
ROS Signaling Mediates Directional Cell Elongation and Somatic Cell Fusion in the Red Alga Griffithsia monilis



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Algal Cultures
2.2. Time-Lapse Observation of Wound Response
2.3. Fluorescent and Histochemical Staining of ROS
2.4. Inhibition Experiment
2.5. RNA Extraction and cDNA Synthesis
2.6. 5′RACE PCR and Structure Analysis
2.7. Quantitative PCR and Statistical Analysis
3. Results
3.1. Wound-Healing Process
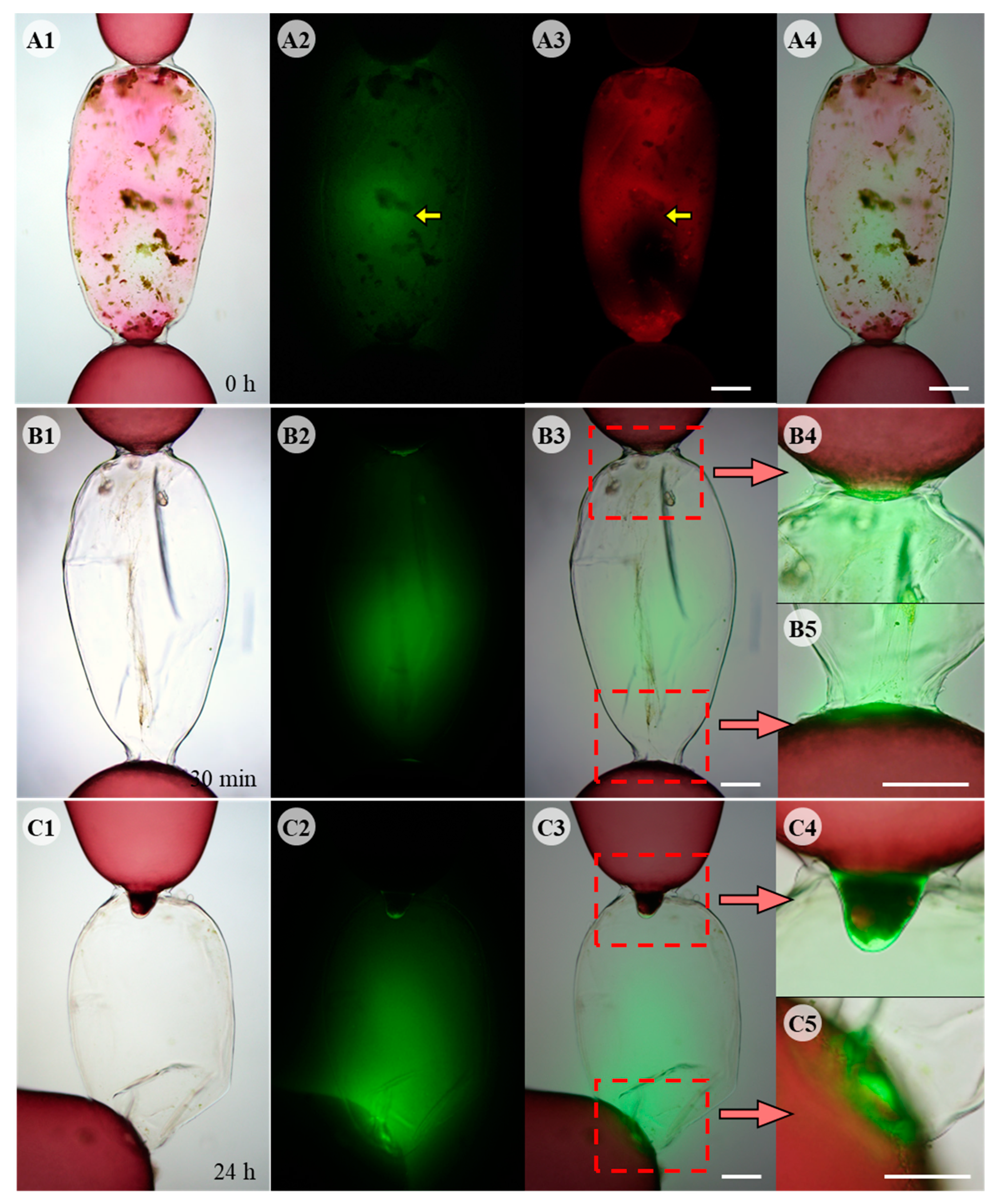
3.2. Visualization of ROS during the Wound-Healing Process
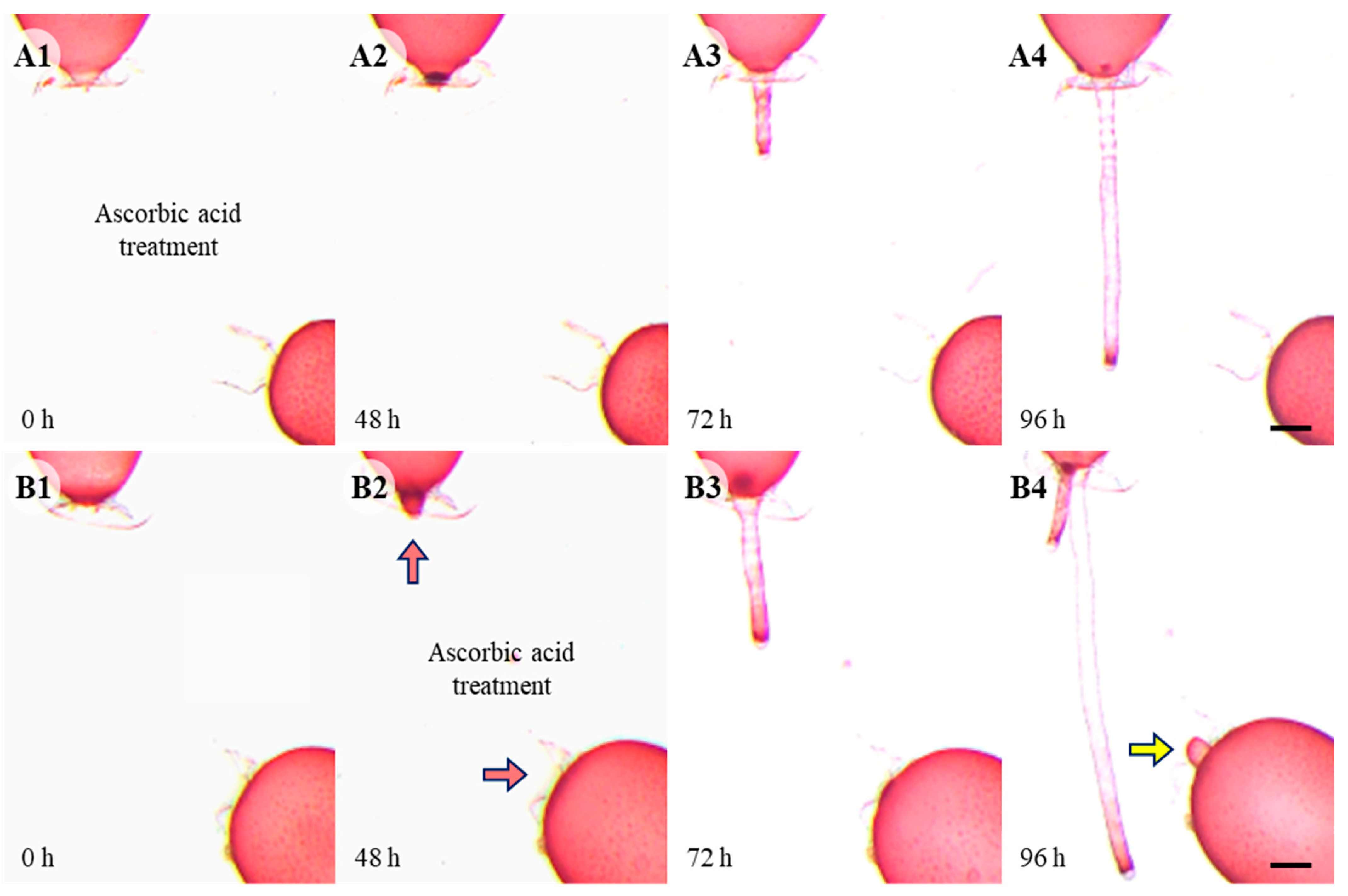
3.3. Effects of ROS and Calcium Inhibitors
3.4. NADPH-Oxidase Homologues and Their Expression in Response to Wounding
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sonnemann, K.J.; Bement, W.M. Wound repair: Toward understanding and integration of single-cell and multicellular wound responses. Annu. Rev. Cell. Dev. Biol. 2011, 27, 237–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, N.; Mittler, R. Reactive oxygen species-dependent wound responses in animals and plants. Free Radic. Biol. Med. 2012, 53, 2269–2276. [Google Scholar] [CrossRef] [PubMed]
- Dunnill, C.; Patton, T.; Brennan, J.; Barrett, J.; Dryden, M.; Cooke, J.; Leaper, D.; Georgopoulos, N.T. Reactive oxygen species (ROS) and wound healing: The functional role of ROS and emerging ROS-modulating technologies for augmentation of the healing process. Int. Wound J. 2017, 14, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lennicke, C.; Rahn, J.; Lichtenfels, R.; Wessjohann, L.A.; Seliger, B. Hydrogen peroxide–production, fate and role in redox signaling of tumor cells. Cell Commun. Signal. 2015, 13, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Anand, A.; Kumari, A.; Thakur, M.; Koul, A. Hydrogen peroxide signaling integrates with phytohormones during the germination of magnetoprimed tomato seeds. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Shim, E.; Lee, J.W.; Park, H.; Zuccarello, G.C.; Kim, G.H. Hydrogen peroxide signalling mediates fertilization and post-fertilization development in the red alga Bostrychia moritziana. J. Exp. Bot. 2022, 73, 727–741. [Google Scholar] [CrossRef]
