
Hydrogen Sulfide Improves Angiogenesis by Regulating the Transcription of pri-miR-126 in Diabetic Endothelial Cells
Abstract
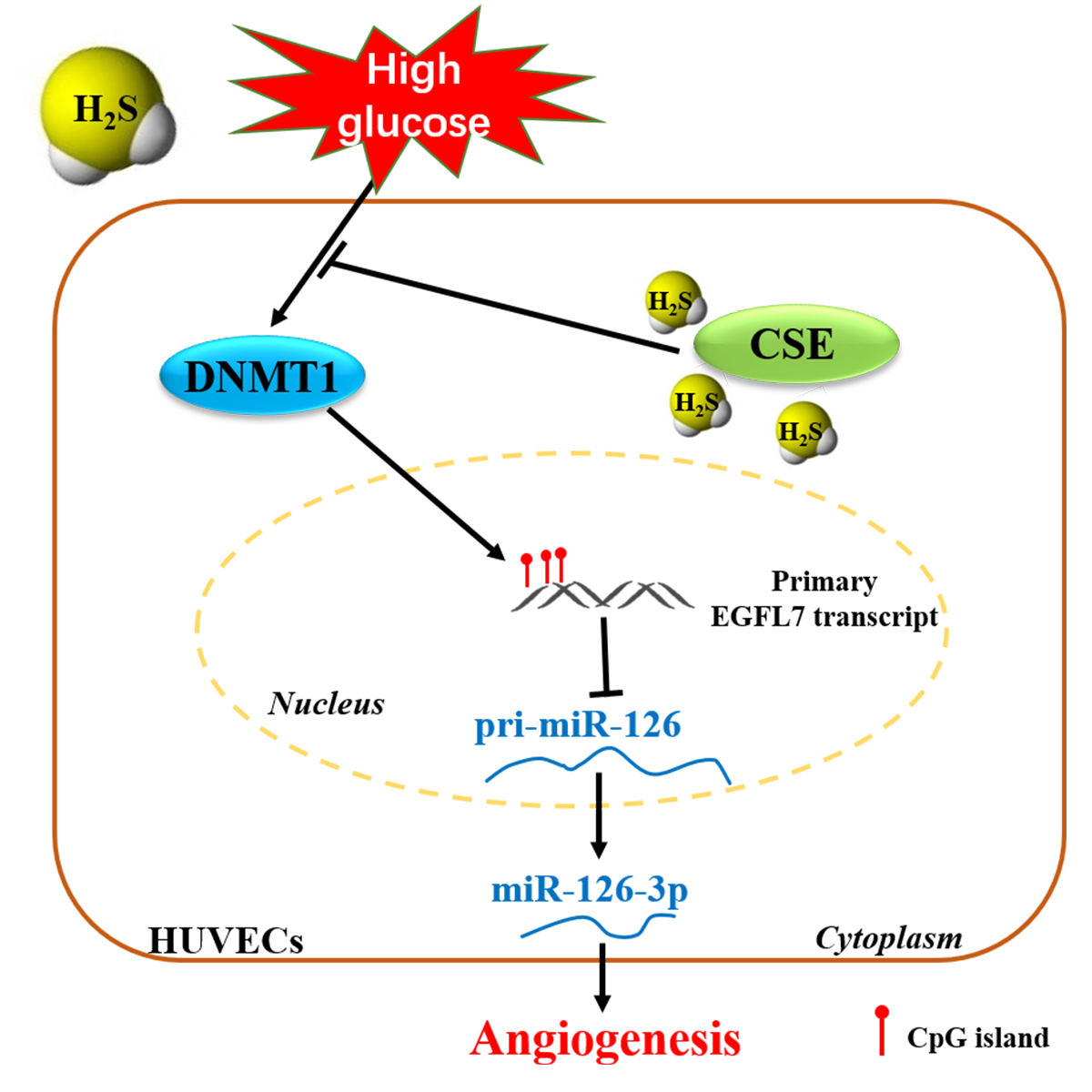
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Modeling of Type I Diabetic Mice
2.2. Murine Hindlimb Ischemia Model
2.3. Immunofluorescence Assay
2.4. GEO Datasets of MicroRNA between the Diabetic and Control Groups
2.5. Data Preprocess and Differential MicroRNA Expression Analysis
2.6. Affymetrix MicroRNA Profiling
2.7. Reverse Transcription and Real-Time PCR
2.8. In Vivo Matrigel Plugs Analysis
2.9. Cell Culture
2.10. Cell Transfection and Lentivirus Infection
2.11. Measurements of the H2S Level
2.12. Scratch Wound-Healing Assay
2.13. Methylated DNA Immunoprecipitation (MeIP)
2.14. Ethics Approval and Consent to Participate
2.15. Statistical Analysis
3. Results
3.1. Exogenous H2S Improved Angiogenesis in Diabetic Mice
3.2. MicroRNAs were Regulated by Both Diabetes and H2S
3.3. Exogenous H2S Regulated the miR-126-3p and DNMT1 Levels in the Ischemic Gastrocnemius Muscles of Diabetic Mice
3.4. miR-126-3p Was Downstream of DNMT1 to Regulate Angiogenesis in Endothelial Cells
3.5. CSE Overexpression Increased the Transcription of pri-miR-126 under High-Glucose Conditions
3.6. CSE Overexpression Rescued the miR-126-3p Level through Regulating DNMT1 Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, J.; Hale, N. Evidence and Implications of the Affordable Care Act for Racial/Ethnic Disparities in Diabetes Health During and Beyond the Pandemic. Popul. Health Manag. 2022, 25, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.C.W. Epidemiology of diabetes and diabetic complications in China. Diabetologia 2018, 61, 1249–1260. [Google Scholar] [CrossRef] [PubMed]
- Willyard, C. Limb-saving medicines sought to prevent amputations. Nat. Med. 2012, 18, 328. [Google Scholar] [CrossRef] [PubMed]
- Jonasson, J.M.; Ye, W.; Sparén, P.; Apelqvist, J.; Nyrén, O.; Brismar, K. Risks of Nontraumatic Lower-Extremity Amputations in Patients with Type 1 Diabetes: A population-based cohort study in Sweden. Diabetes Care 2008, 31, 1536–1540. [Google Scholar] [CrossRef] [PubMed]
- Sumpio, B.E.; Riley, J.T.; Dardik, A. Cells in focus: Endothelial cell. Int. J. Biochem. Cell Biol. 2002, 34, 1508–1512. [Google Scholar] [CrossRef]
- Kaur, R.; Kaur, M.; Singh, J. Endothelial dysfunction and platelet hyperactivity in type 2 diabetes mellitus: Molecular insights and therapeutic strategies. Cardiovasc. Diabetol. 2018, 17, 1–17. [Google Scholar] [CrossRef]
- Dinh, T.; Scovell, S.; Veves, A. Review: Peripheral Arterial Disease and Diabetes: A Clinical Update. Int. J. Low. Extremity Wounds 2009, 8, 75–81. [Google Scholar] [CrossRef]
- Łowicka, E.; Bełtowski, J. Hydrogen sulfide (H2S)—the third gas of interest for pharmacologists. Pharmacol. Rep. 2007, 59, 4–24. [Google Scholar]
