1. Introduction
According to the WHO statistics, 9.9 million people died from cancer in 2020 [
1]. Chemotherapy is the most often used treatment strategy against advanced and disseminated tumors, as an estimated 60% of all cancer patients receive chemotherapeutics [
2]. To control tumor growth, conventional chemotherapy agents target rapidly dividing cancer cells, exploiting vulnerabilities linked to increased DNA synthesis and enhanced use of cell division machinery. Unfortunately, the effects of chemotherapy are often short-lived, as cancer cells develop various mechanisms to cope with continuous drug exposure.
Recently, the tumor microenvironment (TME) attracted increasing attention as a possibly important contributor to drug resistance. TME consists of immune cells, vascular endothelial cells, and mesenchymal stem cells (MSCs) or fibroblasts, which can have a critical role in tumor progression, metastasis, and drug resistance. MSCs are multipotent cells, with self-renewable capacity, immunosuppressive effect, tissue regeneration, and high migration trait. These cells can be isolated from different sources, such as adipose tissue, bone-marrow, umbilical cord, dental pulp, or dermal tissue. MSCs have the potential to migrate to the primary tumor site, mostly from the local adipose tissue and the distant bone marrow niche due to endocrine and paracrine signals (bFGF, HDGF, MCP-2, UPA) secreted by the cancer cells [
3]. These MSCs can form a major subpopulation in the tumor stroma, for example, in breast- and pancreatic carcinoma, the MSCs ratio can be nearly 80% of the whole tumor mass [
4]. Because of the complexity and cellular heterogeneity of the TME, there is an intensive crosstalk between the different cell types [
5].
MSCs, as one of the most abundant cell types in TME [
6], are known to affect neighboring cells directly or indirectly. Through releasing a complete arsenal of different cytokines, chemokines, and matrix metalloproteinases (MMP1, MMP2), MSCs can reorganize the extracellular matrix or by secreting interleukins (e.g., IL-6) can alter the behavior and the gene expression profile of other cells. These multipotent cells can have a direct and dynamic interaction with cancer cells via membrane- and microvesicules/exosomes exchange, cell fusion, or by transferring different proteins and mRNAs [
7].
MSCs can have a somewhat controversial dual role during tumor progression: these cells can exert either pro-tumorigenic or antitumoral effects depending on the phase of the process. Their primary anti-cancer activity—expressing antiangiogenic agents (PDGF), cytotoxic agents (TRAIL, INF-γ), anti-proliferation factors (Wnt, BMP, PTEN), and immunomodulators targeting NK-cells, B-cells, and T-cells. Later, pro-tumorigenic activity will prevail by secreting survival factors (bFGF, HGF, IGF1), pro-angiogenic agents (VEGF, TGFB), immunosuppressive cytokines (IDO, TGFB, IL8, IL10), and transform to cancer-associated fibroblast (CAF) [
7].
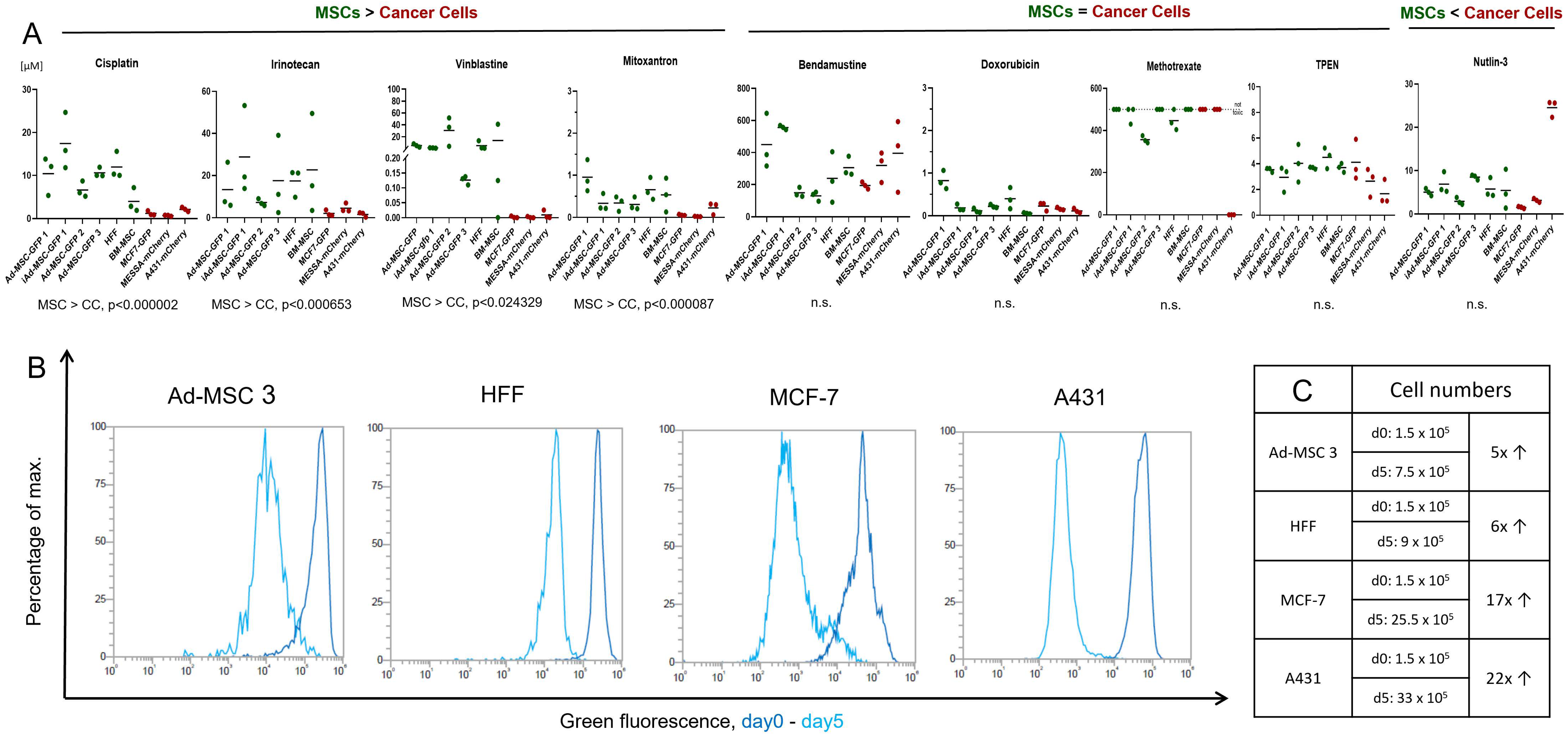
