
Nrf2-Mediated Dichotomy in the Vascular System: Mechanistic and Therapeutic Perspective

Abstract
:1. Introduction
2. Nrf2 Signaling
2.1. Basic Functions
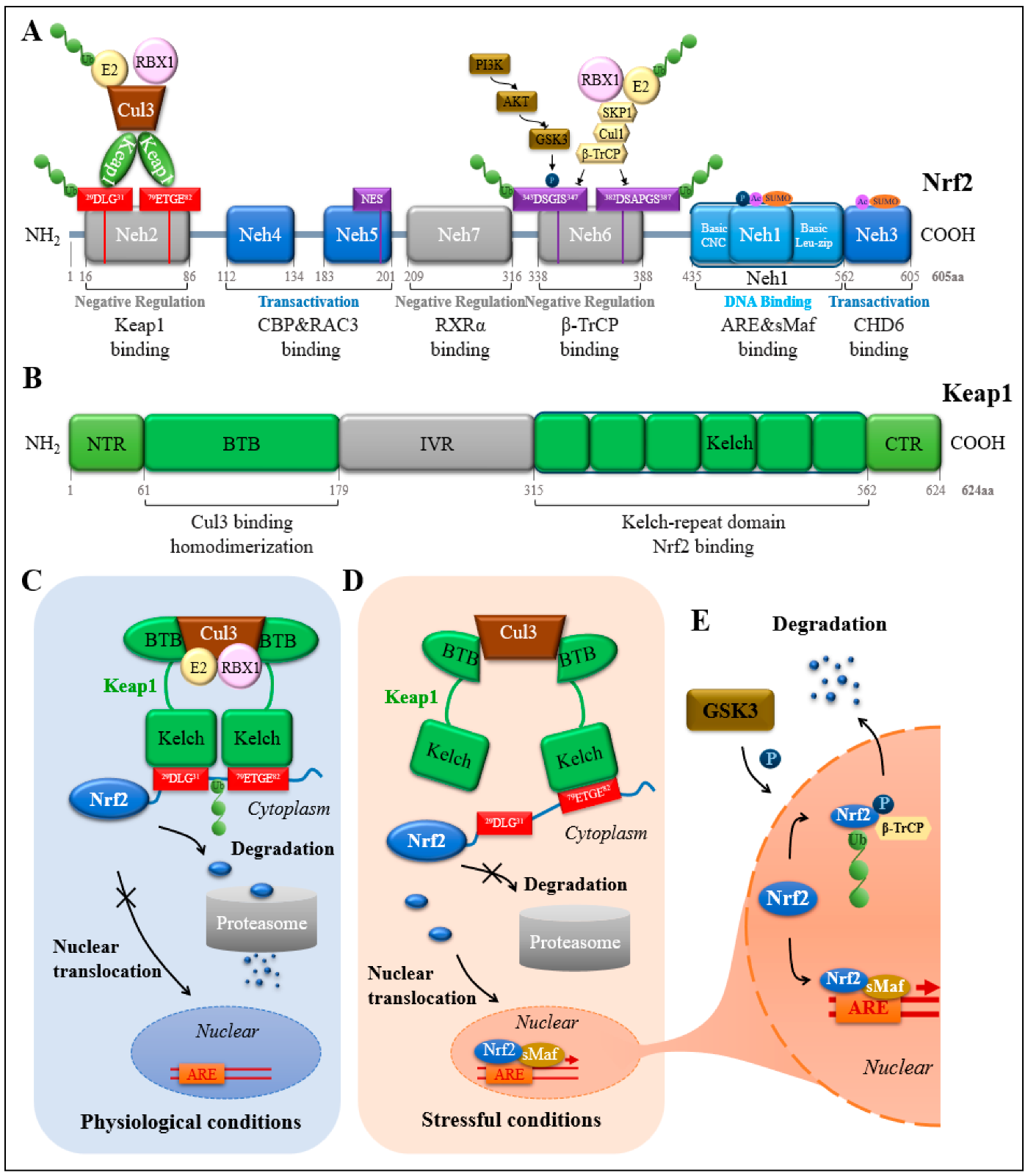
2.2. Protein Structural Domain of Nrf2
2.3. Nrf2 Transcription-Related
2.4. Ubiquitinated Degradation of Nrf2
3. Nrf2 and Vascular Physiology and Pathology
3.1. Nrf2 and Endothelial Cells

{kind=link}
| Approach | Nrf2 Signaling | Finding | Reference |
|---|---|---|---|
| Compounds | Atherosclerosis (AS) Related | ||
| Melatonin | Nrf2/NLRP3 | Reduces cigarette smoke extract (CSE) treatment-induced pyroptosis in human aortic endothelial cells (HAECs) in vitro and cigarette smoke exposure-enhanced intimal hyperplasia in rat carotid arteries induced by balloon injury in vivo | [75] |
| Astragaloside | Nrf2/HO-1 | Prevents oxidized low-density lipoprotein (oxLDL)-induced human umbilical vein endothelial cells (HUVECs) injury in vitro | [87] |
| Isoflavone | Nrf2/AER | Reduces oxLDL-induced oxidative stress damage in EA.hy926 cells in vitro and atherosclerosis in apolipoprotein E deficiency (ApoE−/−) mice fed with a high-fat diet in vivo | [88] |
| Flavonoids from a Deep-Sea-Derived Arthrinium sp. | AKT/Nrf2/HO-1 | Protects HUVECs against ox-LDL-induced oxidative stress in vitro | [89] |
| Tanshinone IIA | Nrf2 | Suppresses human coronary artery endothelial cells (HCAECs) ferroptosis in vitro | [90] |
| Chalcone derivative | Nrf2/HO-1 | Inhibits cholesterol efflux and suppresses inflammatory responses in HUVECs in vitro Reduces lipid accumulation and plaque formation in LDL receptor knockout (Ldlr−/−) mice fed with a high-fat diet | [91] |
| Ilexgenin A | Nrf2/PSMB5 | Suppresses mitochondrial fission, and improves endothelial dysfunction induced by palmitate (PA) in vitro | [92] |
| Acacetin | Nrf2 | Exerts antioxidant potential in ApoE−/− mice and in EA.hy926 cells induced by human oxLDL in vitro | [93] |
| Irisin | AKT/mTOR/S6K1/Nrf2 | Attenuates oxLDL-impaired angiogenesis of human microvascular ECs in vitro and in a chicken embryo membrane (CAM) model in vivo | [94] |
| A peptide from microalgae Isochrysis zhanjiangensis | Nrf2 | Inhibits oxLDL-induced inflammation and apoptosis of HUVECs in vitro | [95] |
| Kaempferol | PI3K/AKT/Nrf2 | Attenuates oxLDL-induced injury via activating G protein-coupled estrogen receptor (GPER) associated with upregulation of PI3K/AKT/Nrf2 signaling in HAECs in vitro Suppresses atherosclerotic lesion formation in ovariectomized ApoE−/− mice fed with a high-fat diet in vivo | [96] |
| Xanthoangelol | Nrf2/ARE | Prevents oxLDL-induced HUVECs injury in vitro | [97] |
| Equol | Nrf2/t-BHP/CHOP | Attenuates atherosclerosis in ApoE−/− mice fed with a high-fat diet in vivo and endoplasmic reticulum stress and apoptosis in HUVECs induced by tert-butyl hydroperoxide (t-BHP) and thapsigargin in vitro | [98] |
| Kirenol | PI3K/AKT/Nrf2 | Prevents B[a]P-induced redox imbalance in HUVECs in vitro | [99] |
| Resveratrol | Nrf2/ICAM-1 | Suppressive effects on pro-inflammatory responses in ECs and accelerated atherosclerosis in carotid arteries induced by ApoE−/− mice | [100] |