- Foyer, C.H. Making sense of hydrogen peroxide signals. Nature 2020, 578, 518–519. [Google Scholar] [CrossRef] [Green Version]
- Lambeth, J.D.; Neish, A.S. Nox enzymes and new thinking on reactive oxygen: A double-edged sword revisited. Annu. Rev. Pathol. 2014, 9, 119–145. [Google Scholar] [CrossRef]
- Laurindo, F.R.; Araujo, T.L.; Abrahao, T.B. Nox NADPH oxidases and the endoplasmic reticulum. Antioxid. Redox Signal 2014, 20, 2755–2775. [Google Scholar] [CrossRef] [Green Version]
- Hable, W.E.; Hart, P.E. Signaling mechanisms in the establishment of plant and fucoid algal polarity. Mol. Reprod. Dev. 2010, 77, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Reis, J.; Massari, M.; Marchese, S.; Ceccon, M.; Aalbers, F.S.; Corana, F.; Valente, S.; Mai, A.; Magnani, F.; Mattevi, A. A closer look into NADPH oxidase inhibitors: Validation and insight into their mechanism of action. Redox Biol. 2020, 32, 101466. [Google Scholar] [CrossRef] [PubMed]
- Hervé, C.; Tonon, T.; Collén, J.; Corre, E.; Boyen, C. NADPH oxidases in Eukaryotes: Red algae provide new hints! Curr. Genet. 2006, 49, 190–204. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Zhu, Z.; Yang, R.; Qian, F.; Yan, X.; Chen, H. Characterization of a respiratory burst oxidase homologue from Pyropia haitanensis with unique molecular phylogeny and rapid stress response. J. Appl. Phycol. 2015, 27, 945–955. [Google Scholar] [CrossRef]
- Foreman, J.; Demidchik, V.; Bothwell, J.H.; Mylona, P.; Miedema, H.; Torres, M.A.; Linstead, P.; Costa, S.; Brownlee, C.; Jones, J.D.; et al. Reactive oxygen species produced by NADPH oxidase regulate plant cell growth. Nature 2003, 422, 442–446. [Google Scholar] [CrossRef]
- Brawley, S.H. The fast block against polyspermy in fucoid algae is an electrical block. Dev. Biol. 1991, 144, 94–106. [Google Scholar] [CrossRef]
- Roberts, S.K.; Berger, F.; Brownlee, C. The role of Ca2+ in signal transduction following fertilization in Fucus serratus. J. Exp. Biol. 1993, 184, 197–212. [Google Scholar] [CrossRef]
- Coelho, S.; Brownlee, C.; Bothwell, J.H. A tip-high, Ca2+-interdependent, reactive oxygen species gradient is associated with polarized growth in Fucus serratus zygotes. Planta 2008, 227, 1037–1046. [Google Scholar] [CrossRef]