According to the widely accepted view, chemotherapeutics causes less damage to healthy/non-proliferating/non-tumor cells, targeting only the highly proliferative cancer cells. Our initial aim was to test nine, structurally and mechanistically different, clinically applied anticancer agents in vitro both on MSCs and cancer cells to identify cellular mechanisms helping healthy cells survive under various chemotherapy treatment/conditions. Surprisingly, in our drug sensitivity screening, we found that, while there are some compounds that kill cancer cells significantly more efficiently, 5 out of 9 molecules were equally toxic to both cancer cells and MSCs. These striking results challenge the basics of conventional chemotherapy, shed light on the complex interplay between cancer cells and the TME, and could lead to novel approaches in therapy design.
Here, we compare the response of cancer cells and MSCs isolated from healthy donors to a panel of chemotherapeutic agents, investigate the effect of these treatments on DNA damage, reactive oxygen species production, apoptosis, and cellular senescence, while we try to dissect the differences in the reaction to these treatments.
2. Materials and Methods
2.1. Cell Culture
To compare drug sensitivity, a panel of six MSCs and three cancer cell lines were used.
Three human primary adipose tissue-derived mesenchymal stem cells, Ad-MSC 1-3, an immortalized Ad-MSC 1 (iAd-MSC 1), originating from the Ad-MSC 1 primary cell line with lentiviral mediated overexpression of polycomb complex protein Bmi-1, and human telomerase reverse transcriptase (hTERT) according to Tátrai et al. (2012) were used [
8]. Bone-marrow-derived mesenchymal stem cell (BM-MSC) was isolated and characterized by our group, according to Szepesi et al. (2016). Detailed characterization of the BM-MSC cell line is shown in
Supplementary Figure S1. Human foreskin fibroblast (HFF), human skin epidermoid carcinoma (A431), and human uterine sarcoma (MES-SA) cells were purchased from ATCC, and the breast adenocarcinoma (MCF-7) cell line was obtained from the National Cancer Institute’s Developmental Therapeutics Program. Cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM-F12, Gibco, Waltham, Massachusetts) supplemented with 10% fetal bovine serum (FBS, Gibco), 1% L-glutamine (Gibco), 0.1% gentamicin (Gibco, 50 mg/mL), and 16 ng/mL fibroblast growth factor 2 (Peprotech, London, UK). Cells were kept at 37 °C and 5% CO
2. At approximately 80% confluency, cells were trypsinized (0.1% for MSCs and 0.2% for cancer cells) for between 2 and 10 min. Followed by centrifugation (1500 rpm/5 min), resuspension, and replating for expansion. The medium was renewed every 72 h on each cell line and MSCs were used up to passage 25. GFP or mCherry fluorescent protein expressing sublines were previously established with lentiviral transduction by our group: Ad-MSCs-GFP, immortalized Ad-MSC-GFP 1, A431-mCh, MES-SA-mCh, and MCF-7-GFP according to Tátrai et al. [
9]. Drug sensitivity of Ad-MSC-GFP 3 and A431-mCherry is similar to their parental cell lines (
Supplementary Figure S2).
2.2. Drugs and Chemicals
Chemotherapeutic agents were purchased from different sources: cisplatin (Vnr461201, Accord Healthcare, UK), methotrexate (M9929, Sigma Aldrich, St. Louis, MO, USA), irinotecan hydrochloride (AB252173, abcr, Karlshure, Germany), doxorubicin hydrochloride (25316-40-9, Sigma Aldrich, United States), vinblastine (HY-13780, MedChemExpress, Monmouth Junction, NJ, USA), TPEN (P4413, Tocris, Bristol, UK), nutlin-3 (3984, Tocris, UK), mitoxantrone (M2305000, Sigma Aldrich, UK), and bendamustine was a kind gift from Servier.