| Zedoarondiol | Nrf2/HO-1 | Attenuates oxLDL-induced injury, oxidative stress, and inflammatory responses in HUVECs in vitro | [101] |
| Dietary ellagic acid | Nrf2/HO-1 | Protective effects on damage in HAECs induced by hypochlorous acid (HOCl) in vitro and endothelial dysfunction in the mouse model of accelerated atherosclerosis in carotid arteries induced by partial ligation in vivo | [102] |
| Salvianolic acid B | Nrf2/HO-1 | Inhibits tumor necrosis factor-alpha-induced NF-kappaB activation in HUVECs | [103] |
| Dihydromyricetin | ERK&AKT/Nrf2/HO-1 | Protects HUVECs from oxLDL-induced oxidative injury in vitro | [104] |
| Miltirone | Nrf2/HO-1 | Protects human EA.hy926 ECs from oxidative stress-associated injury induced by t-BHP and oxLDL in vitro | [105] |
| Z-Ligustilide | Nrf2/ARE | Protects EA.hy926 cells from t-BHP-induced oxidative stress in vitro and attenuates atherogenesis in Ldlr−/− mice fed with a high-fat diet in vivo | [106] |
| Nrf2/HO-1 | Attenuates inflammatory responses in HUVECs in vitro | [107] | |
| Vitexin | Wnt/beta-catenin and Nrf2 | Protects HUVECs from high glucose-induced injury in vitro | [108] |
| Theaflavin | miR-24/Nrf2/HO-1 | Alleviates oxidative injury and atherosclerosis progression in ApoE−/− mice fed with a high-fat diet in vivo and protects against cholesterol-induced oxidative injuries in HUVECs in vitro | [109] |
| PI3K/AKT/Nrf2 | Attenuates t-BHP-induced oxidative stress in HUVECs in vitro and enhances vascularization in regenerated tissues and accelerates wound healing in vivo | [110] | |
| β-Farrerol | GSK-3/Nrf2-ARE | Protect EA.hy926 cells against oxidative stress-induced injuries in vitro | [111] |
| Compounds | Diabetes Related | ||
| Metformin | Nrf2 | Ameliorates the inhibitory effect of high glucose on migration and angiogenesis of HUVECs in vitro | [84] |
| Carnosol | Nrf2/t-BHP | Protects against t-BHP-induced human retinal microvascular endothelial cells (HRMECs) injury | [112] |
| β-Buyang Huanwu Decoction | AKT/GSK3/Nrf2 | Enhances revascularization in a mouse model of diabetic hindlimb ischemia (HLI) | [113] |
| Antrodin C | Nrf2/HO-1 | Prevents hyperglycemia-induced senescence and apoptosis in human ECs in vitro | [114] |
| Allicin | Nrf2 | Alleviates aortic inflammatory responses associated with type 1 diabetes induced by intraperitoneal injection of streptozotocin (STZ) in mice and high glucose-induced growth inhibition and death in HUVECs in vitro | [115] |
| Compounds | Hypertension Related | ||
| Tilapia by-product oligopeptide | Nrf2/NF-κB | Protects against Ang II-induced hypertensive injury in HUVECs in vitro | [82] |
| A novel angiotensin-I-converting enzyme inhibitory peptide from microalgae Isochrysis zhanjiangensis | Nrf2 | Inhibits Ang II-induced vascular factor secretion, inflammatory responses, and apoptosis in HUVECs in vitro | [116] |
| Hydrogen sulfide | Nrf2 | Ameliorates endothelial dysfunction associated with hypertension in spontaneously hypertensive rats (SHR) and Ang II-induced cellular damage and ROS formation in HUVECs in vitro | [117] |
| Compounds | Other Diseases Related | ||
