
Insights into Metabolic Activity and Structure of the Retina through Multiphoton Fluorescence Lifetime Imaging Microscopy in Mice



{kind=link}
{kind=link}
{kind=link}
{kind=link}
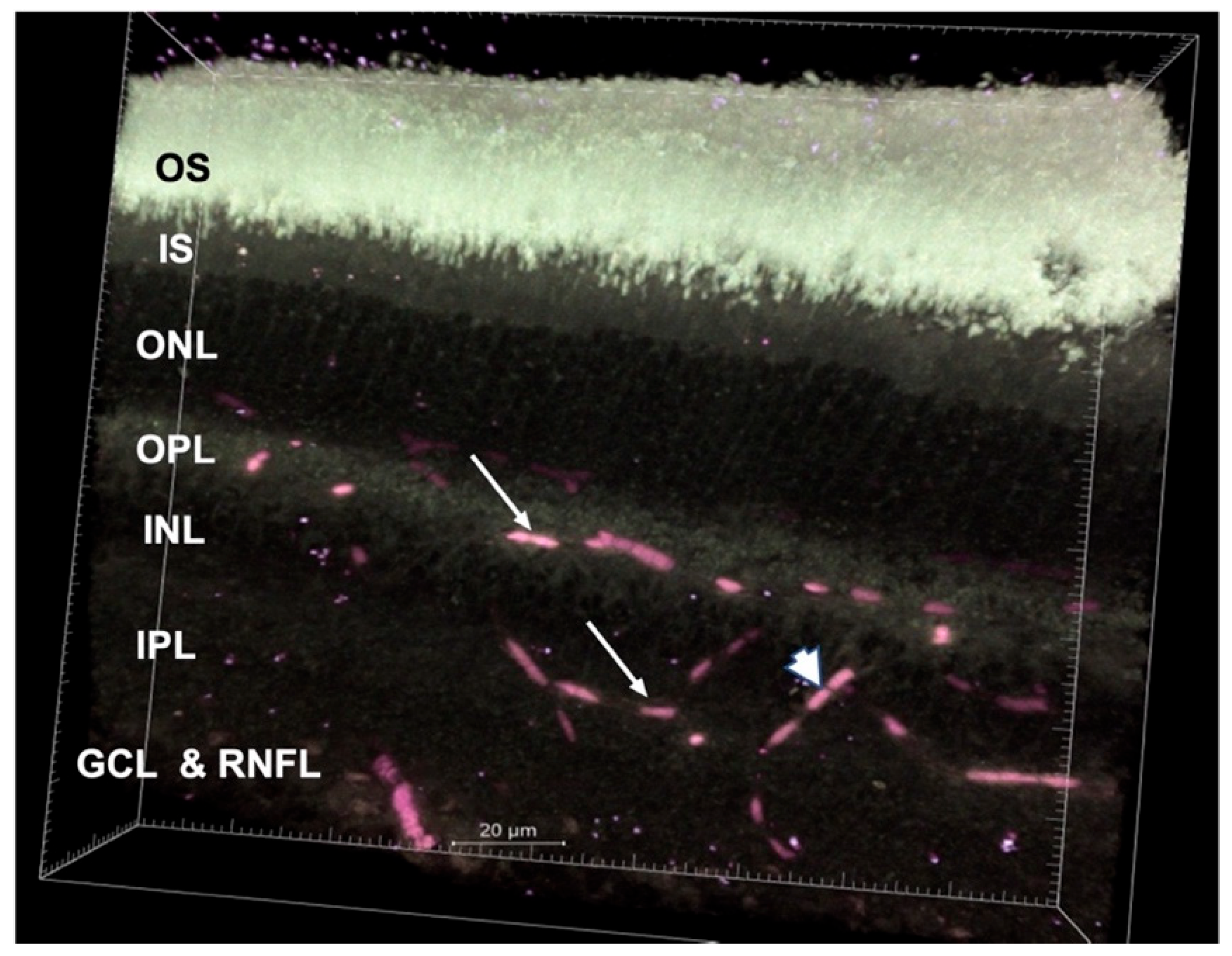{kind=link}
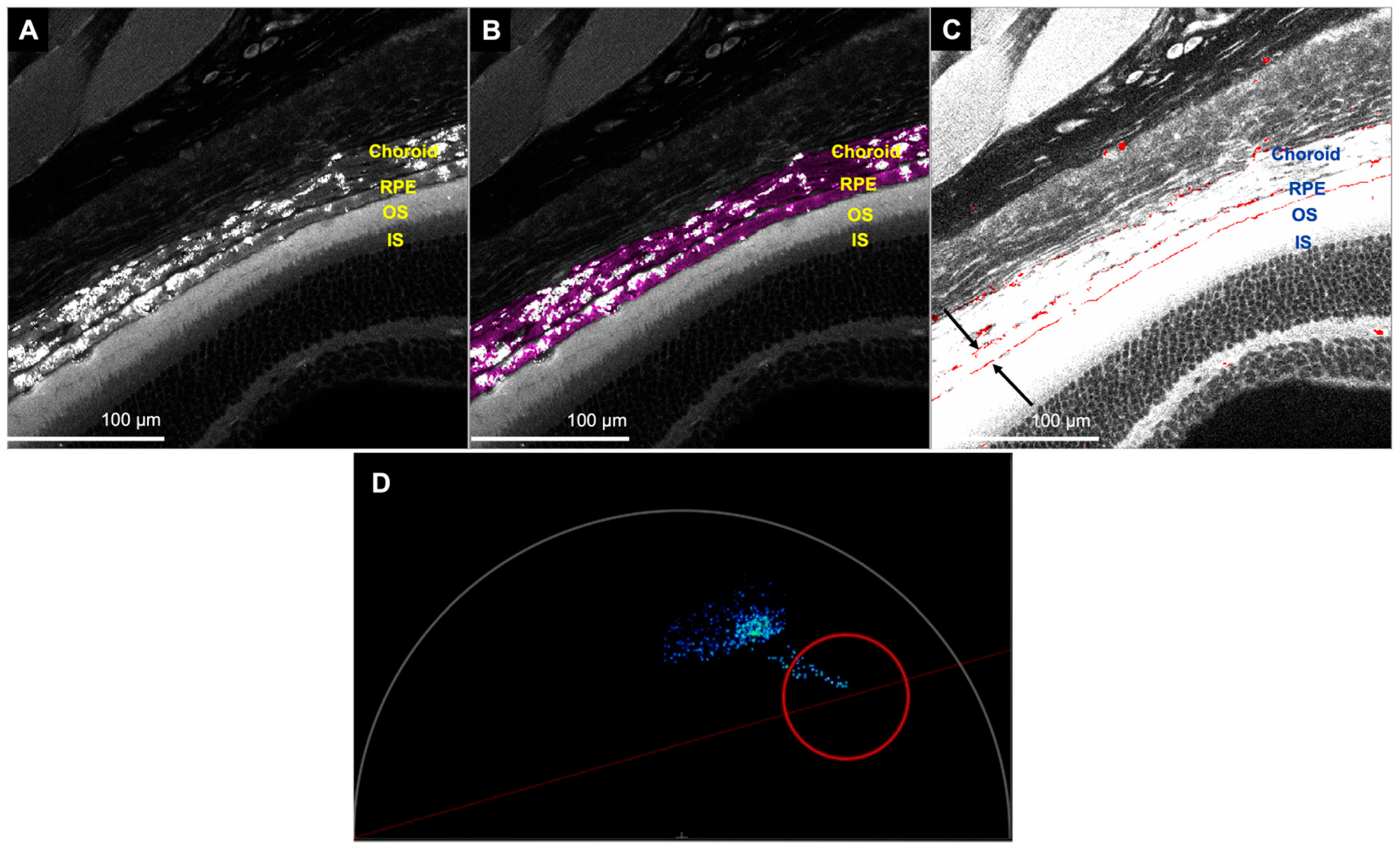{kind=link}
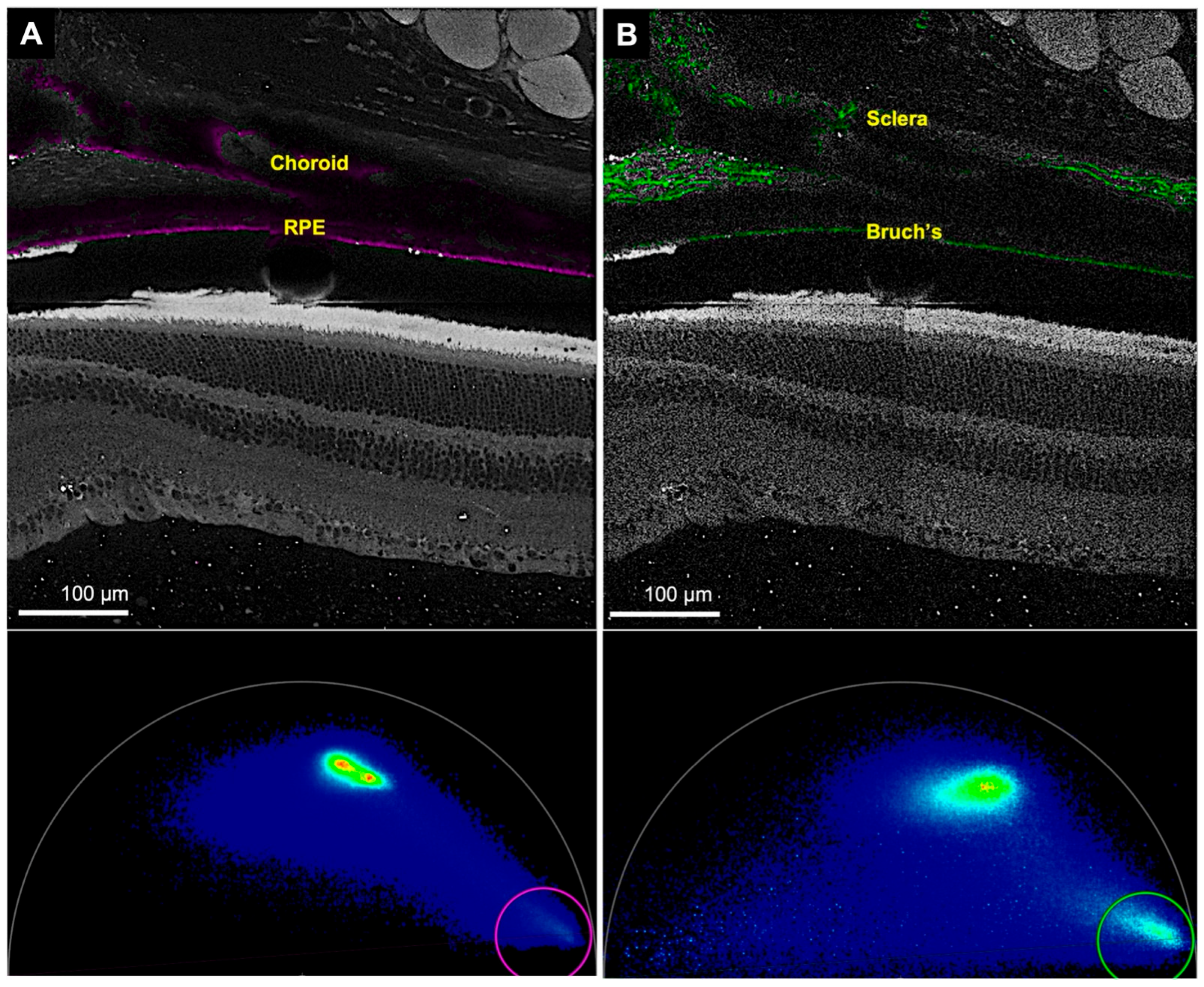{kind=link}
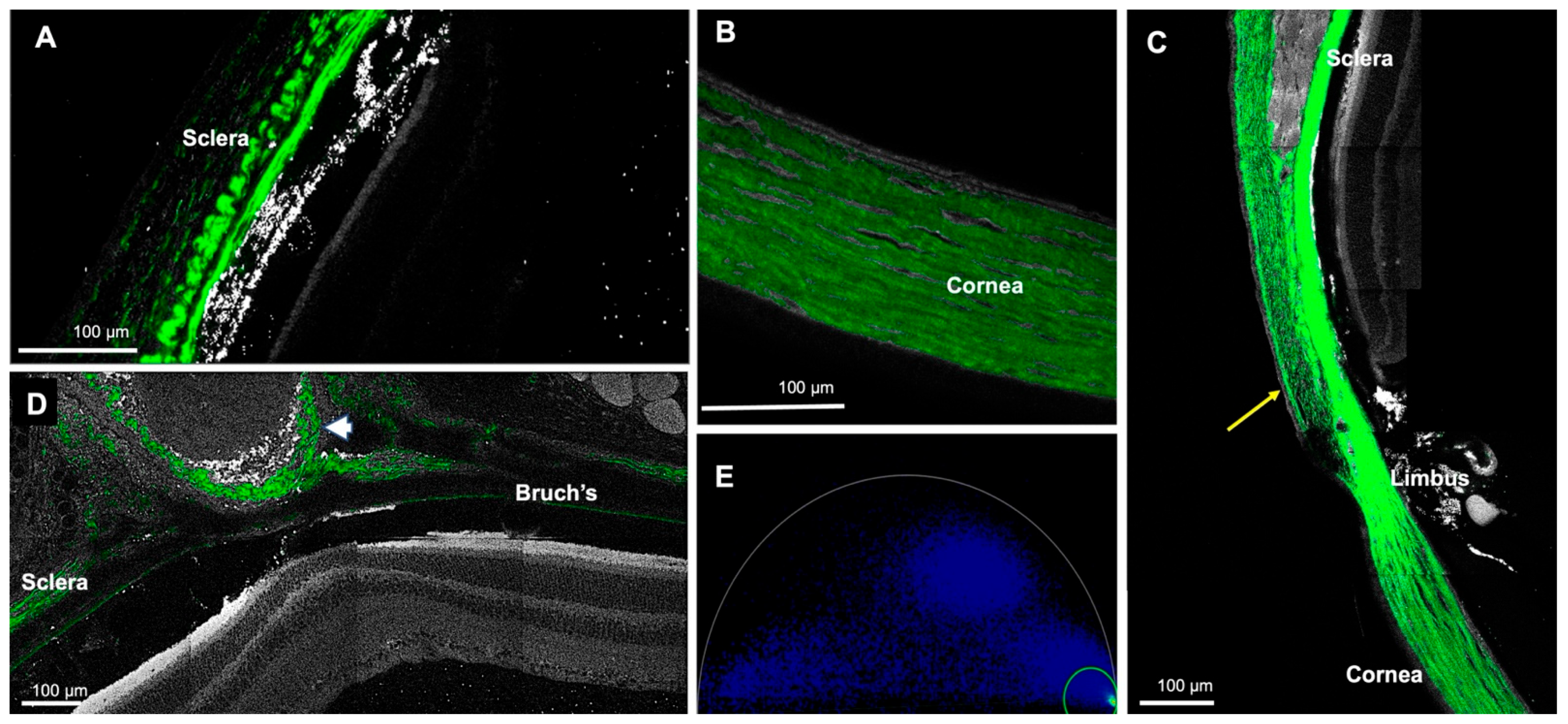{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Enucleation and Fixation of Eyes
2.3. Embedding and Sectioning
2.4. Multiphoton FLIM Imaging
2.5. FLIM Analysis
2.6. Statistical Analysis
3. Results
3.1. Metabolic Imaging
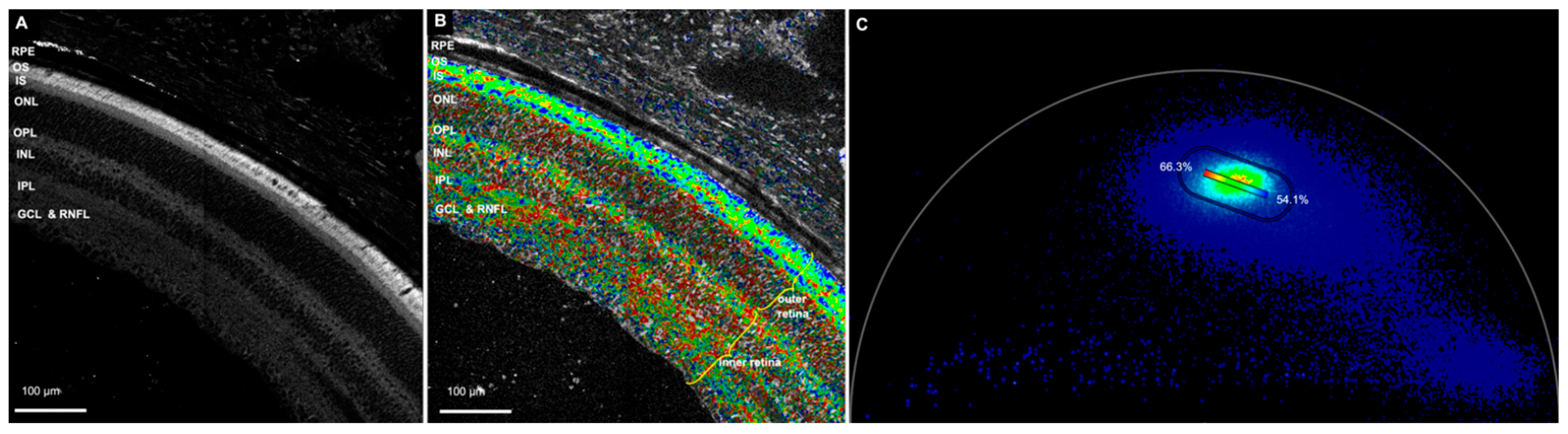
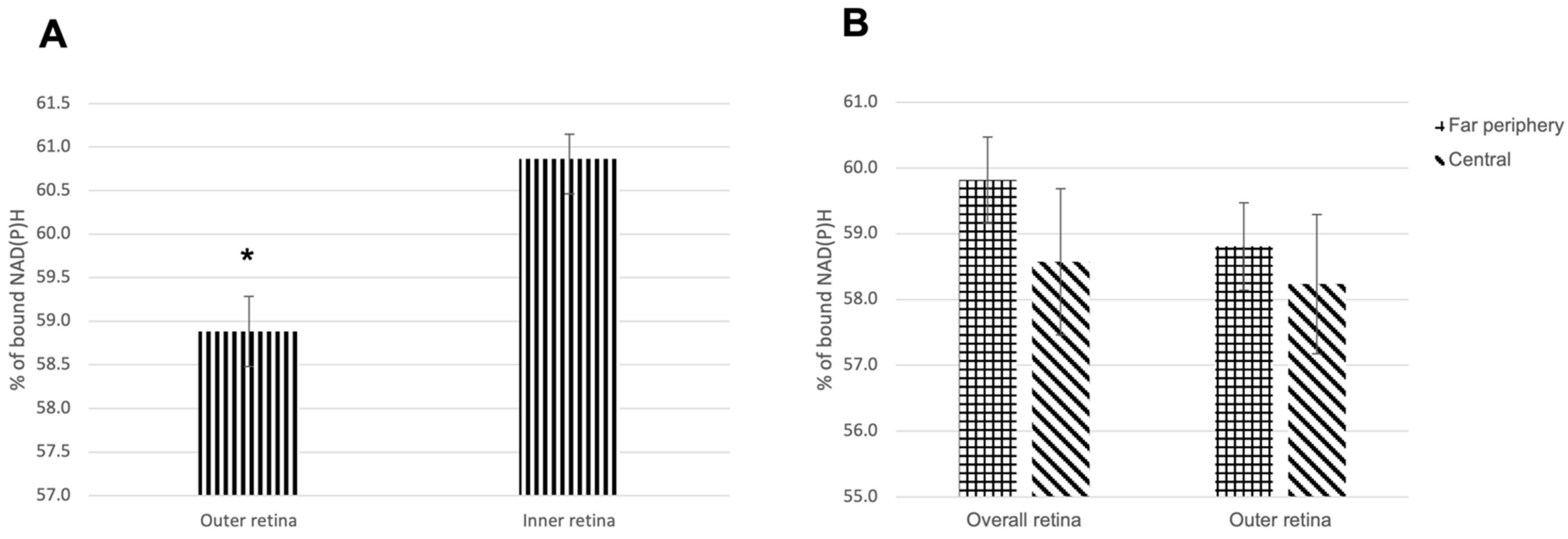
3.1.1. NAD(P)H Channel
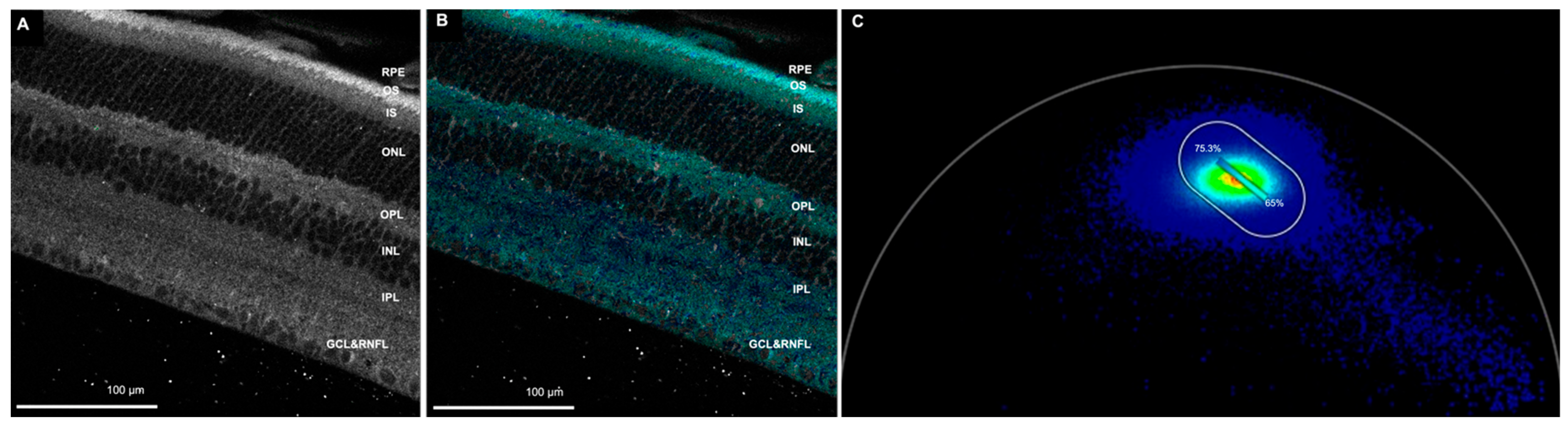
3.1.2. FAD Channel
3.2. Structural Imaging
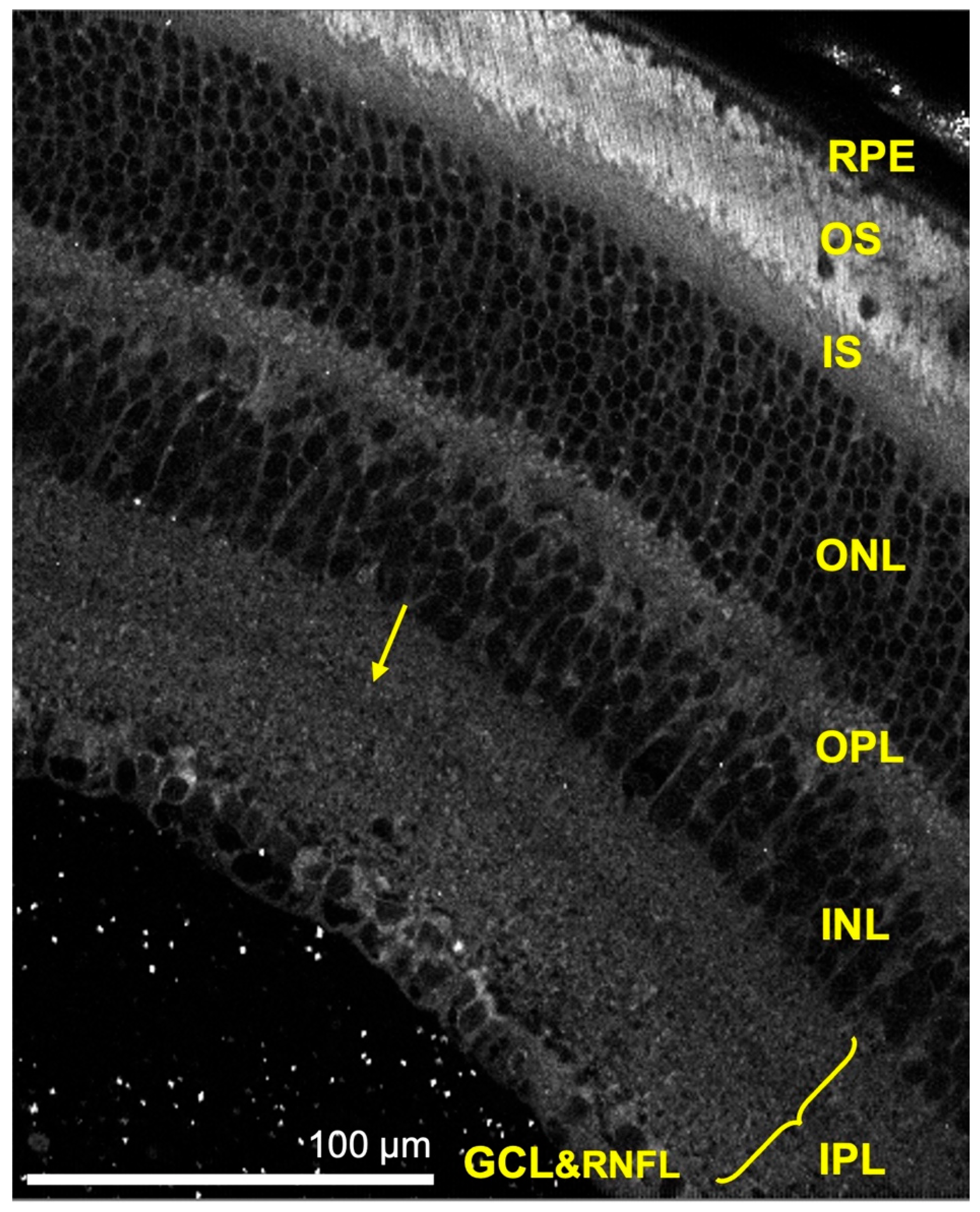
3.2.1. Hemoglobin and Melanin Channel
3.2.2. Collagen Channel
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lakowicz, J.R.; Szmacinski, H.; Nowaczyk, K.; Johnson, M.L. Fluorescence lifetime imaging of free and protein-bound NADH. Proc. Natl. Acad. Sci. USA 1992, 89, 1271–1275. [Google Scholar] [CrossRef] [PubMed]
- Skala, M.C.; Riching, K.M.; Gendron-Fitzpatrick, A.; Eickhoff, J.; Eliceiri, K.W.; White, J.G.; Ramanujam, N. In vivo multiphoton microscopy of NADH and FAD redox states, fluorescence lifetimes, and cellular morphology in precancerous epithelia. Proc. Natl. Acad. Sci. USA 2007, 104, 19494–19499. [Google Scholar] [CrossRef] [PubMed]