2.3. Cytotoxicity Assay and Cell Viability Assay
Cells were seeded in a 96-well tissue culture plate in 100 µL of medium. Ad-MSC-GFP 1, iAd-MSC-GFP 1, Ad-MSC-GFP 2, Ad-MSC-GFP 3, BM-MSC, and HFF were seeded at 5 × 103/100 µL/well density and A431-mCh, MES-SA-mCh, and MCF-7-GFP at 4 × 103 cells/100 µL/well concentration in DMEM-F12 media. Different initial cell numbers were used due to different proliferation rates of different cell types. Cells were allowed to attach overnight at 37 °C and the day after were treated with the drugs using the indicated concentrations in 100 µL. Cell viability was measured by using PrestoBlue™ Cell Viability Reagent (A13232, Invitrogen, Waltham, MA, USA). To address metabolic differences between cancer cells and MSCs, the assay was optimized for each cell line, cells were incubated with 5 or 12.5% PrestoBlue™ Cell Viability Reagent diluted in PBS at 37 °C/5% CO2 for 1–1.5 h 5 days after the treatment. The fluorescence signal was measured by an EnSpire (Perkin Elmer, Waltham, MA, USA) fluorometer at 555 nm (excitation)/585 nm (emission) wavelength.
2.4. Dose–Response Curves and IC50 Calculation
After the incubation period, the relative fluorescence data was measured. The data was normalized with the fluorescence of blank samples (cell culture media). A sigmoidal dose–response curve was fitted to the normalized measurement data and the logarithm of the concentration using the least square method. An inhibitory concentration of 50% (IC50 and log10) was interpolated from the fitted graph. In the case of co-culture, we experienced that at low concentrations the data did not fit the sigmoidal curve. In these cases, IC50 was calculated from the line connecting the measurement points. For repeated measurements, average and standard deviation was calculated based on the logarithm of the IC50 values.
2.5. Apoptosis Assay
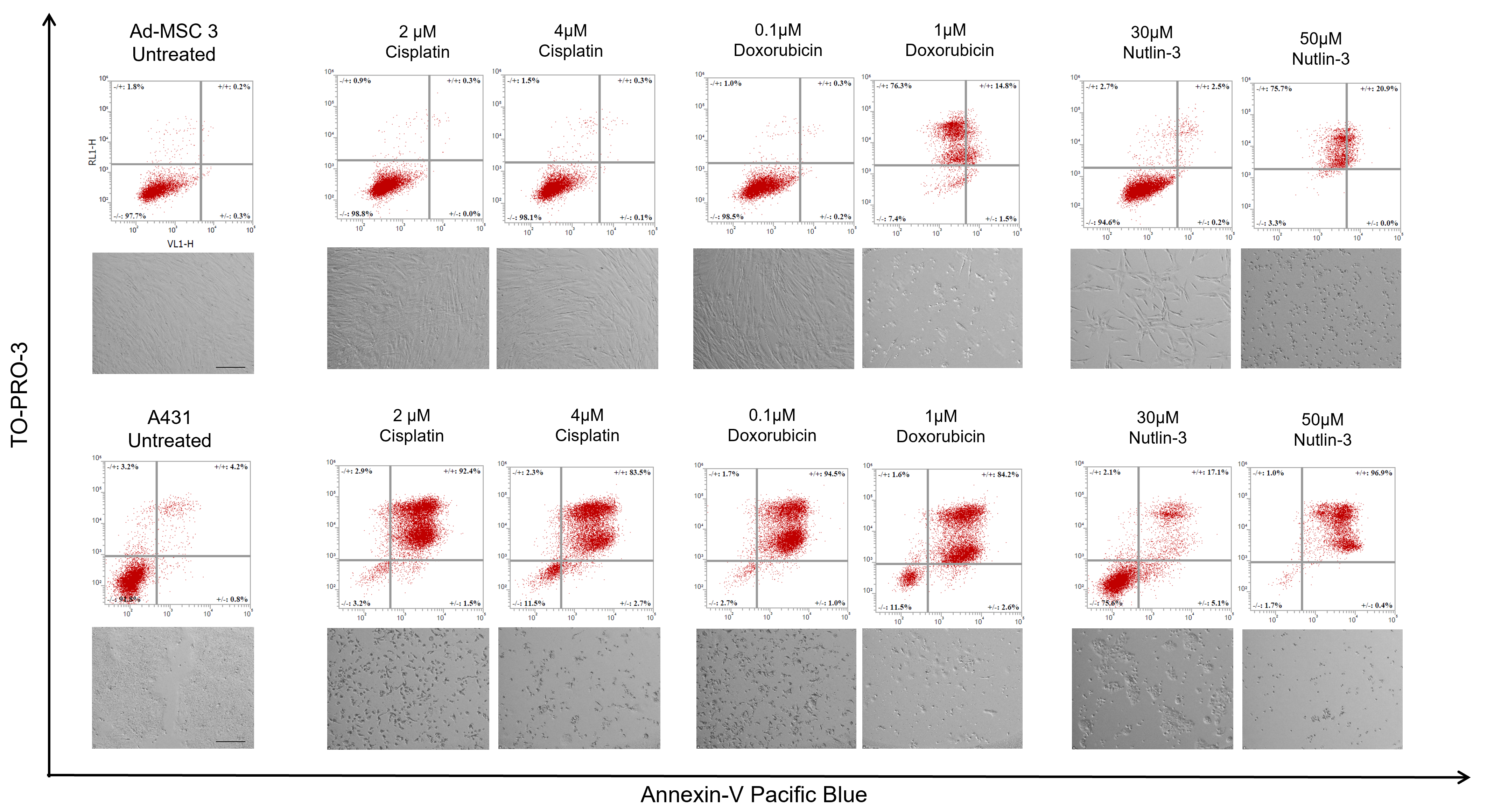
Apoptosis detection was based on the Annexin-V molecule binding to phosphatidylserine membrane molecules. Two cell lines were used to study apoptosis: Ad-MSC 3 cells were plated in a 5 × 104 cells/well concentration, while 4 × 104/well A431-mCh were seeded into each well in a 12-well tissue culture plate. Cells were incubated overnight, and treated with cisplatin, doxorubicin, and nutlin-3 at the indicated concentrations in 1 mL of culture media. At day 5, cells were trypsinized and centrifuged, while the supernatant was also collected and combined with the detached cells to include all already dead cells. Cells were washed twice with 500 µL of 1% BSA-PBS (bovine serum albumin dissolved in PBS, A8022, Sigma-Aldrich, St. Louis, MO, USA) solution and centrifuged at 400× G for 4 