
The Effects of Cannabidiol on Aqueous Humor Outflow and Trabecular Meshwork Cell Signaling


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Porcine Anterior Segment–Perfused Organ Culture Model
2.2. Porcine Trabecular Meshwork Cell Cultures
2.3. Collagen Gel Contraction Assays
2.4. Western Blot Analysis
2.5. RhoA Activation Assay
2.6. Data Analysis
3. Results
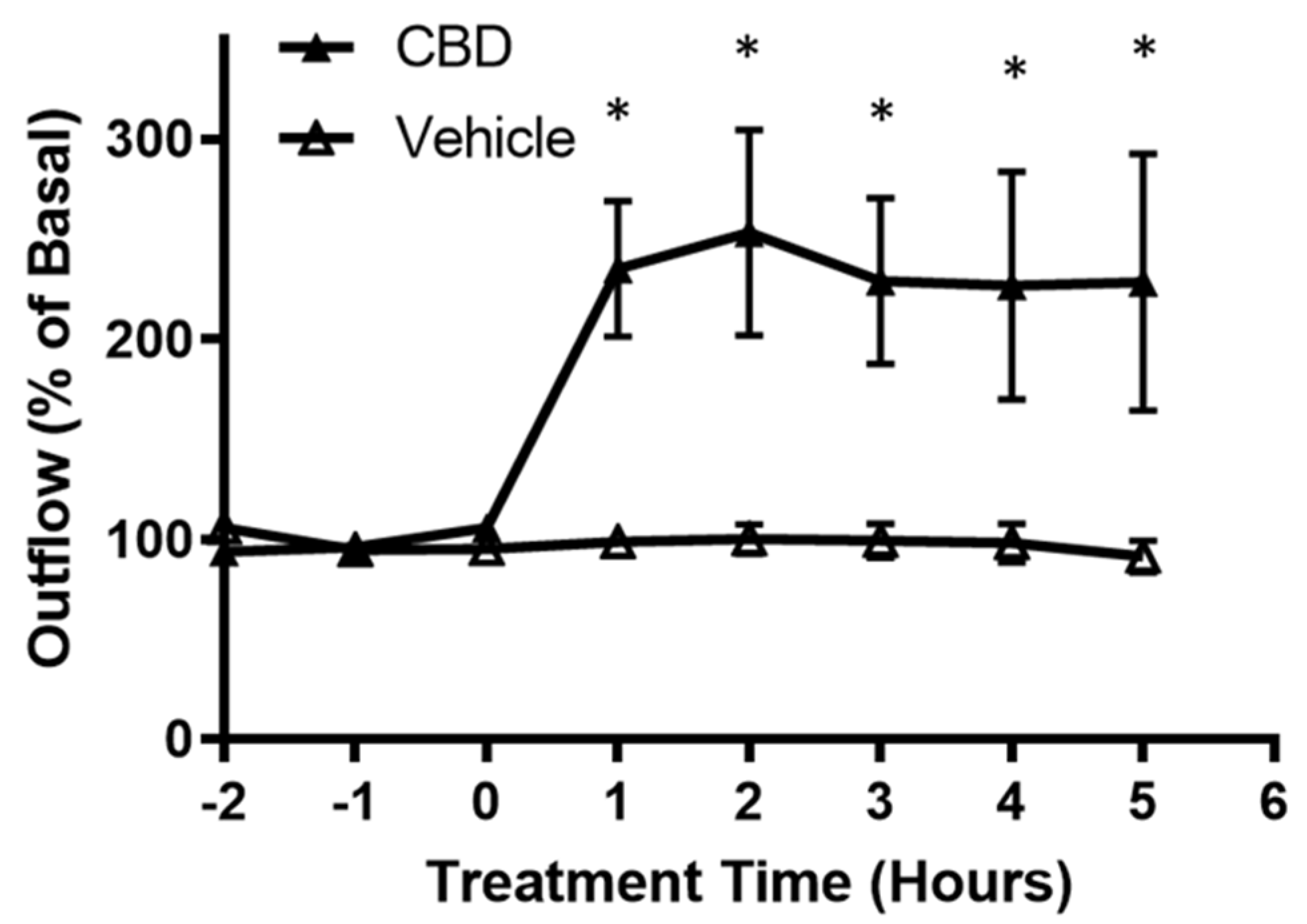
3.1. The Effects of CBD on Aqueous Humor Outflow
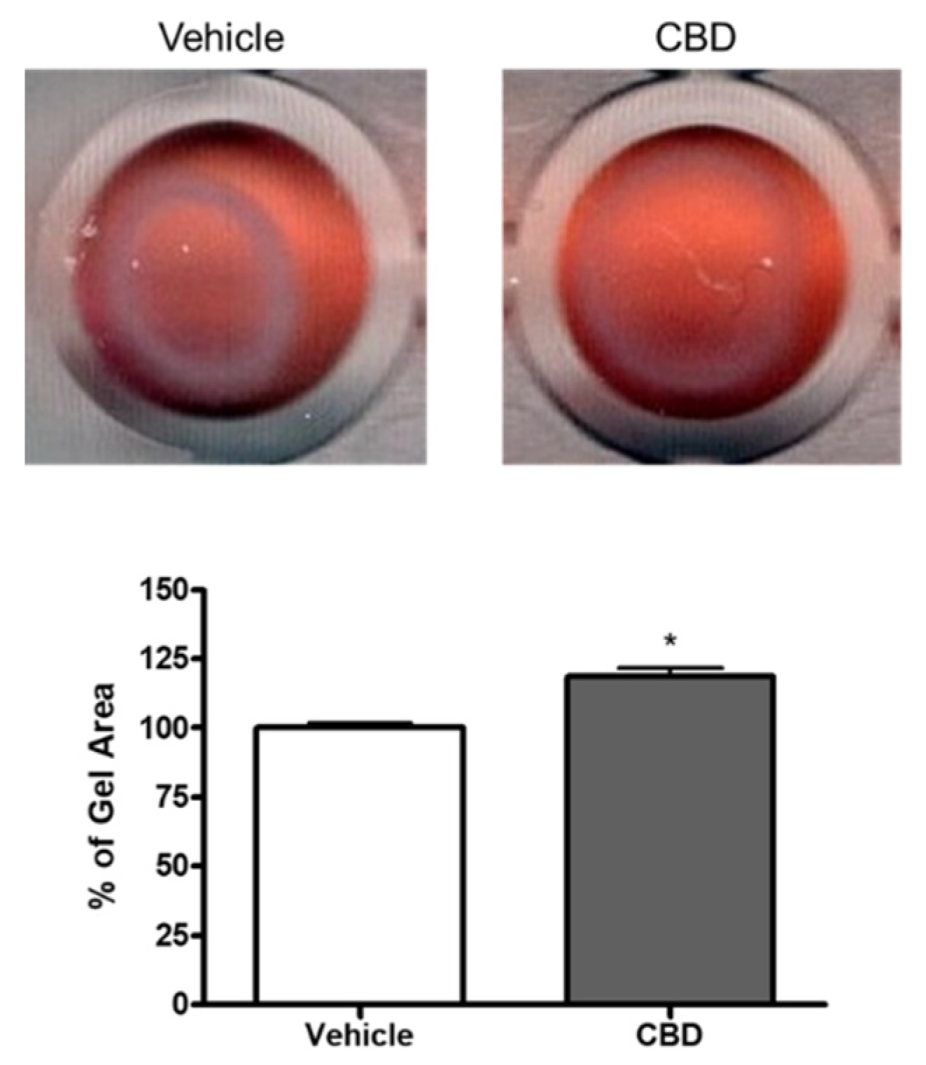
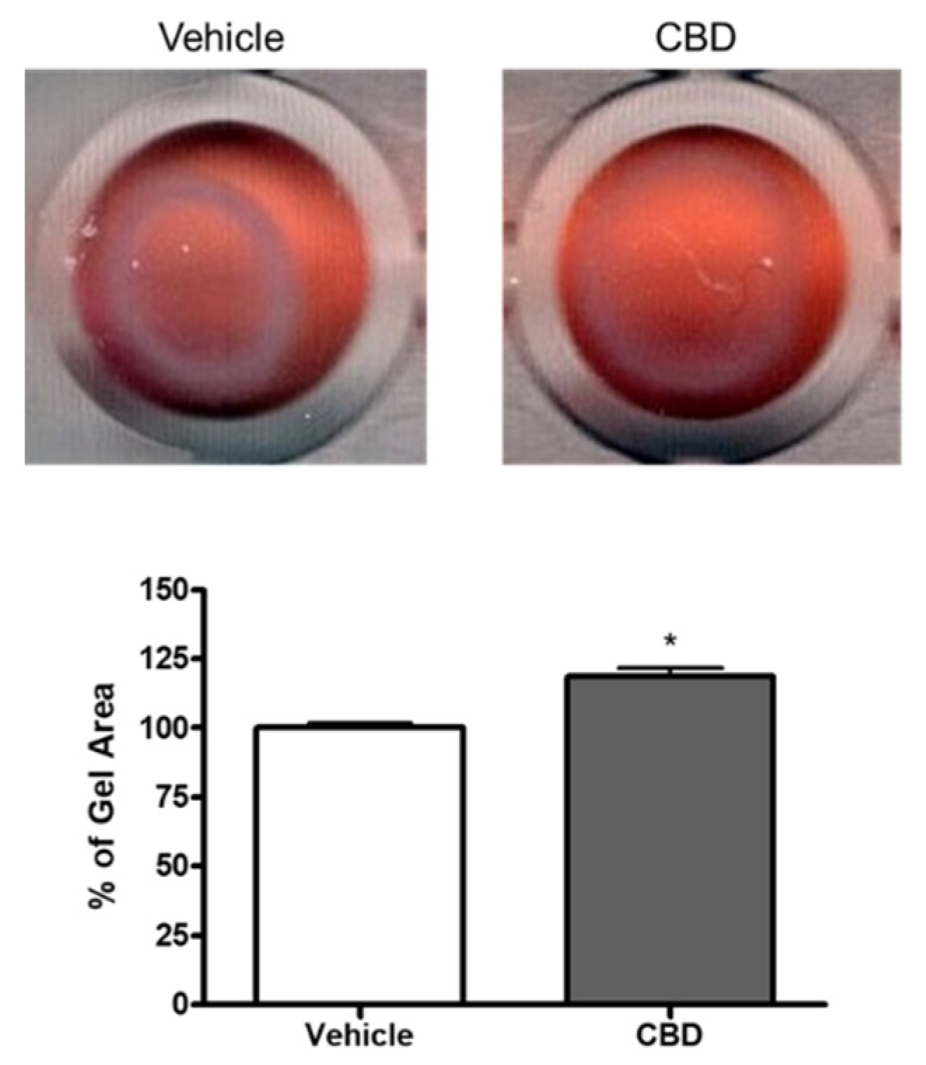
3.2. The Effect of CBD on Collagen Gel Contraction Mediated by TM Cells
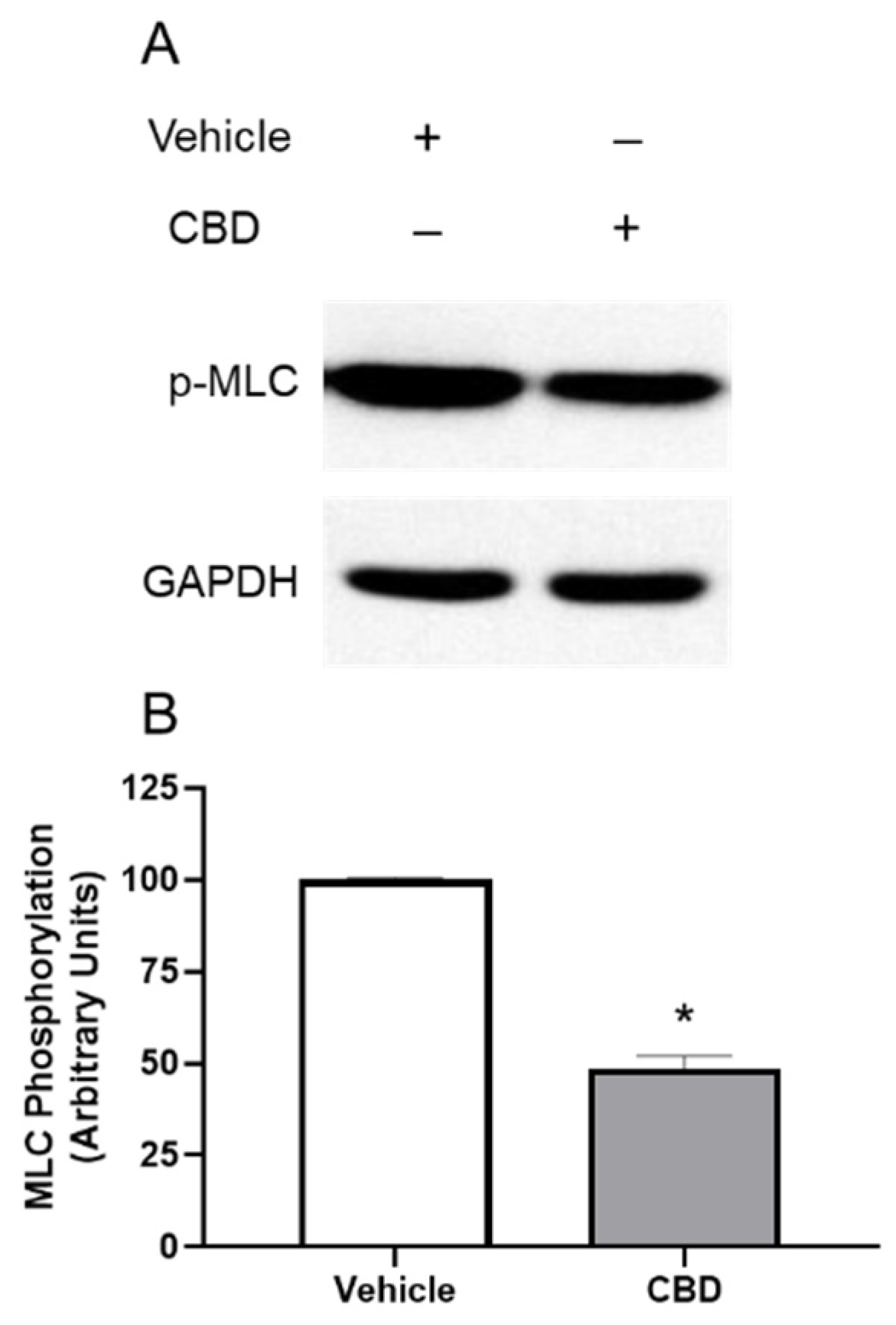
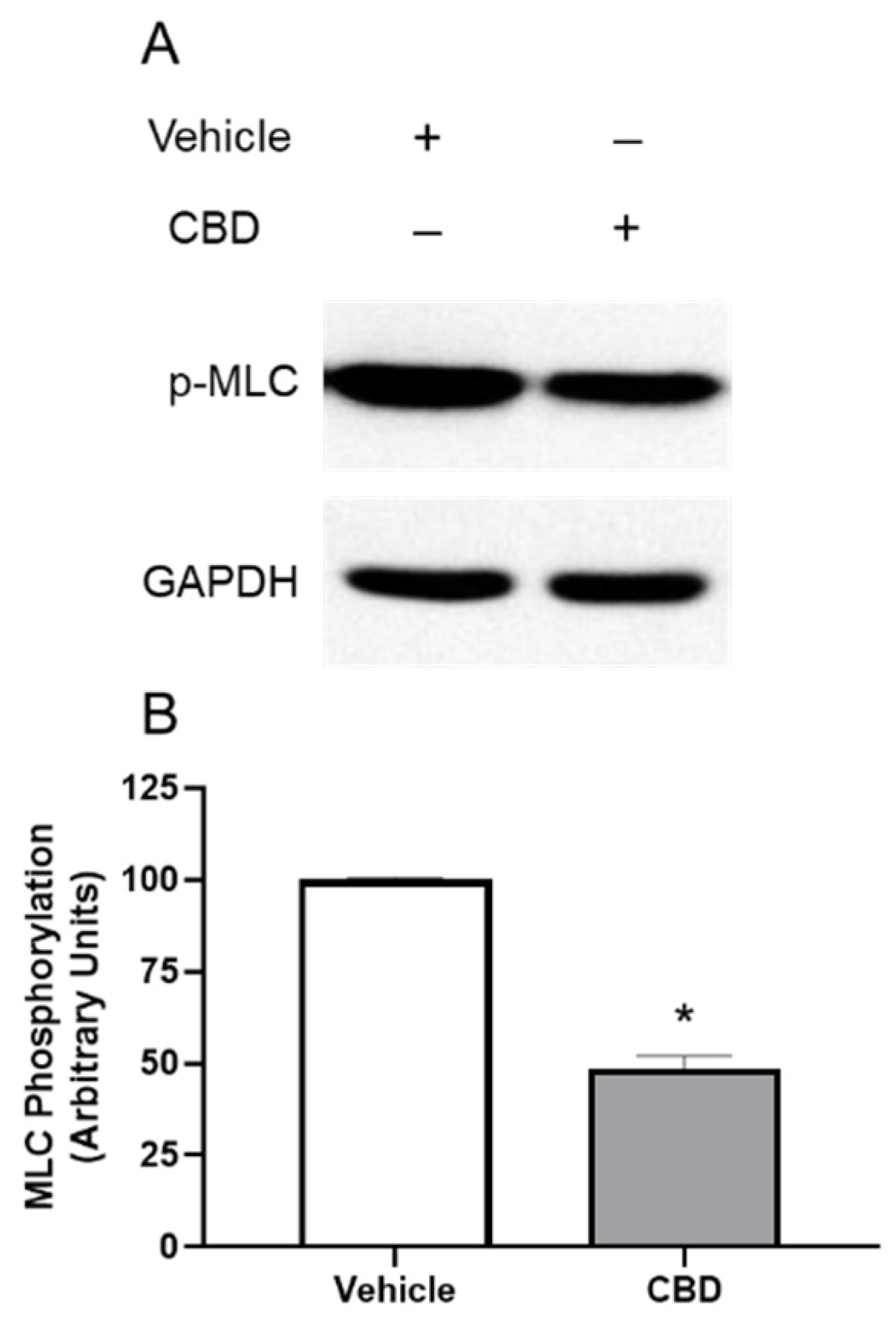