- Coelho, S.M.; Peters, A.F.; Charrier, B.; Roze, D.; Destombe, C.; Valero, M.; Cock, J.M. Complex life cycles of multicellular eukaryotes: New approaches based on the use of model organisms. Gene 2007, 406, 152–170. [Google Scholar] [CrossRef]
- Brawley, S.H.; Blouin, N.A.; Ficko-Blean, E.; Wheeler, G.L.; Lohr, M.; Goodson, H.V.; Jenkins, J.W.; Blaby-Haas, C.E.; Helliwell, K.E.; Chan, C.X.; et al. Insights into the red algae and eukaryotic evolution from the genome of Porphyra umbilicalis (Bangiophyceae, Rhodophyta). Proc. Natl. Acad. Sci. USA 2017, 114, E6361–E6370. [Google Scholar] [CrossRef] [Green Version]
- Grosberg, R.K.; Strathmann, R.R. The evolution of multicellularity: A minor major transition? Annu. Rev. Ecol. Evol. Syst. 2007, 38, 621–654. [Google Scholar] [CrossRef] [Green Version]
- Sharpe, S.C.; Eme, L.; Brown, M.W.; Roger, A.J. Timing the origins of multicellular eukaryotes through phylogenomics and relaxed molecular clock analyses. In Evolutionary Transitions to Multicellular Life; Ruiz-Trillo, I., Nedelcu, A., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 3–29. [Google Scholar]
- Justice, S.S.; Hunstad, D.A.; Cegelski, L.; Hultgren, S.J. Morphological plasticity as a bacterial survival strategy. Nat. Rev. Microbiol. 2008, 6, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Lyons, N.A.; Kolter, R. On the evolution of bacterial multicellularity. Curr. Opin. Microbiol. 2015, 24, 21–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, H.S.; Zuccarello, G.C.; Bhattacharya, D. Evolutionary history and taxonomy of red algae. In Red Algae in the Genomic Age; Seckbach, J., Chapman, D.J., Weber, A., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 25–42. [Google Scholar]
- Waaland, S.D.; Cleland, R.E. Cell repair through cell fusion in the red alga Griffithsia pacifica. Protoplasma 1974, 79, 185–196. [Google Scholar] [CrossRef]
- Kim, G.H.; Fritz, L. Ultrastructure and cytochemistry of early spermatangial development in Antithamnion nipponicum (Ceramiaceae, Rhodophyta). J. Phycol. 1993, 29, 797–805. [Google Scholar] [CrossRef]
- Kim, H.S.; Kim, G.H.; Lee, I.K. Wound-healing in several filamentous red algae, Ceramiales. Korean J. Phycol. 1988, 3, 15–27. [Google Scholar]