min. Cells were resuspended in 100 µL of 1× Annexin Binding Buffer (422201, Biolegend, San Diego, CA, USA), and 3 µL of Annexin V-Pacific Blue (A35122, Biolegend, San Diego, CA, USA) and 1 µL of TO-PRO-3 (R37170, Invitrogen, Waltham, MA, USA) were added to each sample. After 15 min of incubation, labeled and non-labeled samples were diluted 5-fold with 1× Annexin Binding Buffer and analyzed by Attune™ NxT flow cytometer.
2.6. Cell Proliferation Analysis
Cell division was analyzed by the fluorescent CytoTell Green dye (AAT Bioquest, Sunnyvale, CA, USA) according to the manufacturer’s guidelines. Briefly, 3 × 105 cells were centrifuged and stained with 2 µL of CytoTell dye diluted in 1 mL of serum-free DMEM and incubated at 37 °C for 30 min. After centrifugation, half of the cells (1.5 × 105) were plated into T25 cell culture flasks in 5 mL of media and analyzed at day 5. Day 1 samples were co-stained with Zombie Violet dye (423113, Biolegend) to exclude dead cells and analyzed by Attune™ NxT flow cytometry.
2.7. Immunocytochemistry
Ad-MSC-GFP 3 cells were seeded into glass-bottomed chamber slides (ibidi, Germany, Grafelfing, Germany) at 1 × 104/well and A431-mCh at 1.6 × 104/well density. The day after, cells were treated with 2 µM of cisplatin, 0.1 µM of doxorubicin, and 30 µM of nutlin-3 diluted in 200 µL of DMEM-F12. Cells in all chambers were fixed with 4% PFA for 15 min and washed twice with PBS. Blocking solution (0.5% BSA-PBS, 0.1% TritonX-100, 5% goat serum, and 1% fish gelatin) was used for 1 h. γ-H2A.X primary antibody (MA1-2022, Invitrogen, United States) was diluted in blocking solution at 1:500 concentration and incubated overnight at 4 °C. Afterwards, the cells were washed with PBS twice and goat anti-mouse A633 secondary antibody (A21052, abcam, Cambridge, UK) was diluted at 1:250 concentration in blocking solution. Nuclei was stained with a Hoechst-33342 (23491-52-3, Dojindo, Kumamoto, Japan) at 1:1000 concentration. Cells were observed with a Zeiss LSM-710 confocal microscopy at 40× magnification with the same settings for all samples.
2.8. γ-H2AX Image Analysis
To quantify DNA damage based on immunofluorescence against γ-H2AX, a semi-automated image analysis pipeline was set up using CellProfiler v4.1.3 [
10]. First, images showing DAPI-stained nuclei were pre-processed by intensity normalization and Gaussian filtering (ϭ = 20 px) to suppress noise and remove artifacts caused by debris. Nuclei candidates were identified by threshold-based segmentation; false candidates were removed from the analysis manually. Using the remaining nuclei regions as masks, pixel intensities were measured on the γ-H2AX channel, thus, excluding non-nuclei signals.