| Celastrol | Nrf2/ERK1/2/Nox2 | Attenuates Ang II-mediated HUVECs damage in vitro | [78] |
| Schisandrin C | Keap1/Nrf2 | As an antioxidative agent for the treatment of Ang II-induced vascular endothelial deficits in vitro | [80] |
| Osthole | Keap1/Nrf2 | Protects against Ang II-induced apoptosis of rat aortic endothelial cells (RAECs) in vitro | [81] |
| Memantine | Nrf2/HO-1 | Protects against inflammatory responses and impaired endothelial tube formation induced by oxygen-glucose deprivation/reperfusion in HUVECs in vitro | [83] |
| Cyanidin-3-O-glucoside | Nrf2/Bach1 and NF-κB | Improves intracellular redox status of HUVECs exposed to palmitic acid (PA) in vitro | [118] |
| Panax notoginseng Saponin | PI3K/AKT/Nrf2 | Protects against cerebral ischemia/reperfusion (I/R)-induced blood–brain barrier disruption in cerebral microvascular endothelial cells (bEnd.3) in vitro | [119] |
| Ginsenoside Rg3 | Nrf2/ARE | Antagonizes adriamycin-induced cardiotoxicity by improving endothelial dysfunction both in vivo and in vitro | [120] |
| Procyanidin B2 | Nrf2/PPARγ/sFlt-1 | Ameliorates endothelial dysfunction and impaired angiogenesis of HUVECs in vitro and in a rat model of preeclampsia induced by uterine perfusion pressure (RUPP) in vivo | [121] |
| Rice bran phenolic Compounds | Nrf2/HO-1/NQO1/eNOS | An antioxidant/anti-inflammatory effect on HUVECs with induced oxidative stress in vitro | [122] |
| Chlorogenic acid | Nrf2/HO-1 | Protective effects on Ang II infusion-induced vascular senescence in mice and H2O2-induced senescence in HUVECs in vitro | [123] |
| Pterostilbene | Keap1/Nrf2/HO-1 | Protects against uraemic serum (US)-mediated injury in HUVECs in vitro | [124] |
| Lipoxin A4 | Nrf2/HO-1 | Attenuates H2O2-evoked cytotoxic injury in HUVECs in vitro | [125] |
| Hydrogen sulfide | Nrf2/HIF-1α | Prevents balloon injury-induced neointimal hyperplasia in carotid arteries of rats in vivo and enhances HUVECs tube formation and migration in vitro | [126] |
| Paeoniflorin | Nrf2/HO-1 | Alleviates t-BHP-stimulated HUVECs cellular dysfunction and apoptosis in vitro and enhances the vascularization of regenerated tissues and promotes flap survival in vivo | [127] |
| Irigenin | Nrf2 | Alleviates Ang II-induced oxidative stress and apoptosis in HUVECs in vitro | [128] |
| Cinnamaldehyde | Nrf2/HO-1 | Protects against H2O2 or TNFα-induced inflammatory responses in HUVECs and inhibits lipopolysaccharide (LPS)-induced inflammatory cell infiltration in vivo | [129] |
| Protandim | Nrf2 | Protects against salt-induced vascular dysfunction in vivo by restoring redox homeostasis in the vasculature | [130] |
| Magnesium lithospermate B | PI3K/AKT/Nrf2 | Protects against LPS-induced endothelial dysfunction in a murine acute inflammation model and in human dermal microvascular endothelial cells (HMECs-1) in vitro | [131] |
| Ginkgo biloba extract | Nrf2/HO-1 | Reduces leukocyte adherence to injury arteries, enhances HO-1 expression in circulating monocytes and arteries after wire injury, and reduces TNF-alpha-stimulated endothelial adhesiveness | [132] |