- Chance, B.; Baltscheffsky, H. Respiratory Enzymes in Oxidative Phosphorylation. J. Biol. Chem. 1958, 233, 736–739. [Google Scholar] [CrossRef]
- Browne, A.W.; Arnesano, C.; Harutyunyan, N.; Khuu, T.; Martinez, J.C.; Pollack, H.A.; Koos, D.S.; Lee, T.C.; Fraser, S.E.; Moats, R.A.; et al. Structural and Functional Characterization of Human Stem-Cell-Derived Retinal Organoids by Live Imaging. Investig. Opthalmology Vis. Sci. 2017, 58, 3311–3318. [Google Scholar] [CrossRef]
- Sameni, S.; Syed, A.; Marsh, J.L.; Digman, M.A. The phasor-FLIM fingerprints reveal shifts from OXPHOS to enhanced glycolysis in Huntington Disease. Sci. Rep. 2016, 6, 34755. [Google Scholar] [CrossRef]
- Coletta, V.C.; Rezende, C.A.; da Conceição, F.R.; Polikarpov, I.; Guimarães, F.E.G. Mapping the lignin distribution in pretreated sugarcane bagasse by confocal and fluorescence lifetime imaging microscopy. Biotechnol. Biofuels 2013, 6, 43. [Google Scholar] [CrossRef]
- Denk, W.; Strickler, J.H.; Webb, W.W. Two-photon laser scanning fluorescence microscopy. Science 1990, 248, 73–76. [Google Scholar] [CrossRef]
- Stelzer, E.H.; Hell, S.; Lindek, S.; Stricker, R.; Pick, R.; Storz, C.; Ritter, G.; Salmon, N. Nonlinear absorption extends confocal fluorescence microscopy into the ultra-violet regime and confines the illumination volume. Opt. Commun. 1994, 104, 223–228. [Google Scholar] [CrossRef]
- Centonze, V.E.; White, J.G. Multiphoton Excitation Provides Optical Sections from Deeper within Scattering Specimens than Confocal Imaging. Biophys. J. 1998, 75, 2015–2024. [Google Scholar] [CrossRef]