2.9. ROS Induction
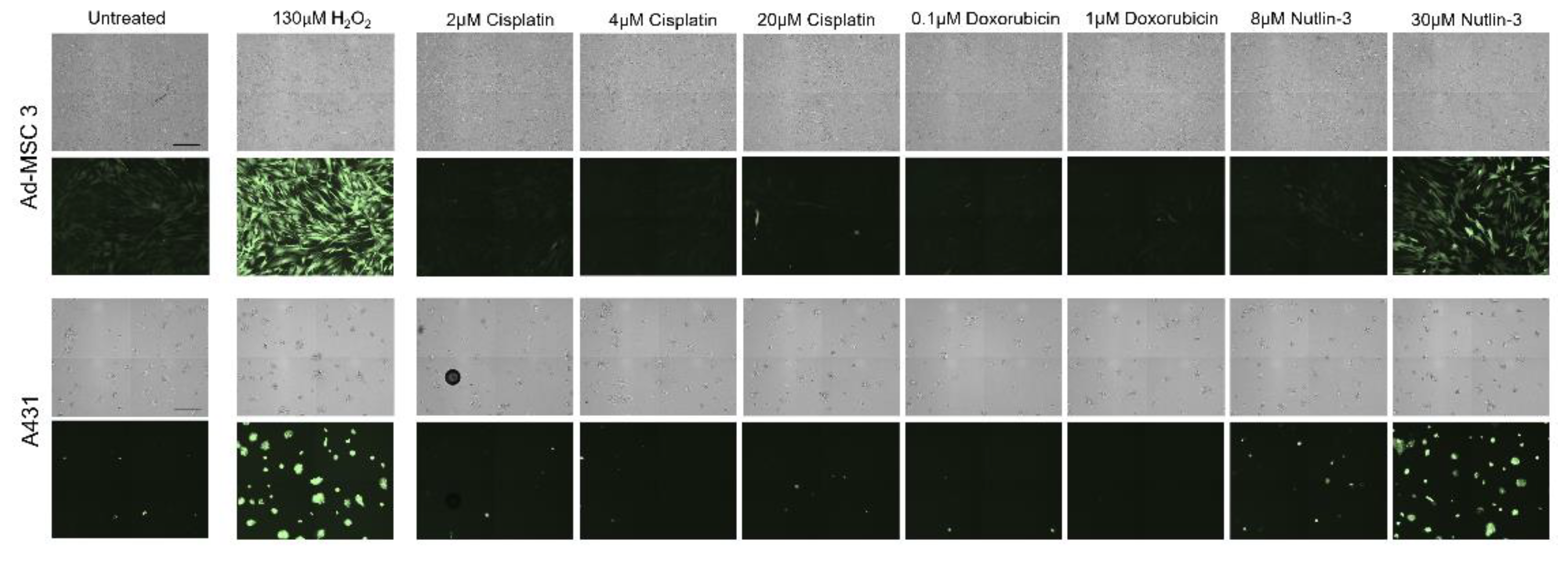
Ad-MSC 3 and A431 cells were seeded in a 96-well tissue culture plate and incubated for 24 h as described above. Cells were washed twice with HBSS and incubated in 100 µL of DCFH-DA dye (R252-10, Dojindo) according to the manufacturer’s instructions. Cells were incubated for 30 min at 37 °C and washed twice with HBSS. Cells were treated with 200 µL of cell culture medium with appropriate concentrations of H2O2, cisplatin, doxorubicin, and nutlin-3 for 1 h. Supernatant was removed, and the cells were washed with HBSS before imaging with the JuLITM Stage Cell History Recorder (NanoEnTek).
2.10. Senescence Staining
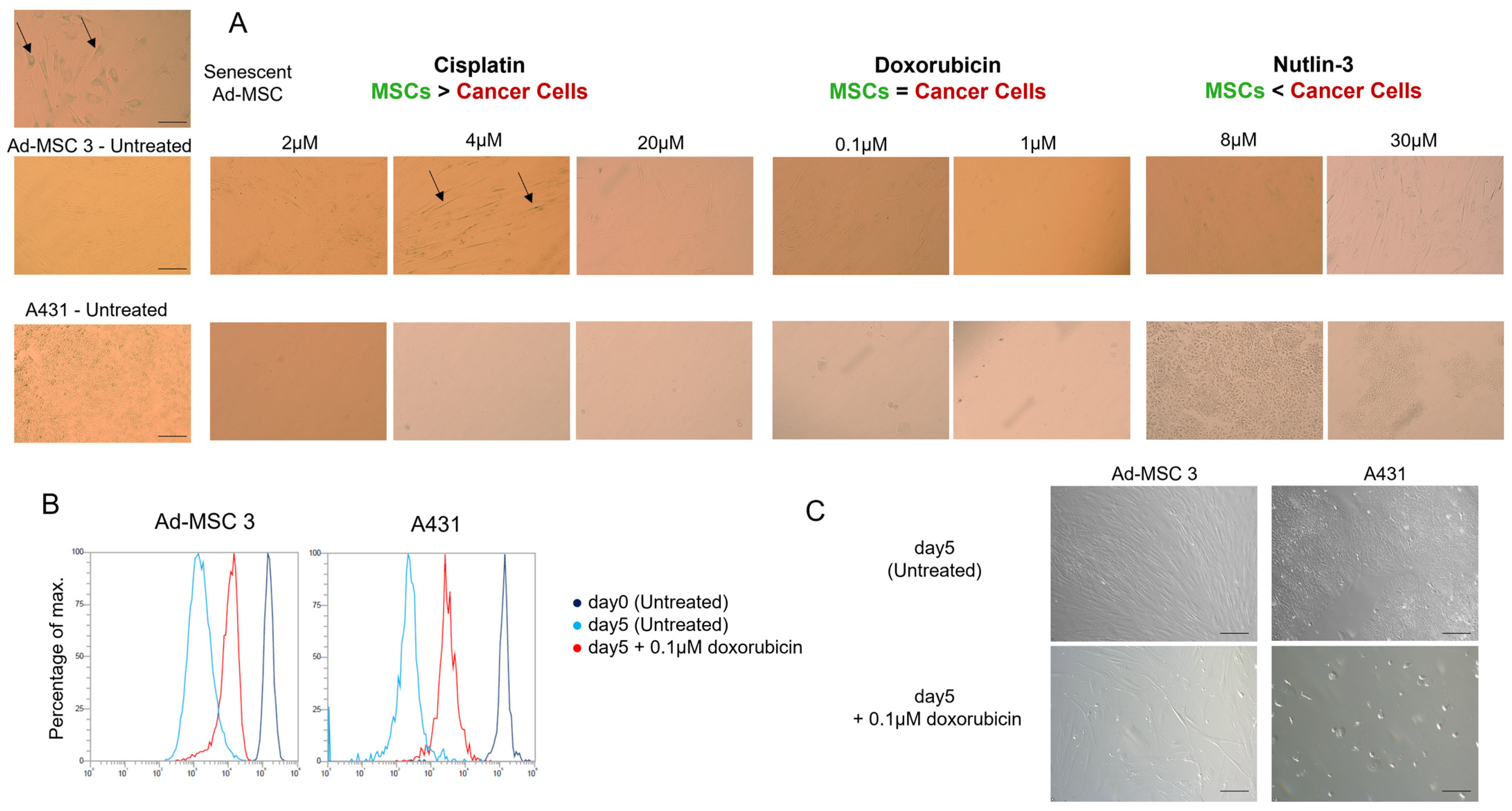
Ad-MSC 3 and A431 cells were seeded in a 96-well tissue culture plate and treated with cisplatin, doxorubicin, or nutlin-3. At day 5, media was removed and replaced with 200 µL of completed DMEM-F12 for 7 days. The Senescence β-Galactosidase Staining Kit (9860S, Cell Signaling Technology, Danvers, MA, USA) was used to detect senescent cells. Cells were fixed with fixing solution, washed with PBS, stained with X-gal solution overnight at 37 °C without CO2, and coated with 70% glycerol.
2.11. Live Cell Imaging