| Brazilian Green Propolis | PI3K/AKT/mTOR/Nrf2/HO-1 | Inhibits oxLDL-stimulated oxidative stress in HUVECs in vitro | [133] |
| Unripe Carica papaya fruit extract | Nrf2 | Protects ECs challenged with H2O2 in vitro | [134] |
| Lycopene | Nrf2/HO-1 | Inhibits cyclic strain-induced ET-1 gene expression of HUVECs in vitro | [135] |
| Hippocampus abdominalis-derived peptides | Nrf2/HO-1 | Protects H2O2-induced cell death through antiapoptotic action in vitro | [136] |
| Aspirin eugenol ester | Nrf2 | Attenuates oxidative injury in a hamster model of atherosclerosis induced by a high-fat diet and H2O2-induced apoptosis in HUVECs, an in vitro model of oxidative stress | [137] |
| miRNAs | |||
| miR-24 | Nrf2/HO-1 | Promotes endothelial repair caused by oxidative stress after balloon injury in diabetic rats | [138] |
| miR-200a | Nrf2 | Thymosin beta-4 (Tbeta4) attenuates H/R-induced cardiac microvascular endothelial cells (CMECs) injury by miR-200a-Nrf2 signaling in vitro | [139] |
| miR-140-5p | Nrf2 and Sirt2 | Upregulation of miR-140-5p in the atherosclerotic aorta in ApoE−/− mice Increases oxidative stress in HUVECs in vitro | [140] |
3.2. Nrf2 and Vascular Smooth Muscle Cells
3.3. Nrf2 and Vascular Disease
| Nrf2 Signaling | Animal Model | Pathological Setting | Treatment | Phenotype | Proposed Mechanism | Reference |
|---|---|---|---|---|---|---|
| Nrf2/CD36 | Nrf2−/−, ApoE−/− | Atherosclerosis | None | Nrf2 knockout decreases susceptibility to atherosclerotic plaque formation in ApoE−/− mice. | Nrf2 upregulates CD36 thereby promoting foam cell formation and the pathogenesis of atherosclerosis. | [177] |
| Nrf2/CD36 | Nrf2−/−, ApoE−/− | Atherosclerosis | None | Nrf2 expression promotes atherosclerotic lesion formation. | Nrf2 deficiency results in decreased cholesterol Influx, correlated with lower CD36 expression. | [178] |
| Nrf2/IL-1 | Nrf2−/−, ApoE−/− | Atherosclerosis | None | Nrf2-deficient ApoE−/− mice are highly protected against diet-induced atherogenesis. | Nrf2 aggravates atherosclerosis by enhancing IL-1-mediated vascular inflammation. | [179] |
| Nrf2 | Nrf2−/−, ApoE−/− | Atherosclerosis | None | Nrf2 deletion in bone marrow-derived cells is protective against atherosclerosis. | Nrf2−/−; ApoE−/− decreases expression of macrophage M1-subtype genes in mice. | [180] |
| Nrf2 | ApoE−/− | Atherosclerosis | PCB29-pQ | PCB29-pQ driven CD163+ macrophage accumulated in the aortic valve. | Nrf2 activation is the main reason for macrophage differentiation. | [181] |
| Nrf2 | Atherosclerosis | Vitamin E | Nrf2 can mediate both pro-and anti-atherosclerotic effects. | Nrf2 activation can promote CD36-mediated cholesterol uptake by macrophages, increase induction of Nrf2-mediated antioxidant genes, and is likely to contribute to decreased lesion progression. | [182] | |
| Nrf2 | Nrf2−/−, Ldlr−/−, Apob100/100 | Atherosclerosis | None | Nrf2 deficiency impairs atherosclerotic lesion development but promotes the features of plaque instability in hyper-cholesterolemic mice. | Nrf2 deficiency in Ldlr−/− mice reduces total plasma cholesterol and triglycerides in Ldlr−/−ApoB100/100 mice and aggravates aortic plaque maturation as it increased plaque calcification. | [183] |