- Schweitzer, D.; Schenke, S.; Hammer, M.; Schweitzer, F.; Jentsch, S.; Birckner, E.; Becker, W.; Bergmann, A. Towards metabolic mapping of the human retina. Microsc. Res. Tech. 2007, 70, 410–419. [Google Scholar] [CrossRef]
- Schweitzer, D.; Hammer, M.; Schweitzer, F.; Anders, R.; Doebbecke, T.; Schenke, S.; Gaillard, E.R.; Gaillard, E.R. In vivo measurement of time-resolved autofluorescence at the human fundus. J. Biomed. Opt. 2004, 9, 1214–1222. [Google Scholar] [CrossRef] [PubMed]
- Dysli, C.; Schuerch, K.; Escher, P.; Wolf, S.; Zinkernagel, M.S. Fundus Autofluorescence Lifetime Patterns in Retinitis Pigmentosa. Investig. Opthalmol. Vis. Sci. 2018, 59, 1769–1778. [Google Scholar] [CrossRef] [PubMed]
- Dysli, C.-S.; Dysli, M.; Enzmann, V.; Wolf, S.; Zinkernagel, M.S. Fluorescence Lifetime Imaging of the Ocular Fundus in Mice. Investig. Opthalmol. Vis. Sci. 2014, 55, 7206–7215. [Google Scholar] [CrossRef] [PubMed]
- Peters, S.; Hammer, M.; Schweitzer, D. Two-photon excited fluorescence microscopy application for ex vivo investigation of ocular fundus samples. In Proceedings of the European Conferences on Biomedical Optics, Munich, Germany, 22–26 May 2011; Volume 8086, p. 808605. [Google Scholar] [CrossRef]
- Batista, A.; Breunig, H.G.; König, A.; Morgado, A.M.; König, K. Assessment of the metabolism and morphology of the porcine cornea, lens and retina by 2-photon imaging. J. Biophotonics 2018, 11, e201700324. [Google Scholar] [CrossRef] [PubMed]