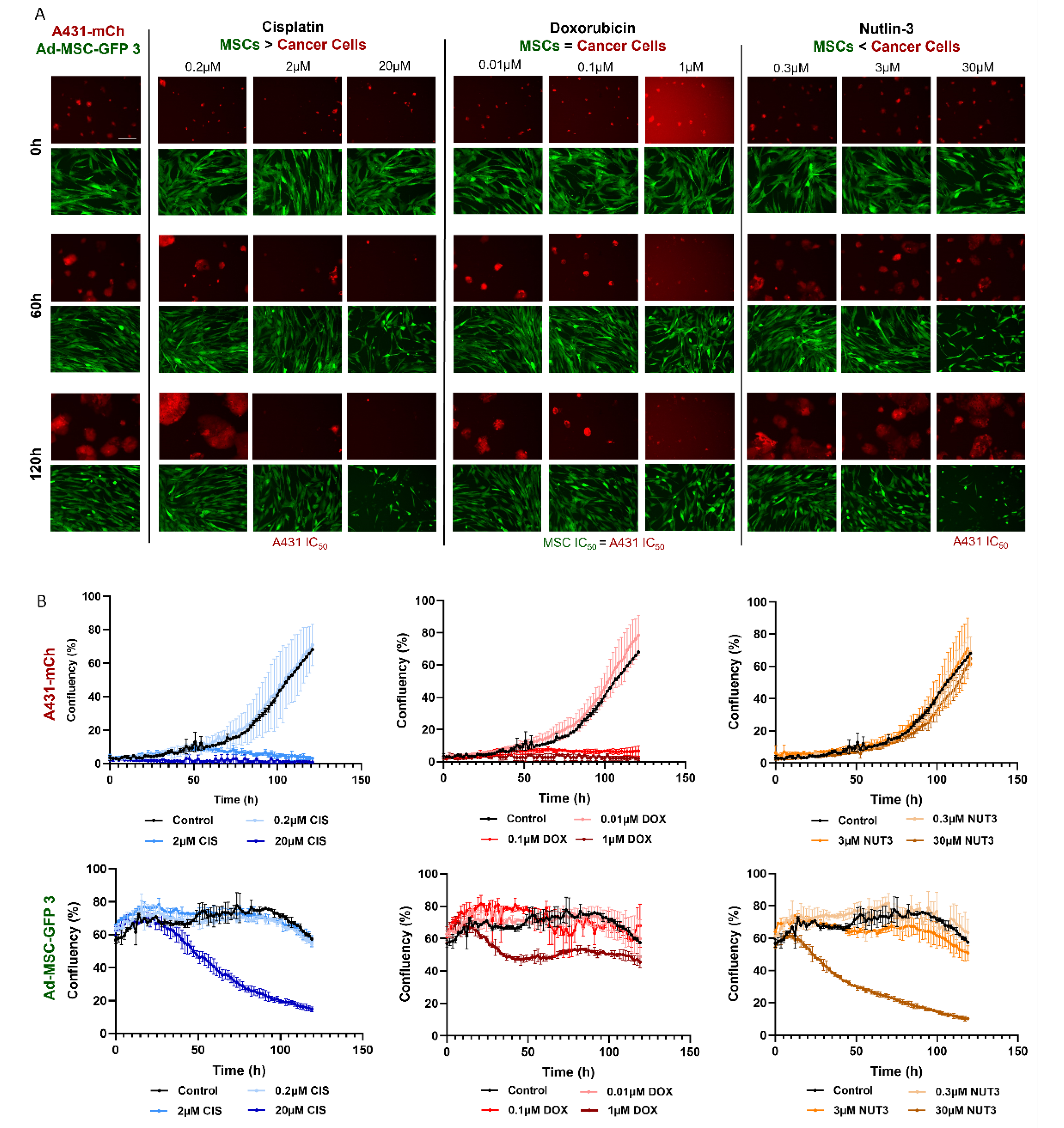
JuLI™ Stage Cell History Recorder was used to monitor a five-day cytotoxicity assay on Ad-MSC-GFP 3 and A431-mCh cell lines. Cells were seeded and treated with cisplatin, doxorubicin, and nutlin-3 as described above. Images were taken every 105 min for 120 h at 10× objective magnification. Live cell microscopy studies were evaluated using the “Growth curve” function of the JuLI STAT image analysis software (v2.0.1.0).
Brightfield images were taken with the Olympus IX51 microscope. The scale bar is the same in every image.
2.12. Statistical Analysis
To compare the mean IC50 values of six mesenchymal stem cell lines (Group-1) and three cancer cell lines (Group-2), first an F-test was performed to check equal variances between the groups, then two-sample unpaired t-tests were performed to calculate significance using Microsoft® Excel® 2016 (16.0.4266.1001).
4. Discussion
Although MSCs are one of the most abundant cell types in the TME, their response to the applied chemotherapies is mostly unknown and a systematic drug screen on MSCs has never been performed before [
11]. In this work, we compared the drug sensitivity of 6 MSC lines isolated from cancer-free individuals and 3 widely used cancer cell lines to 9 structurally and mechanistically different compounds and characterized their response to 3 of these in detail.
In our screening, MSCs showed strong resistance, sometimes 10,000-fold, to DNA intercalation (cisplatin), topoisomerase I and II inhibition (irinotecan, mitoxantrone), and microtubule disturbances (vinblastine). Similar results were observed previously by others testing the same compounds on MSCs. Nicolay et al. showed bone marrow (BM)-derived MSCs and differentiated fibroblasts exhibit resistance to irinotecan and etoposide, most likely due to efficient DNA repair [
12]. Both Mueller and Liang et al. found that BM- and adipose tissue-derived MSCs pose resistance to cisplatin, vincristine, and camptothecin, and rapidly recover after drug exposure [
13,
14]. It is common knowledge that chemotherapy targets rapidly dividing cancer cells, therefore, healthy tissues are affected significantly less than tumors. The widely accepted explanation is that quiescent and slow cycling normal cells have less active DNA synthesis and other proliferation-related activities, which are usually targeted by chemotherapeutics [
15,
16]. Considering MSCs’ significantly lower proliferation rate, the observed resistance can be explained.