| Nrf2 | Nrf2−/− | Abdominal aortic aneurysm | None | Lacking Nrf2 transcriptional activity attenuates AAA formation under simvastatin treatment. | The inhibition of Nrf2 transcriptional activity facilitates AAA formation in mice, which can be prevented by simvastatin. | [191] |
4. Therapeutic Potential of Targeting Nrf2 Pathway
5. Closing Remark
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Y.; Murugesan, P.; Huang, K.; Cai, H. NADPH oxidases and oxidase crosstalk in cardiovascular diseases: Novel therapeutic targets. Nat. Rev. Cardiol. 2020, 17, 170–194. [Google Scholar] [CrossRef] [PubMed]
- Sack, M.N.; Fyhrquist, F.Y.; Saijonmaa, O.J.; Fuster, V.; Kovacic, J.C. Basic Biology of Oxidative Stress and the Cardiovascular System: Part 1 of a 3-Part Series. J. Am. Coll. Cardiol. 2017, 70, 196–211. [Google Scholar] [CrossRef] [PubMed]
- Munzel, T.; Camici, G.G.; Maack, C.; Bonetti, N.R.; Fuster, V.; Kovacic, J.C. Impact of Oxidative Stress on the Heart and Vasculature: Part 2 of a 3-Part Series. J. Am. Coll. Cardiol. 2017, 70, 212–229. [Google Scholar] [CrossRef] [PubMed]
- Niemann, B.; Rohrbach, S.; Miller, M.R.; Newby, D.E.; Fuster, V.; Kovacic, J.C. Oxidative Stress and Cardiovascular Risk: Obesity, Diabetes, Smoking, and Pollution: Part 3 of a 3-Part Series. J. Am. Coll. Cardiol. 2017, 70, 230–251. [Google Scholar] [CrossRef]
- Munzel, T.; Gori, T.; Bruno, R.M.; Taddei, S. Is oxidative stress a therapeutic target in cardiovascular disease? Eur. Heart J. 2010, 31, 2741–2748. [Google Scholar] [CrossRef]
- Burgoyne, J.R.; Mongue-Din, H.; Eaton, P.; Shah, A.M. Redox signaling in cardiac physiology and pathology. Circ. Res. 2012, 111, 1091–1106. [Google Scholar] [CrossRef]
- Burgoyne, J.R.; Din, H.M.; Eaton, P.; Shah, A.M. Response to Detailed aspects of redox signaling in cardiac physiology and pathology. Circ. Res. 2013, 112, e2. [Google Scholar] [CrossRef]
- Stocker, R.; Keaney, J.F., Jr. Role of oxidative modifications in atherosclerosis. Physiol. Rev. 2004, 84, 1381–1478. [Google Scholar] [CrossRef]
- Freed, J.K.; Gutterman, D.D. Mitochondrial reactive oxygen species and vascular function: Less is more. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 673–675. [Google Scholar] [CrossRef]
- Zu, X.; Yu, L.; Sun, Y.; Tian, J.; Liu, F.; Sun, Q.; He, S.; Sun, G.; Luo, W.; Jiang, Y. Global mapping of ZBTB7A transcription factor binding sites in HepG2 cells. Cell. Mol. Biol. Lett. 2010, 15, 260–271. [Google Scholar] [CrossRef]
- Qin, Q.; Qu, C.; Niu, T.; Zang, H.; Qi, L.; Lyu, L.; Wang, X.; Nagarkatti, M.; Nagarkatti, P.; Janicki, J.S.; et al. Nrf2-Mediated Cardiac Maladaptive Remodeling and Dysfunction in a Setting of Autophagy Insufficiency. Hypertension 2016, 67, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Zang, H.; Wu, W.; Qi, L.; Tan, W.; Nagarkatti, P.; Nagarkatti, M.; Wang, X.; Cui, T. Autophagy Inhibition Enables Nrf2 to Exaggerate the Progression of Diabetic Cardiomyopathy in Mice. Diabetes 2020, 69, 2720–2734. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Qin, Q.; Ding, Y.; Zang, H.; Li, D.S.; Nagarkatti, M.; Nagarkatti, P.; Wang, W.; Wang, X.; Cui, T. Autophagy Controls Nrf2-Mediated Dichotomy in Pressure Overloaded Hearts. Front. Physiol. 2021, 12, 673145. [Google Scholar] [CrossRef] [PubMed]
