563 |
| Frequency of abdominal pain | 0.69 | 0.047 | 0.94 | 0.52 | 0.196 | 0.49 | 0.22 | 0.358 | 0.716 |
| Number of bowel movements | −0.73 | 0.007 | 0.035 | 0.64 | 0.029 | 0.145 | −0.18 | 0.365 | 0.608 |
| Stool form | −0.66 | 0.007 | 0.023 | 0.57 | 0.024 | 0.24 | −0.27 | 0.123 | 0.41 |
| Dyspepsia | −0.21 | 0.834 | 1 | −0.13 | 0.857 | 0.857 | 0.02 | 0.937 | 0.937 |
| IBS-SSS | −0.03 | 0.938 | 0.938 | 0.24 | 0.582 | 0.831 | 0.1 | 0.678 | 0.753 |
| -Dyspeptic | 0.02 | 0.989 | - | 0.54 | 0.297 | - | −0.04 | 0.91 | - |
| -Non-dyspeptic | - Ŧ | - Ŧ | - | - Ŧ | - Ŧ | - | 0.2 | 0.714 | - |
| Holmes-Rage scale | 0.04 | 0.887 | 0.986 | −0.1 | 0.671 | 0.746 | 0.17 | 0.318 | 0.795 |
| Cohen scale | −0.52 | 0.035 | 0.088 | 0.14 | 0.589 | 0.736 | −0.35 | 0.032 | 0.32 |
| Beck’s Depression Inventory | −0.73 | <0.001 | 0.006 | 0.28 | 0.248 | 0.496 | −0.3 | 0.066 | 0.33 |
| CD117+ cells | −0.59 | 0.008 | 0.024 | 0.68 | 0.001 | 0.003 | −0.21 | 0.209 | 0.209 |
| CD3+ cells | −0.51 | 0.027 | 0.041 | 0.27 | 0.245 | 0.368 | 0.27 | 0.106 | 0.159 |
| Eosin+ cells | −0.37 | 0.125 | 0.125 | 0.27 | 0.253 | 0.253 | −0.33 | 0.047 | 0.141 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pardo-Camacho, C.; Ganda Mall, J.-P.; Martínez, C.; Pigrau, M.; Expósito, E.; Albert-Bayo, M.; Melón-Ardanaz, E.; Nieto, A.; Rodiño-Janeiro, B.; Fortea, M.; et al. Mucosal Plasma Cell Activation and Proximity to Nerve Fibres Are Associated with Glycocalyx Reduction in Diarrhoea-Predominant Irritable Bowel Syndrome: Jejunal Barrier Alterations Underlying Clinical Manifestations. Cells 2022, 11, 2046. https://doi.org/10.3390/cells11132046
Pardo-Camacho C, Ganda Mall J-P, Martínez C, Pigrau M, Expósito E, Albert-Bayo M, Melón-Ardanaz E, Nieto A, Rodiño-Janeiro B, Fortea M, et al. Mucosal Plasma Cell Activation and Proximity to Nerve Fibres Are Associated with Glycocalyx Reduction in Diarrhoea-Predominant Irritable Bowel Syndrome: Jejunal Barrier Alterations Underlying Clinical Manifestations. Cells. 2022; 11(13):2046. https://doi.org/10.3390/cells11132046
Chicago/Turabian StylePardo-Camacho, Cristina, John-Peter Ganda Mall, Cristina Martínez, Marc Pigrau, Elba Expósito, Mercé Albert-Bayo, Elisa Melón-Ardanaz, Adoración Nieto, Bruno Rodiño-Janeiro, Marina Fortea, and et al. 2022. "Mucosal Plasma Cell Activation and Proximity to Nerve Fibres Are Associated with Glycocalyx Reduction in Diarrhoea-Predominant Irritable Bowel Syndrome: Jejunal Barrier Alterations Underlying Clinical Manifestations" Cells 11, no. 13: 2046. https://doi.org/10.3390/cells11132046
APA StylePardo-Camacho, C., Ganda Mall, J.-P., Martínez, C., Pigrau, M., Expósito, E., Albert-Bayo, M., Melón-Ardanaz, E., Nieto, A., Rodiño-Janeiro, B., Fortea, M., Guagnozzi, D., Rodriguez-Urrutia, A., Torres, I. d., Santos-Briones, I., Azpiroz, F., Lobo, B., Alonso-Cotoner, C., Santos, J., González-Castro, A. M., & Vicario, M. (2022). Mucosal Plasma Cell Activation and Proximity to Nerve Fibres Are Associated with Glycocalyx Reduction in Diarrhoea-Predominant Irritable Bowel Syndrome: Jejunal Barrier Alterations Underlying Clinical Manifestations. Cells, 11(13), 2046. https://doi.org/10.3390/cells11132046