- Hayaran, A.; Bijlani, V. Polyacrylamide as an infiltrating and embedding medium for vibratome sectioning of human fetal cerebellum containing Dil-filled axons. J. Neurosci. Methods 1992, 42, 65–68. [Google Scholar] [CrossRef]
- Zipfel, W.R.; Williams, R.M.; Christie, R.; Nikitin, A.Y.; Hyman, B.T.; Webb, W.W. Live tissue intrinsic emission microscopy using multiphoton-excited native fluorescence and second harmonic generation. Proc. Natl. Acad. Sci. USA 2003, 100, 7075–7080. [Google Scholar] [CrossRef]
- Ranjit, S.; Dvornikov, A.; Stakic, M.; Hong, S.-H.; Levi, M.; Evans, R.M.; Gratton, E. Imaging Fibrosis and Separating Collagens using Second Harmonic Generation and Phasor Approach to Fluorescence Lifetime Imaging. Sci. Rep. 2015, 5, srep13378. [Google Scholar] [CrossRef]
- Gao, S.; Lan, X.; Liu, Y.; Shen, Z.; Lu, J.; Ni, X. Characteristics of blood fluorescence spectra using low-level, 457.9-nm excitation from Ar+ laser. Chin. Opt. Lett. 2004, 2, 160–161. [Google Scholar]
- Digman, M.A.; Caiolfa, V.R.; Zamai, M.; Gratton, E. The Phasor Approach to Fluorescence Lifetime Imaging Analysis. Biophys. J. 2008, 94, L14–L16. [Google Scholar] [CrossRef]
- Stringari, C.; Cinquin, A.; Cinquin, O.; Digman, M.A.; Donovan, P.J.; Gratton, E. Phasor approach to fluorescence lifetime microscopy distinguishes different metabolic states of germ cells in a live tissue. Proc. Natl. Acad. Sci. USA 2011, 108, 13582–13587. [Google Scholar] [CrossRef]