- Suzuki, K.; Sagara, M.; Aoki, C.; Tanaka, S.; Aso, Y. Clinical Implication of Plasma Hydrogen Sulfide Levels in Japanese Patients with Type 2 Diabetes. Intern. Med. 2017, 56, 17–21. [Google Scholar] [CrossRef]
- Dutta, M.; Biswas, U.K.; Chakraborty, R.; Banerjee, P.; Raychaudhuri, U.; Kumar, A. Evaluation of plasma H2S levels and H2S synthesis in streptozotocin induced Type-2 diabetes-an experimental study based on Swietenia macrophylla seeds. Asian Pac. J. Trop. Biomed. 2014, 4, S483–S487. [Google Scholar] [CrossRef]
- Cai, W.-J.; Wang, M.-J.; Moore, P.K.; Jin, H.-M.; Yao, T.; Zhu, Y.-C. The novel proangiogenic effect of hydrogen sulfide is dependent on Akt phosphorylation. Cardiovasc. Res. 2007, 76, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, W.; Chen, Q.; Jiang, Y.; Lu, X.; Zhao, X. Hydrogen sulfide accelerates wound healing in diabetic rats. Int. J. Clin. Exp. Pathol. 2015, 8, 5097–5104. [Google Scholar] [PubMed]
- Zhao, H.; Lu, S.; Chai, J.; Zhang, Y.; Ma, X.; Chen, J.; Guan, Q.; Wan, M.; Liu, Y. Hydrogen sulfide improves diabetic wound healing in ob/ob mice via attenuating inflammation. J. Diabetes Its Complicat. 2017, 31, 1363–1369. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-G.; Li, W. Hydrogen sulfide improves vessel formation of the ischemic adductor muscle and wound healing in diabetic db/db mice. Iran J. Basic Med. Sci. 2019, 22, 1192–1197. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Chen, D.-D.; Sun, X.; Xie, H.-H.; Yuan, H.; Jia, W.; Chen, A.F. Hydrogen Sulfide Improves Wound Healing via Restoration of Endothelial Progenitor Cell Functions and Activation of Angiopoietin-1 in Type 2 Diabetes. Diabetes 2014, 63, 1763–1778. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, X.-H.; Xue, W.-L.; Jin, S.; Li, M.-Y.; Zhang, C.-C.; Yu, B.; Zhu, L.; Liang, K.; Chen, M.Y.; et al. YB-1 Recruits Drosha to Promote Splicing of pri-miR-192 to Mediate the Proangiogenic Effects of H2S. Antioxidants Redox Signal. 2022, 36, 760–783. [Google Scholar] [CrossRef]
- Icli, B.; Wu, W.; Ozdemir, D.; Li, H.; Cheng, H.S.; Haemmig, S.; Liu, X.; Giatsidis, G.; Avci, S.N.; Lee, N.; et al. MicroRNA-615-5p Regulates Angiogenesis and Tissue Repair by Targeting AKT/eNOS (Protein Kinase B/Endothelial Nitric Oxide Synthase) Signaling in Endothelial Cells. Arter. Thromb. Vasc. Biol. 2019, 39, 1458–1474. [Google Scholar] [CrossRef]
- Wang, S.; Aurora, A.B.; Johnson, B.A.; Qi, X.; McAnally, J.; Hill, J.A.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. The Endothelial-Specific MicroRNA miR-126 Governs Vascular Integrity and Angiogenesis. Dev. Cell 2008, 15, 261–271. [Google Scholar] [CrossRef]
- Chistiakov, D.A.; Orekhov, A.N.; Bobryshev, Y.V. The role of miR-126 in embryonic angiogenesis, adult vascular homeostasis, and vascular repair and its alterations in atherosclerotic disease. J. Mol. Cell. Cardiol. 2016, 97, 47–55. [Google Scholar] [CrossRef]