3.3. The Effects of CBD on MLC Phosphorylation in TM Cells
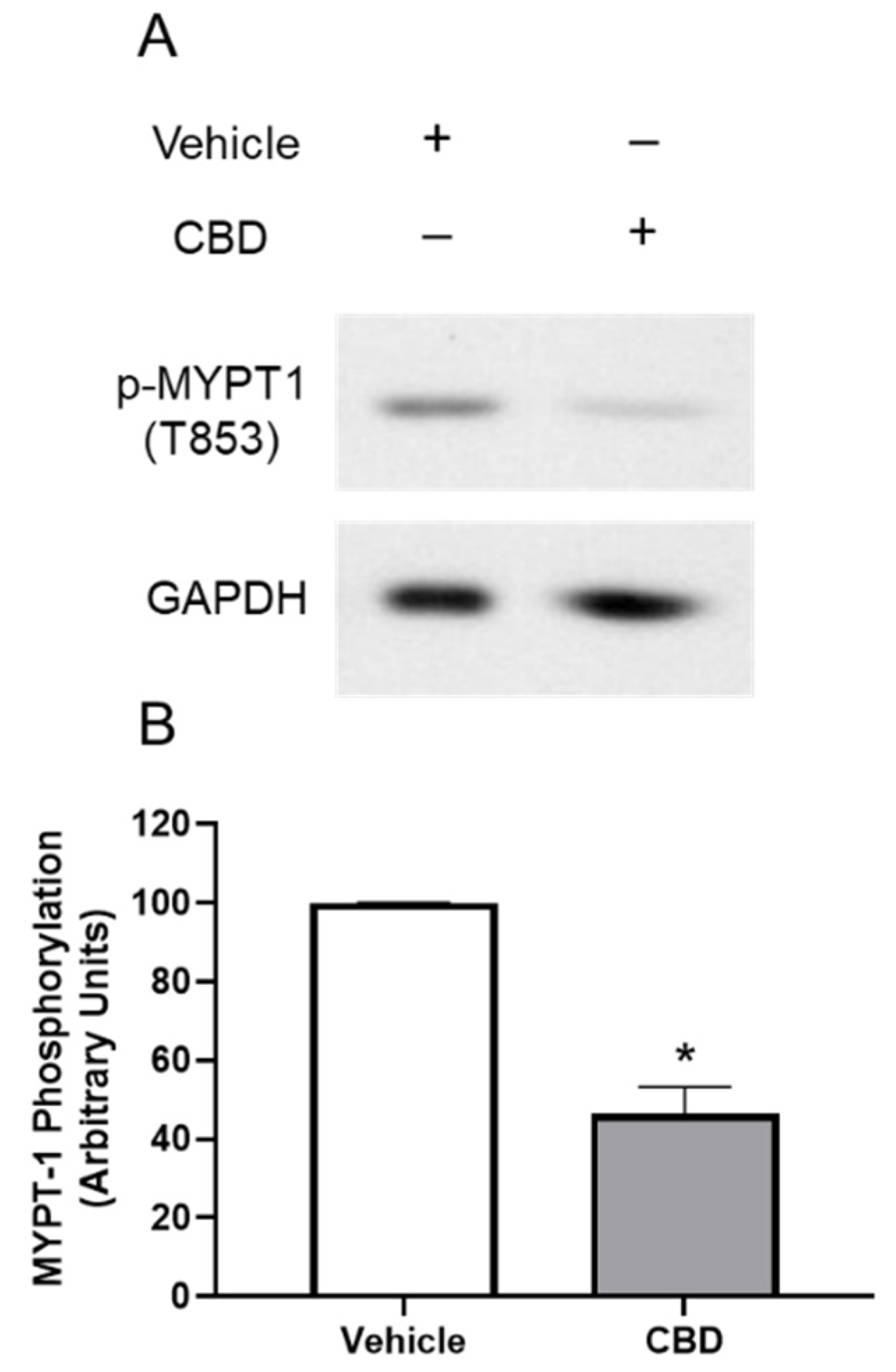
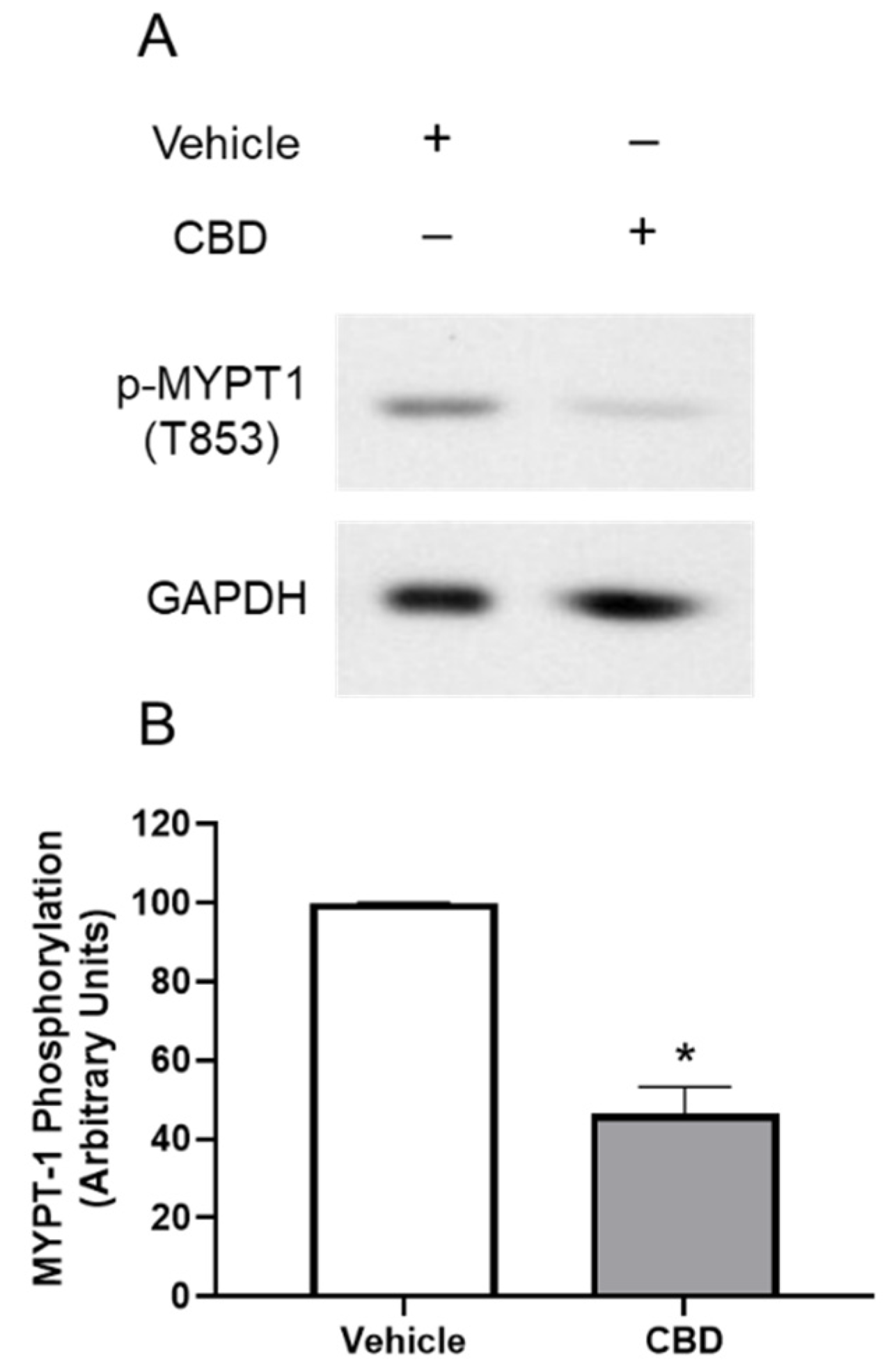
3.4. The Effects of CBD on MYPT1 Phosphorylation in TM Cells
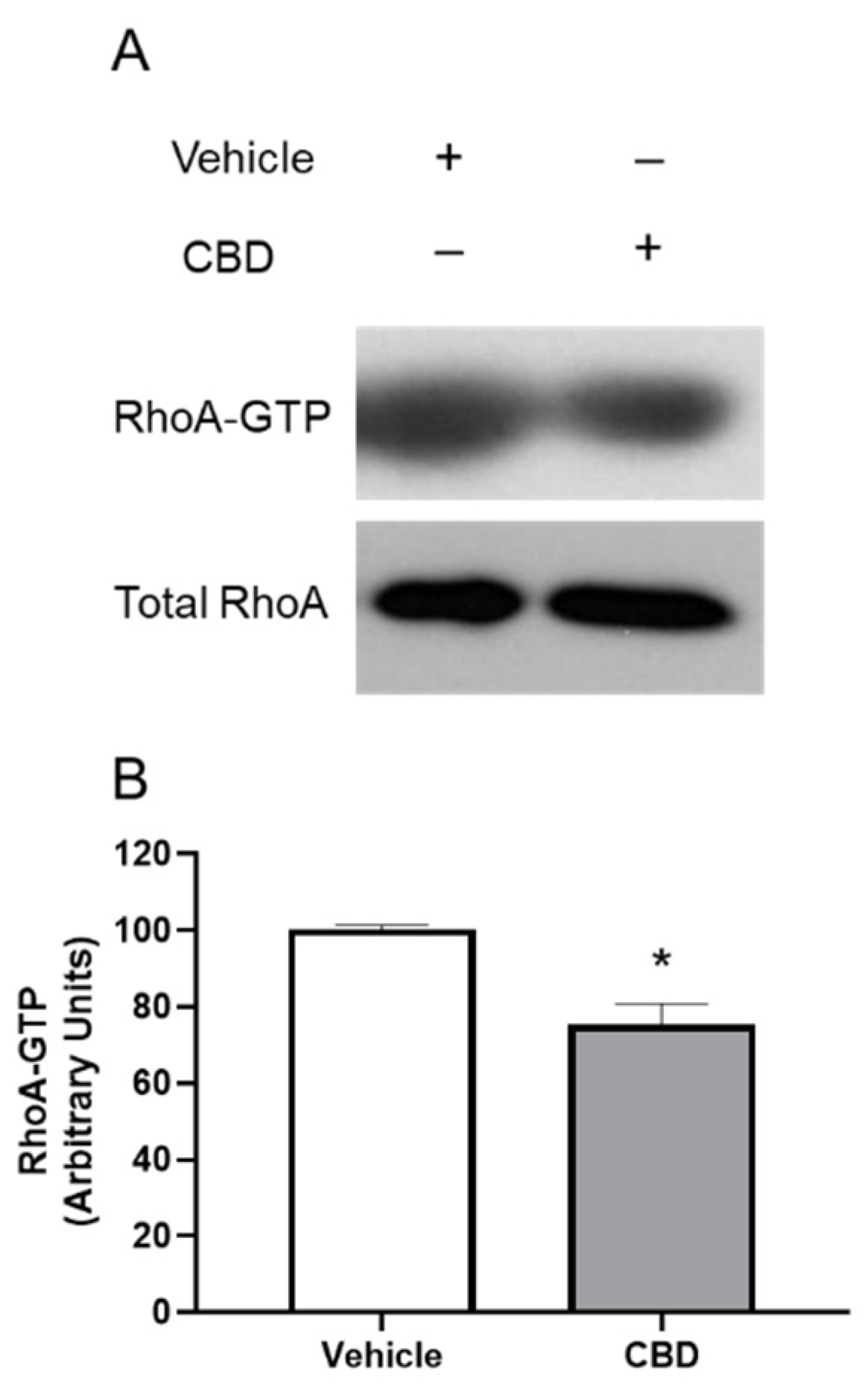
3.5. The Effects of CBD on RhoA Activation in TM Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Crowston, J.G.; Weinreb, R.N. Glaucoma medication and aqueous humor dynamics. Curr. Opin. Ophthalmol. 2005, 16, 94–100. [Google Scholar] [CrossRef]
- Quigley, H.A.; Broman, A.T. The number of people with glaucoma worldwide in 2010 and 2020. Br. J. Ophthalmol. 2006, 90, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Weinreb, R.N.; Khaw, P.T. Primary open-angle glaucoma. Lancet 2004, 363, 1711–1720. [Google Scholar] [CrossRef]