- Zang, H.; Mathew, R.O.; Cui, T. The Dark Side of Nrf2 in the Heart. Front. Physiol. 2020, 11, 722. [Google Scholar] [CrossRef]
- Yamamoto, M.; Kensler, T.W.; Motohashi, H. The KEAP1-NRF2 System: A Thiol-Based Sensor-Effector Apparatus for Maintaining Redox Homeostasis. Physiol. Rev. 2018, 98, 1169–1203. [Google Scholar] [CrossRef]
- Li, J.; Ichikawa, T.; Janicki, J.S.; Cui, T. Targeting the Nrf2 pathway against cardiovascular disease. Expert Opin. Ther. Targets 2009, 13, 785–794. [Google Scholar] [CrossRef]
- Maher, J.; Yamamoto, M. The rise of antioxidant signaling—The evolution and hormetic actions of Nrf2. Toxicol. Appl. Pharmacol. 2010, 244, 4–15. [Google Scholar] [CrossRef]
- Chan, J.Y.; Kwong, M.; Lo, M.; Emerson, R.; Kuypers, F.A. Reduced oxidative-stress response in red blood cells from p45NFE2-deficient mice. Blood 2001, 97, 2151–2158. [Google Scholar] [CrossRef]
- Tiwari, S.; Italiano, J.E., Jr.; Barral, D.C.; Mules, E.H.; Novak, E.K.; Swank, R.T.; Seabra, M.C.; Shivdasani, R.A. A role for Rab27b in NF-E2-dependent pathways of platelet formation. Blood 2003, 102, 3970–3979. [Google Scholar] [CrossRef]
- Lecine, P.; Italiano, J.E., Jr.; Kim, S.W.; Villeval, J.L.; Shivdasani, R.A. Hematopoietic-specific beta 1 tubulin participates in a pathway of platelet biogenesis dependent on the transcription factor NF-E2. Blood 2000, 96, 1366–1373. [Google Scholar] [CrossRef] [Green Version]
- Leung, L.; Kwong, M.; Hou, S.; Lee, C.; Chan, J.Y. Deficiency of the Nrf1 and Nrf2 transcription factors results in early embryonic lethality and severe oxidative stress. J. Biol. Chem. 2003, 278, 48021–48029. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, A.; Ito, E.; Toki, T.; Kogame, K.; Takahashi, S.; Igarashi, K.; Hayashi, N.; Yamamoto, M. Molecular cloning and functional characterization of a new Cap‘n’ collar family transcription factor Nrf3. J. Biol. Chem. 1999, 274, 6443–6452. [Google Scholar] [CrossRef] [PubMed]
- Derjuga, A.; Gourley, T.S.; Holm, T.M.; Heng, H.H.; Shivdasani, R.A.; Ahmed, R.; Andrews, N.C.; Blank, V. Complexity of CNC transcription factors as revealed by gene targeting of the Nrf3 locus. Mol. Cell. Biol. 2004, 24, 3286–3294. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Pepe, A.E.; Wang, G.; Luo, Z.; Zhang, L.; Zeng, L.; Zhang, Z.; Hu, Y.; Ye, S.; Xu, Q. Nrf3-Pla2g7 interaction plays an essential role in smooth muscle differentiation from stem cells. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 730–744. [Google Scholar] [CrossRef]
- Pepe, A.E.; Xiao, Q.; Zampetaki, A.; Zhang, Z.; Kobayashi, A.; Hu, Y.; Xu, Q. Crucial role of nrf3 in smooth muscle cell differentiation from stem cells. Circ. Res. 2010, 106, 870–879. [Google Scholar] [CrossRef]
- Oyake, T.; Itoh, K.; Motohashi, H.; Hayashi, N.; Hoshino, H.; Nishizawa, M.; Yamamoto, M.; Igarashi, K. Bach proteins belong to a novel family of BTB-basic leucine zipper transcription factors that interact with MafK and regulate transcription through the NF-E2 site. Mol. Cell. Biol. 1996, 16, 6083–6095. [Google Scholar] [CrossRef]