- Sitiwin, E.; Madigan, M.C.; Gratton, E.; Cherepanoff, S.; Conway, R.M.; Whan, R.; Macmillan, A. Shedding light on melanins within in situ human eye melanocytes using 2-photon microscopy profiling techniques. Sci. Rep. 2019, 9, 18585. [Google Scholar] [CrossRef] [PubMed]
- Roth, S.; Freund, I. Second harmonic generation in collagen. J. Chem. Phys. 1979, 70, 1637–1643. [Google Scholar] [CrossRef]
- Chacko, J.V.; Eliceiri, K.W. NAD(P)H fluorescence lifetime measurements in fixed biological tissues. Methods Appl. Fluoresc. 2019, 7, 044005. [Google Scholar] [CrossRef] [PubMed]
- Rueda, E.M.; Johnson, J.E., Jr.; Giddabasappa, A.; Swaroop, A.; Brooks, M.J.; Sigel, I.; Chaney, S.Y.; Fox, D.A. The cellular and compartmental profile of mouse retinal glycolysis, tricarboxylic acid cycle, oxidative phosphorylation, and ~P transferring kinases. Mol. Vis. 2016, 22, 847–885. [Google Scholar]
- Volland, S.; Esteve-Rudd, J.; Hoo, J.; Yee, C.; Williams, D.S. A Comparison of Some Organizational Characteristics of the Mouse Central Retina and the Human Macula. PLoS ONE 2015, 10, e0125631. [Google Scholar] [CrossRef]
- Dräger, U.C.; Olsen, J.F. Origins of crossed and uncrossed retinal projections in pigmented and albino mice: Mouse Retinal Ganglion Cells. J. Comp. Neurol. 1980, 191, 383–412. [Google Scholar] [CrossRef]
- Alam, S.R.; Wallrabe, H.; Svindrych, Z.; Chaudhary, A.K.; Christopher, K.G.; Chandra, D.; Periasamy, A. Investigation of Mitochondrial Metabolic Response to Doxorubicin in Prostate Cancer Cells: An NADH, FAD and Tryptophan FLIM Assay. Sci. Rep. 2017, 7, 2977. [Google Scholar] [CrossRef]