- Weinreb, R.N.; Aung, T.; Medeiros, F.A. The Pathophysiology and Treatment of Glaucoma: A review. JAMA 2014, 311, 1901–1911. [Google Scholar] [CrossRef] [PubMed]
- Shalaby, W.S.; Shankar, V.; Razeghinejad, R.; Katz, L.J. Current and new pharmacotherapeutic approaches for glaucoma. Expert Opin. Pharmacother. 2020, 21, 2027–2040. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.J.; Tsai, J.C.; Liu, J. Novel Pharmacologic Candidates for Treatment of Primary Open-Angle Glaucoma. Yale J. Biol. Med. 2017, 90, 111–118. [Google Scholar]
- Toris, C.B. Pharmacotherapies for Glaucoma. Curr. Mol. Med. 2010, 10, 824–840. [Google Scholar] [CrossRef] [PubMed]
- Schehlein, E.M.; Novack, G.; Robin, A.L. New pharmacotherapy for the treatment of glaucoma. Expert Opin. Pharmacother. 2017, 18, 1939–1946. [Google Scholar] [CrossRef] [PubMed]
- Izzo, A.A.; Borrelli, F.; Capasso, R.; Di Marzo, V.; Mechoulam, R. Non-psychotropic plant cannabinoids: New therapeutic opportunities from an ancient herb. Trends Pharmacol. Sci. 2009, 30, 515–527. [Google Scholar] [CrossRef]
- Zuardi, A.W. Cannabidiol: From an inactive cannabinoid to a drug with wide spectrum of action. Rev. Bras. Psiquiatr. 2008, 30, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Massi, P.; Solinas, M.; Cinquina, V.; Parolaro, D. Cannabidiol as potential anticancer drug. Br. J. Clin. Pharmacol. 