- Kim, G.H.; Lee, I.K.; Fritz, L. The wound-healing responses of Antithamnion nipponicum and Griffithsia pacifica (Ceramiales, Rhodophyta) monitored by lectins. Phycol. Res. 1995, 43, 161–166. [Google Scholar] [CrossRef]
- Lee, Y.K.; Hwang, M.S.; Lee, I.K. Sexual differentiation of Griffithsia monilis (Ceramiales, Rhodophyta) in hybrids between female and male thalli. Bot. Mar. 2001, 44, 547–557. [Google Scholar] [CrossRef]
- Kim, G.H.; Nagasato, C.; Kwak, M.; Lee, J.W.; Hong, C.Y.; Klochkova, T.A.; Motomura, T. Intercellular transport across pit-connections in the filamentous red alga Griffithsia monilis. Algae 2022, 37, 75–84. [Google Scholar] [CrossRef]
- Waaland, S.D.; Watson, B.A. Isolation of a cell-fusion hormone from Griffithsia pacifica Kylin, a red alga. Planta 1980, 149, 493–497. [Google Scholar] [CrossRef]
- Watson, B.A.; Waaland, S.D. Partial purification and characterization of a glycoprotein cell fusion hormone from Griffithsia pacifica, a red alga. Plant Physiol. 1983, 71, 327–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, B.A.; Waaland, S.D. Further biochemical characterization of a cell fusion hormone from the red alga, Griffithsia pacifica. Plant Cell Physiol. 1986, 27, 1043–1050. [Google Scholar]
- Kim, G.H.; Klochkova, T.A.; Yoon, K.S.; Song, Y.S.; Lee, K.P. Purification and characterization of a lectin, bryohealin, involved in the protoplast formation of a marine green alga bryopsis plumosa (chlorophyta). J. Phycol. 2005, 42, 86–95. [Google Scholar] [CrossRef]
- Pickett-Heaps, J.D.; West, J. Time-lapse video observations on sexual plasmogamy in the red alga Bostrychia. Europ. J. Phycol. 1998, 33, 43–56. [Google Scholar] [CrossRef]
- Daudi, A.; O’Brien, J.A. Detection of hydrogen peroxide by DAB staining in Arabidopsis leaves. Bio Protoc. 2012, 2, e263. [Google Scholar] [CrossRef] [Green Version]
- Kumar, D.; Yusuf, M.A.; Singh, P.; Sardar, M.; Sarin, N.B. Histochemical detection of superoxide and H2O2 accumulation in Brassica juncea seedlings. Bio. Protoc. 2014, 4, e1108. [Google Scholar] [CrossRef]
- Taylor, A.R.; Manison, N.F.; Fernandez, C.; Wood, J.; Brownlee, C. Spatial organization of calcium signaling involved in cell volume control in the Fucus rhizoid. Plant Cell. 1996, 8, 2015–2031. [Google Scholar] [CrossRef]