- Hayes, J.D.; Dinkova-Kostova, A.T. The Nrf2 regulatory network provides an interface between redox and intermediary metabolism. Trends Biochem. Sci. 2014, 39, 199–218. [Google Scholar] [CrossRef]
- Cui, T.; Lai, Y.; Janicki, J.S.; Wang, X. Nuclear factor erythroid-2 related factor 2 (Nrf2)-mediated protein quality control in cardiomyocytes. Front. Biosci. (Landmark Ed.) 2016, 21, 192–202. [Google Scholar] [CrossRef]
- Kopacz, A.; Kloska, D.; Forman, H.J.; Jozkowicz, A.; Grochot-Przeczek, A. Beyond repression of Nrf2: An update on Keap1. Free Radic. Biol. Med. 2020, 157, 63–74. [Google Scholar] [CrossRef]
- Silva-Islas, C.A.; Maldonado, P.D. Canonical and non-canonical mechanisms of Nrf2 activation. Pharmacol. Res. 2018, 134, 92–99. [Google Scholar] [CrossRef]
- Fukutomi, T.; Takagi, K.; Mizushima, T.; Ohuchi, N.; Yamamoto, M. Kinetic, thermodynamic, and structural characterizations of the association between Nrf2-DLGex degron and Keap1. Mol. Cell. Biol. 2014, 34, 832–846. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Rojo de la Vega, M.; Schmidlin, C.J.; Ooi, A.; Zhang, D.D. Kelch-like ECH-associated protein 1 (KEAP1) differentially regulates nuclear factor erythroid-2-related factors 1 and 2 (NRF1 and NRF2). J. Biol. Chem. 2018, 293, 2029–2040. [Google Scholar] [CrossRef] [PubMed]
- Katoh, Y.; Itoh, K.; Yoshida, E.; Miyagishi, M.; Fukamizu, A.; Yamamoto, M. Two domains of Nrf2 cooperatively bind CBP, a CREB binding protein, and synergistically activate transcription. Genes Cells 2001, 6, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.; Wilson, C.J.; Karunatilleke, N.C.; Moselhy, M.H.; Karttunen, M.; Choy, W.Y. Exploring the Conformational Landscape of the Neh4 and Neh5 Domains of Nrf2 Using Two Different Force Fields and Circular Dichroism. J. Chem. Theory Comput. 2021, 17, 3145–3156. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Chin, Y.E.; Zhang, D.D. Acetylation of Nrf2 by p300/CBP augments promoter-specific DNA binding of Nrf2 during the antioxidant response. Mol. Cell. Biol. 2009, 29, 2658–2672. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Yu, S.; Chen, J.D.; Kong, A.N. The nuclear cofactor RAC3/AIB1/SRC-3 enhances Nrf2 signaling by interacting with transactivation domains. Oncogene 2013, 32, 514–527. [Google Scholar] [CrossRef]
- Wang, H.; Liu, K.; Geng, M.; Gao, P.; Wu, X.; Hai, Y.; Li, Y.; Li, Y.; Luo, L.; Hayes, J.D.; et al. RXRalpha inhibits the NRF2-ARE signaling pathway through a direct interaction with the Neh7 domain of NRF2. Cancer Res. 2013, 73, 3097–3108. [Google Scholar] [CrossRef]
- Jiang, S.; Yang, Y.; Li, T.; Ma, Z.; Hu, W.; Deng, C.; Fan, C.; Lv, J.; Sun, Y.; Yi, W. An overview of the mechanisms and novel roles of Nrf2 in cardiovascular diseases. Expert Opin. Ther. Targets 2016, 20, 1413–1424. [Google Scholar] [CrossRef]
- Rada, P.; Rojo, A.I.; Chowdhry, S.; McMahon, M.; Hayes, J.D.; Cuadrado, A. SCF/{beta}-TrCP promotes glycogen synthase kinase 3-dependent degradation of the Nrf2 transcription factor in a Keap1-independent manner. Mol. Cell. Biol. 2011, 31, 1121–1133. [Google Scholar] [CrossRef]