Despite MSCs being considered as chemotherapy-resistant in the related literature, our experiments brought striking results that challenge this view. Four chemotherapeutic compounds proved to be surprisingly effective against all cell lines with MSCs having no notable advantage over cancer cells. Intrastrand crosslinking (bendamustine), topoisomerase II inhibition (doxorubicin), ROS-damage (TPEN), and antimetabolite-induced DNA damage (methotrexate) affected MSCs similarly to cancer cells. No direct testing of bendamustine on MSCs was reported previously, however, available results are conflicting and suggest that using the drug can have a negative effect on stem cell mobilization in patients; however, simultaneously, low toxicity was observed on stem cell cultures in vitro [
17,
18]. Doxorubicin and mitoxantrone were found to be slightly toxic to MSCs in some studies, but the main differences were primarily in the expression of a few certain genes (e.g., connexin 43, alkaline phosphatase, troponin T) rather than complete phenotypic changes or cell death. MSCs isolated from doxorubicin-treated rats showed activated DNA damage response pathway, S-phase arrest, and increased sub-G1 subpopulation, while mitoxantrone caused premature senescence in dental pulp MSCs and dermal fibroblasts [
19,
20]. Importantly, in these works, sensitivity of MSCs was not compared to cancer cells. TPEN is a cell permeable Zn
2+ chelator used to induce oxidative stress through the dysregulation of ROS detoxification [
21,
22]. MSCs’ sensitivity to ROS is still unclear: some reports suggest that MSCs have low base levels of ROS with a high amount of the antioxidant glutathione, which protects the cells during oxidative attacks [
23]. In contrast, others showed MSCs are more susceptible to ROS-induced damage than differentiated cells and respond to ROS-inducing agents with DNA damage and senescence; moreover, exogenous antioxidants were required to restore oxidative stress resistance [
24,
25]. In our experiments, TPEN treatment was equally toxic to both MSCs and cancer cells, suggesting MSCs have no advantage against ROS-mediated damage. The antimetabolite methotrexate, used to treat cancer and rheumatoid arthritis, interferes with thymidine synthesis through inhibiting dihydrofolate reductase (DHFR). Methotrexate’s effects rely extensively on DHFR expression and structure, mutations in the active site of the enzyme, or lower expression levels render cells resistant to the drug [
26]. In accordance with our findings, MSCs considered relatively resistant to antimetabolites, such as 5-fluorouracil and gemcitabine, keep their cellular characteristics despite treatment probably as a result of increased DNA metabolism and multidrug resistance transporter expression [
27]. The only significant outlier was the A431 cell line, which proved to be hypersensitive to the compound, which could be the result of the cell-specific DHFR status or the fact that A431 is the only p53-mutant in the cell line panel [
28]. Nutlin-3, an effective MDM2-p53 interaction inhibitor, was designed to selectively kill wild-type p53 expressing cells [
29]. In our assay, only A431 proved to be resistant to nutlin-3, the only cell line with mutated p53 expression.
Finding similar drug sensitivities in both MSCs and cancer cells is even more surprising if we consider that, despite the comparable responses seen in the cytotoxicity assays, the two cell types react quite differently to the same toxic insults. Doxorubicin was equally toxic to MSCs and cancer cells in the cytotoxicity assays, however, in MSCs there were less DNA double-stranded breaks (DSBs), suggesting an increased DNA repair, and while MSCs are known to successfully repair DSBs after treatment with drugs such as bleomycin [
30], it still was not enough to protect cells from the cytotoxic effects of doxorubicin. Furthermore, nutlin-3 caused DNA damage only in cancer cells, yet only killed p53-wt expressing MSCs, which indicates that the amount of DNA DSBs cannot properly predict survival after drug treatment.
Treatment-induced ROS generation was also assessed, but, surprisingly, ROS levels increased significantly only by nutlin-3 in both cancer cells and MSCs. While others hypothesized that nutlin-3 induces ROS through increasing p53 and mitochondrial translocation levels, which in turn result in elevated ROS [
31], in our assay, both p53-wt (MSC) and -mutant (A431) cells showed a robust increase, but, again, only MSCs were killed off after treatment.
The most striking result was, unquestionably, the inability of MSCs to die by apoptosis. There are some findings by others suggesting MSCs have an elevated apoptotic threshold [
13], but the extent of this phenomenon was never shown before. Some annexin-positive, late apoptotic cells were detected in MSC cultures after treatment with high concentrations of doxorubicin and nutlin-3 (1 µM and 50 µM, respectively). However, no distinct apoptotic populations were found, showing that treatments causing massive apoptosis in cancer cells only modestly activate the apoptotic pathways in MSCs. Nevertheless, the same high concentrations of doxorubicin and nutlin-3 caused necrotic cell death in MSCs, suggesting that mesenchymal stem cells can be killed with chemotherapy, but with significantly higher doses and activating different cell death mechanisms. Surprisingly, when the cisplatin concentration was raised to the IC50 level of Ad-MSC-GFP 3 cells (10 µM), notable amounts of late apoptosic cells were observed, suggesting that apoptosis in mesenchymal stem cells can be induced with specific DNA damage insults. We were not able to induce apoptosis in MSCs using several concentrations of mitoxantrone, staurosporine (multikinase inhibitor), H
2O
2 (oxidative stress), and cobalt chloride (hypoxia inducer) in our experiments (data not shown). Other groups reported possible chemotherapy-induced apoptosis in MSCs, however, these works mostly show only a slight increase in cleaved caspase-3 or annexin-V as a marker of apoptotic cells [
12,
20,
27,
32]. This observation could mean MSCs are inherently resistant to apoptosis or more preferably choose other pathways than apoptosis.
To test whether MSCs switch to senescence rather than cell death, the function of senescence-associated ß-galactosidase (SA-ß-GAL) was investigated 12 days after drug treatment, but no significant differences were found. The only alteration found was the decrease in proliferation rate in both cell types. Due to cancer cells being proven to be dying after drug treatment, one possible explanation could be that MSCs slow down their cell cycle progression and buy some time to avoid apoptosis and follow another pathway.
In summary, our results point out important differences in the drug response of MSCs and cancer cells, showing how complex the role of TME could truly be during chemotherapy. These observations could help rational drug design and the development of drug combinations targeting both MSCs and cancer cells in the TME.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