- Länge, S.; Wissmann, J.D.; Plattner, H. Caffeine inhibits Ca2+ uptake by subplasmalemmal calcium stores (‘alveolar sacs’) isolated from Paramecium cells. Biochim. Biophys. Acta (BBA)-Biomembr. 1996, 1278, 191–196. [Google Scholar] [CrossRef] [Green Version]
- Yasuhara, H. Caffeine inhibits callose deposition in the cell plate and the depolymerization of microtubules in the central region of the phragmoplast. Plant Cell Physiol. 2005, 46, 1083–1092. [Google Scholar] [CrossRef] [Green Version]
- Yeung, P.K.K.; Lam, C.M.C.; Ma, Z.Y.; Wong, Y.H.; Wong, J.T. Involvement of calcium mobilization from caffeine-sensitive stores in mechanically induced cell cycle arrest in the dinoflagellate Crypthecodinium cohnii. Cell calcium. 2006, 39, 259–274. [Google Scholar] [CrossRef]
- Lee, J.W.; Kim, G.H. Red and far-red regulation of filament movement correlates with the expression of phytochrome and FHY1 genes in Spirogyra varians (Zygnematales, Streptophyta). J. Phycol. 2019, 55, 688–699. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Smirnoff, N.; Arnaud, D. Hydrogen peroxide metabolism and functions in plants. New Phytol. 2019, 221, 1197–1214. [Google Scholar] [CrossRef] [PubMed]
- Cano Sanchez, M.; Lancel, S.; Boulanger, E.; Neviere, R. Targeting oxidative stress and mitochondrial dysfunction in the treatment of impaired wound healing: A systematic review. Antioxidants 2018, 7, 98. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.A.D.; Tognetti, V.B.; Vandepoele, K.; Gollery, M.; Shulaev, V.; Van Breusegem, F. ROS signaling: The new wave? Trends Plant Sci. 2011, 16, 300–309. [Google Scholar] [CrossRef]
- Miller, G.A.D.; Suzuki, N.; Ciftci-Yilmaz, S.U.L.T.A.N.; Mittler, R.O.N. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef]
- Liszkay, A.; van der Zalm, E.; Schopfer, P. Production of reactive oxygen intermediates (O2−, H2O2, and ˙OH) by maize roots and their role in wall loosening and elongation growth. Plant Physiol. 2004, 136, 3114–3123. [Google Scholar] [CrossRef] [Green Version]
- Miller, A.R. Oxidation of cell wall polysaccharides by hydrogen peroxide: A potential mechanism for cell wall breakdown in plants. Biochem. Biophys. Res. Commun. 1986, 141, 238–244. [Google Scholar] [CrossRef]