- Bassand, K.; Metzinger, L.; Naïm, M.; Mouhoubi, N.; Haddad, O.; Assoun, V.; Zaïdi, N.; Sainte-Catherine, O.; Butt, A.; Guyot, E.; et al. miR-126-3p is essential for CXCL12-induced angiogenesis. J. Cell. Mol. Med. 2021, 25, 6032–6045. [Google Scholar] [CrossRef]
- Zhou, Q.; Anderson, C.; Hanus, J.; Zhao, F.; Ma, J.; Yoshimura, A.; Wang, S. Strand and Cell Type-specific Function of microRNA-126 in Angiogenesis. Mol. Ther. 2016, 24, 1823–1835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, H.; Peng, C.; Wu, L.; Gao, S.; Wang, Z.; Dai, L.; Wu, H. Yiqi-Huoxue granule promotes angiogenesis of ischemic myocardium through miR-126/PI3K/Akt axis in endothelial cells. Phytomedicine 2021, 92, 153713. [Google Scholar] [CrossRef]
- Kuhnert, F.; Mancuso, M.R.; Hampton, J.; Stankunas, K.; Asano, T.; Chen, C.-Z.; Kuo, C.J. Attribution of vascular phenotypes of the murine Egfl7 locus to the microRNA miR-126. Development 2008, 135, 3989–3993. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.J.; Rosenblat, J.D.; Roth, N.C.; Kuliszewski, M.A.; Matkar, P.N.; Rudenko, D.; Liao, C.; Lee, P.J.; Leong-Poi, H. Therapeutic Angiogenesis by Ultrasound-Mediated MicroRNA-126-3p Delivery. Arter. Thromb. Vasc. Biol. 2015, 35, 2401–2411. [Google Scholar] [CrossRef] [PubMed]
- Hackfort, B.T.; Mishra, P.K. Emerging role of hydrogen sulfide-microRNA crosstalk in cardiovascular diseases. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H802–H812. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Wang, Q.; Chen, Y.; Ma, Y.; Wang, D. Involvement of MicroRNA-133a in the Protective Effect of Hydrogen Sulfide against Ischemia/Reperfusion-Induced Endoplasmic Reticulum Stress and Cardiomyocyte Apoptosis. Pharmacology 2018, 103, 1–9. [Google Scholar] [CrossRef]
- Liu, Y.; Liao, S.; Quan, H.; Lin, Y.; Li, J.; Yang, Q. Involvement of microRNA-135a-5p in the Protective Effects of Hydrogen Sulfide Against Parkinson’s Disease. Cell. Physiol. Biochem. 2016, 40, 18–26. [Google Scholar] [CrossRef]
- Wang, C.; Ma, X.; Zhang, J.; Jia, X.; Huang, M. DNMT1 maintains the methylation of miR -152-3p to regulate TMSB10 expression, thereby affecting the biological characteristics of colorectal cancer cells. IUBMB Life 2020, 72, 2432–2443. [Google Scholar] [CrossRef]
- Balakrishnan, A.; Guruprasad, K.P.; Satyamoorthy, K.; Joshi, M.B. Interleukin-6 determines protein stabilization of DNA methyltransferases and alters DNA promoter methylation of genes associated with insulin signaling and angiogenesis. Lab. Investig. 2018, 98, 1143–1158. [Google Scholar] [CrossRef]
- Zhao, J.; Yang, S.; Shu, B.; Chen, L.; Yang, R.; Xu, Y.; Xie, J.; Liu, X.; Qi, S. Transient High Glucose Causes Persistent Vascular Dysfunction and Delayed Wound Healing by the DNMT1-Mediated Ang-1/NF-kappaB Pathway. J. Investig. Dermatol. 2020, 141, 1573–1584. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Q.; Liu, S.; Chen, Y.; Li, R.; Lin, T.; Yu, C.; Zhang, H.; Huang, Z.; Zhao, X.; et al. DNA methyltransferase 1 may be a therapy target for attenuating diabetic nephropathy and podocyte injury. Kidney Int. 2017, 92, 140–153. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.-L.; Chen, R.-Q.; Zhang, Q.-Q.; Li, X.-H.; Cao, L.; Li, M.-Y.; Li, Y.; Lin, G.; Chen, Y.; Wang, M.-J.; et al. Hydrogen sulfide rescues high glucose-induced migration dysfunction in HUVECs by upregulating miR-126-3p. Am. J. Physiol.Cell Physiol. 2020, 318, C857–C869. [Google Scholar] [CrossRef] [PubMed]