- Cao, R.; Wallrabe, H.K.; Periasamy, A. Multiphoton FLIM imaging of NAD(P)H and FAD with one excitation wavelength. J. Biomed. Opt. 2020, 25, 014510. [Google Scholar] [CrossRef]
- Yamagata, M.; Sanes, J.R. Expression and Roles of the Immunoglobulin Superfamily Recognition Molecule Sidekick1 in Mouse Retina. Front. Mol. Neurosci. 2019, 11, 485. [Google Scholar] [CrossRef]
- Contini, M.; Lin, B.; Kobayashi, K.; Okano, H.; Masland, R.H.; Raviola, E. Synaptic input of ON-bipolar cells onto the dopaminergic neurons of the mouse retina. J. Comp. Neurol. 2010, 518, 2035–2050. [Google Scholar] [CrossRef]
- Knop, G.C.; Feigenspan, A.; Weiler, R.; Dedek, K. Inputs Underlying the ON–OFF Light Responses of Type 2 Wide-Field Amacrine Cells in TH::GFP Mice. J. Neurosci. 2011, 31, 4780–4791. [Google Scholar] [CrossRef]
- Clark, M.A.; Worrell, M.B.; Pless, J.E. Postmortem Changes in Soft Tissues. In Forensic Taphonomy: The Postmortem Fate of Human Remains; Haglund, W.D., Sorg, M.H., Eds.; CRC Press: Boca Raton, FL, USA, 1997; pp. 156–164. [Google Scholar]
- Prahst, C.; Ashrafzadeh, P.; Mead, T.; Figueiredo, A.; Chang, K.; Richardson, D.; Venkaraman, L.; Richards, M.; Russo, A.M.; Harrington, K.; et al. Mouse retinal cell behaviour in space and time using light sheet fluorescence microscopy. eLife 2020, 9, e49779. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.-H.; Hsiao, G.; Al-Shabrawey, M. Eicosanoids and Oxidative Stress in Diabetic Retinopathy. Antioxidants 2020, 9, 520. [Google Scholar] [CrossRef] [PubMed]