- Schopfer, P. Hydrogen peroxide-mediated cell-wall stiffening in vitro in maize coleoptiles. Planta 1996, 199, 43–49. [Google Scholar] [CrossRef]
- Černý, M.; Habánová, H.; Berka, M.; Luklová, M.; Brzobohatý, B. Hydrogen peroxide: Its role in plant biology and crosstalk with signalling networks. Int. J. Mol. Sci. 2018, 19, 2812. [Google Scholar] [CrossRef] [Green Version]
- Mangano, S.; Denita-Juarez, S.P.; Choi, H.S.; Marzol, E.; Hwang, Y.; Ranocha, P.; Velasquez, S.M.; Borassi, C.; Barberini, M.L.; Aptekmann, A.A.; et al. Molecular link between auxin and ROS-mediated polar growth. PNAS 2017, 114, 5289–5294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millana Fañanás, E.; Todesca, S.; Sicorello, A.; Masino, L.; Pompach, P.; Magnani, F.; Pastore, A.; Mattevi, A. On the mechanism of calcium-dependent activation of NADPH oxidase 5 (NOX5). FEBS J. 2020, 287, 2486–2503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Im, S.H.; Klochkova, T.A.; Lee, D.J.; Gachon, C.M.; Kim, G.H. Genetic toolkits of the red alga Pyropia tenera against the three most common diseases in Pyropia farms. J Phycol. 2019, 55, 801–815. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. ROS are good. Trends Plant. Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moon, J.-S.; Hong, C.-Y.; Lee, J.-W.; Kim, G.-H. ROS Signaling Mediates Directional Cell Elongation and Somatic Cell Fusion in the Red Alga Griffithsia monilis. Cells 2022, 11, 2124. https://doi.org/10.3390/cells11132124
Moon J-S, Hong C-Y, Lee J-W, Kim G-H. ROS Signaling Mediates Directional Cell Elongation and Somatic Cell Fusion in the Red Alga Griffithsia monilis. Cells. 2022; 11(13):2124. https://doi.org/10.3390/cells11132124
Chicago/Turabian StyleMoon, Jong-Seok, Chan-Young Hong, Ji-Woong Lee, and Gwang-Hoon Kim. 2022. "ROS Signaling Mediates Directional Cell Elongation and Somatic Cell Fusion in the Red Alga Griffithsia monilis" Cells 11, no. 13: 2124. https://doi.org/10.3390/cells11132124
APA StyleMoon, J.-S., Hong, C.-Y., Lee, J.-W., & Kim, G.-H. (2022). ROS Signaling Mediates Directional Cell Elongation and Somatic Cell Fusion in the Red Alga Griffithsia monilis. Cells, 11(13), 2124. https://doi.org/10.3390/cells11132124