2013, 75, 303–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, G.; De Filippis, D.; Cirillo, C.; Iuvone, T.; Capoccia, E.; Scuderi, C.; Steardo, A.; Cuomo, R.; Steardo, L. Cannabidiol in Inflammatory Bowel Diseases: A Brief Overview. Phytotherapy Res. 2012, 27, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Iuvone, T.; Esposito, G.; De Filippis, D.; Scuderi, C.; Steardo, L. Cannabidiol: A Promising Drug for Neurodegenerative Disorders? CNS Neurosci. Ther. 2009, 15, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Ruiz, J.; Sagredo, O.; Pazos, M.R.; García, C.; Pertwee, R.; Mechoulam, R.; Martínez-Orgado, J. Cannabidiol for neurodegenerative disorders: Important new clinical applications for this phytocannabinoid?: Cannabidiol and Neurodegenerative Disorders. Br. J. Clin. Pharmacol. 2013, 75, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Zuardi, A.W.; Crippa, J.A.S.; Hallak, J.E.; Bhattacharyya, S.; Atakan, Z.; Martin-Santos, R.; McGuire, P.; Guimaraes, F.S. A Critical Review of the Antipsychotic Effects of Cannabidiol: 30 Years of a Translational Investigation. Curr. Pharm. Des. 2012, 18, 5131–5140. [Google Scholar] [CrossRef] [PubMed]
- FDA. Epidiolex: Highlights of prescribing information. 2018. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2018/210365lbl.pdf (accessed on 15 August 2022).