- Losordo, D.W.; Isner, J.M. Estrogen and angiogenesis: A review. Arter. Thromb. Vasc. Biol. 2001, 21, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Niiyama, H.; Huang, N.; Rollins, M.D.; Cooke, J.P. Murine Model of Hindlimb Ischemia. J. Vis. Exp. 2009, 21, e1035. [Google Scholar] [CrossRef]
- Whiteman, M.; Moore, P.K. Hydrogen sulfide and the vasculature: A novel vasculoprotective entity and regulator of nitric oxide bioavailability? J. Cell. Mol. Med. 2009, 13, 488–507. [Google Scholar] [CrossRef]
- Smit-McBride, Z.; Nguyen, A.T.; Yu, A.K.; Modjtahedi, S.P.; Hunter, A.A.; Rashid, S.; Moisseiev, E.; Morse, L.S. Unique molecular signatures of microRNAs in ocular fluids and plasma in diabetic retinopathy. PLoS ONE 2020, 15, e0235541. [Google Scholar] [CrossRef]
- Bonauer, A.; Carmona, G.; Iwasaki, M.; Mione, M.; Koyanagi, M.; Fischer, A.; Burchfield, J.; Fox, H.; Doebele, C.; Ohtani, K.; et al. MicroRNA-92a Controls Angiogenesis and Functional Recovery of Ischemic Tissues in Mice. Science 2009, 324, 1710–1713. [Google Scholar] [CrossRef]
- Jiang, L.; Jia, M.; Wei, X.; Guo, J.; Hao, S.; Mei, A.; Zhi, X.; Wang, X.; Li, Q.; Jin, J.; et al. Bach1-induced suppression of angiogenesis is dependent on the BTB domain. eBioMedicine 2020, 51, 102617. [Google Scholar] [CrossRef]
- Sasakura, K.; Hanaoka, K.; Shibuya, N.; Mikami, Y.; Kimura, Y.; Komatsu, T.; Ueno, T.; Terai, T.; Kimura, H.; Nagano, T. Development of a Highly Selective Fluorescence Probe for Hydrogen Sulfide. J. Am. Chem. Soc. 2011, 133, 18003–18005. [Google Scholar] [CrossRef]
- Thu, K.L.; Vucic, E.A.; Kennett, J.Y.; Heryet, C.; Brown, C.; Lam, W.L.; Wilson, I.M. Methylated DNA Immunoprecipitation. J. Vis. Exp. 2009, e935. [Google Scholar] [CrossRef]
- Whiteman, M.; Gooding, K.M.; Whatmore, J.L.; Ball, C.I.; Mawson, D.; Skinner, K.; Tooke, J.E.; Shore, A.C. Adiposity is a major determinant of plasma levels of the novel vasodilator hydrogen sulphide. Diabetologia 2010, 53, 1722–1726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limbourg, A.; Korff, T.; Napp, L.C.; Schaper, W.; Drexler, H.; Limbourg, F. Evaluation of postnatal arteriogenesis and angiogenesis in a mouse model of hind-limb ischemia. Nat. Protoc. 2009, 4, 1737–1748. [Google Scholar] [CrossRef]
- Coletta, C.; Módis, K.; Szczesny, B.; Brunyánszki, A.; Oláh, G.; Rios, E.C.S.; Yanagi, K.; Ahmad, A.; Papapetropoulos, A.; Szabo, C. Regulation of Vascular Tone, Angiogenesis and Cellular Bioenergetics by the 3-Mercaptopyruvate Sulfurtransferase/H2S Pathway: Functional Impairment by Hyperglycemia and Restoration by DL-alpha-Lipoic Acid. Mol. Med. 2015, 21, 1–14. [Google Scholar] [CrossRef]