- Giannakaki-Zimmermann, H.; Kokona, D.; Wolf, S.; Ebneter, A.; Zinkernagel, M.S. Optical Coherence Tomography Angiography in Mice: Comparison with Confocal Scanning Laser Microscopy and Fluorescein Angiography. Transl. Vis. Sci. Technol. 2016, 5, 11. [Google Scholar] [CrossRef] [PubMed]
- Tezel, T.H.; Geng, L.; Lato, E.B.; Schaal, S.; Liu, Y.; Dean, D.; Klein, J.B.; Kaplan, H.J.; Ellis, D.Z.; Dismuke, W.M.; et al. Synthesis and Secretion of Hemoglobin by Retinal Pigment Epithelium. Investig. Opthalmol. Vis. Sci. 2009, 50, 1911–1919. [Google Scholar] [CrossRef]
- Cox, G.; Kable, E.; Jones, A.; Fraser, I.; Manconi, F.; Gorrell, M.D. 3-Dimensional imaging of collagen using second harmonic generation. J. Struct. Biol. 2003, 141, 53–62. [Google Scholar] [CrossRef]
- Harnisch, J.-P.; Buchen, R.; Sinha, P.K.; Barrach, H.J. Ultrastructural identification of type I and II collagen in the cornea of the mouse by means of enzyme labeled antibodies. Albrecht Von Graefes Arch. Für Klin. Exp. Ophthalmol. 1978, 208, 9–13. [Google Scholar] [CrossRef]
- Keeley, F.W.; Morin, J.D.; Vesely, S. Characterization of collagen from normal human sclera. Exp. Eye Res. 1984, 39, 533–542. [Google Scholar] [CrossRef]
- Park, C.Y.; Marando, C.; Liao, J.A.; Lee, J.K.; Kwon, J.; Chuck, R.S. Details of the Collagen and Elastin Architecture in the Human Limbal Conjunctiva, Tenon's Capsule and Sclera Revealed by Two-Photon Excited Fluorescence Microscopy. Investig. Opthalmology Vis. Sci. 2016, 57, 5602–5610. [Google Scholar] [CrossRef]