- Hepler, R.S.; Frank, I.R. Marihuana smoking and intraocular pressure. JAMA 1971, 217, 1392. [Google Scholar] [CrossRef] [PubMed]
- Tomida, I.; Pertwee, R.G.; Azuara-Blanco, A. Cannabinoids and glaucoma. Br. J. Ophthalmol. 2004, 88, 708–713. [Google Scholar] [CrossRef]
- Aebersold, A.; Duff, M.; Sloan, L.; Song, Z.-H. Cannabidiol Signaling in the Eye and Its Potential as an Ocular Therapeutic Agent. Cell. Physiol. Biochem. 2021, 55, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Passani, A.; Posarelli, C.; Sframeli, A.T.; Perciballi, L.; Pellegrini, M.; Guidi, G.; Figus, M. Cannabinoids in Glaucoma Patients: The Never-Ending Story. J. Clin. Med. 2020, 9, 3978. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.V.; Deng, P.; Sasaki, Y.; Epstein, D.L. Regulation of myosin light chain phosphorylation in the trabecular meshwork: Role in aqueous humour outflow facility. Exp. Eye Res. 2005, 80, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Lee, C.; Read, A.T.; Wang, K.; Ha, J.; Kuhn, M.; Navarro, I.; Cui, J.; Young, K.; Gorijavolu, R.; et al. Anti-fibrotic activity of a rho-kinase inhibitor restores outflow function and intraocular pressure homeostasis. eLife 2021, 10, e60831. [Google Scholar] [CrossRef] [PubMed]
- Schehlein, E.M.; Robin, A.L. Rho-Associated Kinase Inhibitors: Evolving Strategies in Glaucoma Treatment. Drugs 2019, 79, 1031–1036. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.V.; Deng, P.F.; Kumar, J.; Epstein, D.L. Modulation of aqueous humor outflow facility by the Rho kinase-specific inhibitor Y-27632. Investig. Ophthalmol. Vis. Sci. 2001, 42, 1029–1037. [Google Scholar]
- Wiederholt, M.; Thieme, H.; Stumpff, F. The regulation of trabecular meshwork and ciliary muscle contractility. Prog. Retin. Eye Res. 2000, 19, 271–295. [Google Scholar] [CrossRef]
- Amano, M.; Nakayama, M.; Kaibuchi, K. Rho-kinase/ROCK: A key regulator of the cytoskeleton and cell polarity. Cytoskeleton 2010, 67, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Khurana, R.N.; Deng, P.-F.; Epstein, D.L.; Rao, P.V. The role of protein kinase C in modulation of aqueous humor outflow facility. Exp. Eye Res. 2002, 76, 39–47. [Google Scholar] [CrossRef]
- Kaufman, P.L.; Gabelt, B.; Tian, B.; Liu, X. Advances in Glaucoma Diagnosis and Therapy for the Next Millennium: New Drugs for Trabecular and Uveoscleral Outflow. Semin. Ophthalmol. 1999, 14, 130–143. [Google Scholar] [CrossRef]
- Epstein, D.L.; Rowlette, L.L.; Roberts, B.C. Acto-myosin drug effects and aqueous outflow function. Investig. Ophthalmol. Vis. Sci. 1999, 40, 74–81. [Google Scholar]
- Vitale, R.; Iannotti, F.; Amodeo, P. The (Poly)Pharmacology of Cannabidiol in Neurological and Neuropsychiatric Disorders: Molecular Mechanisms and Targets. Int. J. Mol. Sci. 2021, 22, 4876. [Google Scholar] [CrossRef]
- Zhong, L.; Geng, L.; Njie, Y.; Feng, W.; Song, Z.-H. CB2 Cannabinoid Receptors in Trabecular Meshwork Cells Mediate JWH015-Induced Enhancement of Aqueous Humor Outflow Facility. Investig. Opthalmol. Vis. Sci. 2005, 46, 1988–1992. [Google Scholar] [CrossRef]
- Njie, Y.F.; Kumar, A.; Qiao, Z.; Zhong, L.; Song, Z.-H. Noladin Ether Acts on Trabecular Meshwork Cannabinoid (CB1) Receptors to Enhance Aqueous Humor Outflow Facility. Investig. Opthalmol. Vis. Sci. 2006, 47, 1999–2005. [Google Scholar] [CrossRef] [PubMed]
- Njie, Y.F.; He, F.; Qiao, Z.; Song, Z.-H. Aqueous humor outflow effects of 2-arachidonylglycerol. Exp. Eye Res. 2008, 87, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Njie, Y.F.; Qiao, Z.; Xiao, Z.; Wang, W.; Song, Z.-H. N-arachidonylethanolamide-Induced Increase in Aqueous Humor Outflow Facility. Investig. Opthalmol. Vis. Sci. 2008, 49, 4528–4534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradley, J.M.; Vranka, J.; Colvis, C.M.; Conger, D.M.; Alexander, J.P.; Fisk, A.S.; Samples, J.R.; Acott, T.S. Effect of matrix metalloproteinases activity on outflow in perfused human organ culture. Investig. Ophthalmol. Vis. Sci. 1998, 39, 2649–2658. [Google Scholar]