- Rennie, W.; Kanoria, S.; Liu, C.; Mallick, B.; Long, D.; Wolenc, A.; Carmack, C.S.; Lu, J.; Ding, Y. STarMirDB: A database of microRNA binding sites. RNA Biol. 2016, 13, 554–560. [Google Scholar] [CrossRef]
- Barutta, F.; Bellini, S.; Mastrocola, R.; Bruno, G.; Gruden, G. MicroRNA and Microvascular Complications of Diabetes. Int. J. Endocrinol. 2018, 2018, 6890501. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Lv, C.; Li, L.; Chen, S.; Liu, S.; Wang, C.; Su, B. Plasma miR-126 Is a Potential Biomarker for Early Prediction of Type 2 Diabetes Mellitus in Susceptible Individuals. BioMed Res. Int. 2013, 2013, 761617. [Google Scholar] [CrossRef]
- Glaich, O.; Parikh, S.; Bell, R.E.; Mekahel, K.; Donyo, M.; Leader, Y.; Shayevitch, R.; Sheinboim, D.; Yannai, S.; Hollander, D.; et al. DNA methylation directs microRNA biogenesis in mammalian cells. Nat. Commun. 2019, 10, 5657. [Google Scholar] [CrossRef] [PubMed]
- Behera, J.; George, A.K.; Voor, M.J.; Tyagi, S.C.; Tyagi, N. Hydrogen sulfide epigenetically mitigates bone loss through OPG/RANKL regulation during hyperho-mocysteinemia in mice. Bone 2018, 114, 90–108. [Google Scholar] [CrossRef]
- Lamalice, L.; Le Boeuf, F.; Huot, J. Endothelial Cell Migration During Angiogenesis. Circ. Res. 2007, 100, 782–794. [Google Scholar] [CrossRef]
- de Mello, A.H.; Liu, T.; Garofalo, R.P.; Casola, A. Hydrogen Sulfide Donor GYY4137 Rescues NRF2 Activation in Respiratory Syncytial Virus Infection. Antioxidants 2022, 11, 1410. [Google Scholar] [CrossRef]
- Li, S.; Yang, G. Hydrogen Sulfide Maintains Mitochondrial DNA Replication via Demethylation of TFAM. Antioxidants Redox Signal. 2015, 23, 630–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuyama, H.; Suzuki, H.I. Systems and Synthetic microRNA Biology: From Biogenesis to Disease Pathogenesis. Int. J. Mol. Sci. 2019, 21, 132. [Google Scholar] [CrossRef] [PubMed]
- Heuslein, J.L.; Gorick, C.M.; Song, J.; Price, R.J. DNA Methyltransferase 1–Dependent DNA Hypermethylation Constrains Arteriogenesis by Augmenting Shear Stress Set Point. J. Am. Heart Assoc. 2017, 6, e007673. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, W.; Zhang, Q.; Chen, Y.; Zhu, Y. Hydrogen Sulfide Improves Angiogenesis by Regulating the Transcription of pri-miR-126 in Diabetic Endothelial Cells. Cells 2022, 11, 2651. https://doi.org/10.3390/cells11172651
Xue W, Zhang Q, Chen Y, Zhu Y. Hydrogen Sulfide Improves Angiogenesis by Regulating the Transcription of pri-miR-126 in Diabetic Endothelial Cells. Cells. 2022; 11(17):2651. https://doi.org/10.3390/cells11172651
Chicago/Turabian StyleXue, Wenlong, Qingqing Zhang, Ying Chen, and Yichun Zhu. 2022. "Hydrogen Sulfide Improves Angiogenesis by Regulating the Transcription of pri-miR-126 in Diabetic Endothelial Cells" Cells 11, no. 17: 2651. https://doi.org/10.3390/cells11172651