- Zyablitskaya, M.; Takaoka, A.; Munteanu, E.L.; Nagasaki, T.; Trokel, S.L.; Paik, D.C. Evaluation of Therapeutic Tissue Crosslinking (TXL) for Myopia Using Second Harmonic Generation Signal Microscopy in Rabbit Sclera. Investig. Opthalmol. Vis. Sci. 2017, 58, 21–29. [Google Scholar] [CrossRef][Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kesavamoorthy, N.; Junge, J.A.; Fraser, S.E.; Ameri, H. Insights into Metabolic Activity and Structure of the Retina through Multiphoton Fluorescence Lifetime Imaging Microscopy in Mice. Cells 2022, 11, 2265. https://doi.org/10.3390/cells11152265
Kesavamoorthy N, Junge JA, Fraser SE, Ameri H. Insights into Metabolic Activity and Structure of the Retina through Multiphoton Fluorescence Lifetime Imaging Microscopy in Mice. Cells. 2022; 11(15):2265. https://doi.org/10.3390/cells11152265
Chicago/Turabian StyleKesavamoorthy, Niranjana, Jason A. Junge, Scott E. Fraser, and Hossein Ameri. 2022. "Insights into Metabolic Activity and Structure of the Retina through Multiphoton Fluorescence Lifetime Imaging Microscopy in Mice" Cells 11, no. 15: 2265. https://doi.org/10.3390/cells11152265
APA StyleKesavamoorthy, N., Junge, J. A., Fraser, S. E., & Ameri, H. (2022). Insights into Metabolic Activity and Structure of the Retina through Multiphoton Fluorescence Lifetime Imaging Microscopy in Mice. Cells, 11(15), 2265. https://doi.org/10.3390/cells11152265