- Polansky, J.R.; Weinreb, R.N.; Baxter, J.D.; Alvarado, J. Human trabecular cells. I. Establishment in tissue culture and growth characteristics. Investig. Ophthalmol. Vis. Sci. 1979, 18, 1043–1049. [Google Scholar]
- Tripathi, R.C.; Tripathi, B.J. Human trabecular endothelium, corneal endothelium, keratocytes, and scleral fibroblasts in primary cell culture. A comparative study of growth characteristics, morphology, and phagocytic activity by light and scanning electron microscopy. Exp. Eye Res. 1982, 35, 611–624. [Google Scholar] [CrossRef]
- Nakamura, Y.; Hirano, S.; Suzuki, K.; Seki, K.; Sagara, T.; Nishida, T. Signaling mechanism of TGF-beta1-induced collagen contraction mediated by bovine trabecular meshwork cells. Investig. Ophthalmol. Vis. Sci. 2002, 43, 74–81. [Google Scholar]
- He, F.; Song, Z.-H. Molecular and cellular changes induced by the activation of CB2 cannabinoid receptors in trabecular meshwork cells. Mol. Vis. 2007, 13, 1348–1356. [Google Scholar]
- Colasanti, B.K.; Brown, R.E.; Craig, C.R. Ocular hypotension, ocular toxicity, and neurotoxicity in response to marihuana extract and cannabidiol. Gen. Pharmacol. Vasc. Syst. 1984, 15, 479–484. [Google Scholar] [CrossRef]
- Green, K.; Wynn, H.; Bowman, K.A. A comparison of topical cannabinoids on intraocular pressure. Exp. Eye Res. 1978, 27, 239–246. [Google Scholar] [CrossRef]
- Perez-reyes, M.; Wagner, D.; Wall, M.E.; Davis, K.H. Intravenous administration of cannabinoids and intraocular pressure. In Pharmacology of Marihuana; Raven Press: New York, NY, USA, 1976; pp. 829–832. [Google Scholar]
- Rao, V.P.; Epstein, D.L. Rho GTPase/Rho Kinase Inhibition as a Novel Target for the Treatment of Glaucoma. BioDrugs 2007, 21, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Lincoln, T.M. Myosin Phosphatase Regulatory Pathways. Circ. Res. 2007, 100, 10–12. [Google Scholar] [CrossRef] [PubMed]
- McGregor, I.S.; Cairns, E.A.; Abelev, S.; Cohen, R.; Henderson, M.; Couch, D.; Arnold, J.C.; Gauld, N. Access to cannabidiol without a prescription: A cross-country comparison and analysis. Int. J. Drug Policy 2020, 85, 102935. [Google Scholar] [CrossRef]
- Iffland, K.; Grotenhermen, F. An Update on Safety and Side Effects of Cannabidiol: A Review of Clinical Data and Relevant Animal Studies. Cannabis Cannabinoid Res. 2017, 2, 139–154. [Google Scholar] [CrossRef]
- Chesney, E.; Oliver, D.; Green, A.; Sovi, S.; Wilson, J.; Englund, A.; Freeman, T.; McGuire, P. Adverse effects of cannabidiol: A systematic review and meta-analysis of randomized clinical trials. Neuropsychopharmacology 2020, 45, 1799–1806. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.; Daily, L.; Leishman, E.; Bradshaw, H.; Straiker, A. Δ9-Tetrahydrocannabinol and Cannabidiol Differentially Regulate Intraocular Pressure. Investig. Opthalmol. Vis. Sci. 2018, 59, 5904–5911. [Google Scholar] [CrossRef] [PubMed]
- Rebibo, L.; Frušić-Zlotkin, M.; Ofri, R.; Nassar, T.; Benita, S. The dose-dependent effect of a stabilized cannabidiol nanoemulsion on ocular surface inflammation and intraocular pressure. Int. J. Pharm. 2022, 617, 121627. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aebersold, A.S.; Song, Z.-H. The Effects of Cannabidiol on Aqueous Humor Outflow and Trabecular Meshwork Cell Signaling. Cells 2022, 11, 3006. https://doi.org/10.3390/cells11193006
Aebersold AS, Song Z-H. The Effects of Cannabidiol on Aqueous Humor Outflow and Trabecular Meshwork Cell Signaling. Cells. 2022; 11(19):3006. https://doi.org/10.3390/cells11193006
Chicago/Turabian StyleAebersold, Alyssa S., and Zhao-Hui Song. 2022. "The Effects of Cannabidiol on Aqueous Humor Outflow and Trabecular Meshwork Cell Signaling" Cells 11, no. 19: 3006. https://doi.org/10.3390/cells11193006
APA StyleAebersold, A. S., & Song, Z.-H. (2022). The Effects of Cannabidiol on Aqueous Humor Outflow and Trabecular Meshwork Cell Signaling. Cells, 11(19), 3006. https://doi.org/10.3390/cells11193006