- Chowdhry, S.; Zhang, Y.; McMahon, M.; Sutherland, C.; Cuadrado, A.; Hayes, J.D. Nrf2 is controlled by two distinct beta-TrCP recognition motifs in its Neh6 domain, one of which can be modulated by GSK-3 activity. Oncogene 2013, 32, 3765–3781. [Google Scholar] [CrossRef] [Green Version]
- Cuadrado, A. Structural and functional characterization of Nrf2 degradation by glycogen synthase kinase 3/beta-TrCP. Free Radic. Biol. Med. 2015, 88, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Rojo, A.I.; Medina-Campos, O.N.; Rada, P.; Zuniga-Toala, A.; Lopez-Gazcon, A.; Espada, S.; Pedraza-Chaverri, J.; Cuadrado, A. Signaling pathways activated by the phytochemical nordihydroguaiaretic acid contribute to a Keap1-independent regulation of Nrf2 stability: Role of glycogen synthase kinase-3. Free Radic. Biol. Med. 2012, 52, 473–487. [Google Scholar] [CrossRef] [PubMed]
- Moi, P.; Chan, K.; Asunis, I.; Cao, A.; Kan, Y.W. Isolation of NF-E2-related factor 2 (Nrf2), a NF-E2-like basic leucine zipper transcriptional activator that binds to the tandem NF-E2/AP1 repeat of the beta-globin locus control region. Proc. Natl. Acad. Sci. USA 1994, 91, 9926–9930. [Google Scholar] [CrossRef]
- Itoh, K.; Chiba, T.; Takahashi, S.; Ishii, T.; Igarashi, K.; Katoh, Y.; Oyake, T.; Hayashi, N.; Satoh, K.; Hatayama, I.; et al. An Nrf2/small Maf heterodimer mediates the induction of phase II detoxifying enzyme genes through antioxidant response elements. Biochem. Biophys. Res. Commun. 1997, 236, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Jain, M.R.; Chen, C.; Yue, X.; Hebbar, V.; Zhou, R.; Kong, A.N. Nrf2 Possesses a redox-insensitive nuclear export signal overlapping with the leucine zipper motif. J. Biol. Chem. 2005, 280, 28430–28438. [Google Scholar] [CrossRef]
- Jain, A.K.; Bloom, D.A.; Jaiswal, A.K. Nuclear import and export signals in control of Nrf2. J. Biol. Chem. 2005, 280, 29158–29168. [Google Scholar] [CrossRef] [PubMed]
- Nioi, P.; Nguyen, T.; Sherratt, P.J.; Pickett, C.B. The carboxy-terminal Neh3 domain of Nrf2 is required for transcriptional activation. Mol. Cell. Biol. 2005, 25, 10895–10906. [Google Scholar] [CrossRef]
- Miao, W.; Hu, L.; Scrivens, P.J.; Batist, G. Transcriptional regulation of NF-E2 p45-related factor (NRF2) expression by the aryl hydrocarbon receptor-xenobiotic response element signaling pathway: Direct cross-talk between phase I and II drug-metabolizing enzymes. J. Biol. Chem. 2005, 280, 20340–20348. [Google Scholar] [CrossRef]
- Kohle, C.; Bock, K.W. Coordinate regulation of Phase I and II xenobiotic metabolisms by the Ah receptor and Nrf2. Biochem. Pharmacol. 2007, 73, 1853–1862. [Google Scholar] [CrossRef]
- Wakabayashi, N.; Skoko, J.J.; Chartoumpekis, D.V.; Kimura, S.; Slocum, S.L.; Noda, K.; Palliyaguru, D.L.; Fujimuro, M.; Boley, P.A.; Tanaka, Y.; et al. Notch-Nrf2 axis: Regulation of Nrf2 gene expression and cytoprotection by notch signaling. Mol. Cell. Biol. 2014, 34, 653–663. [Google Scholar] [CrossRef] [Green Version]
- Yazaki, K.; Matsuno, Y.; Yoshida, K.; Sherpa, M.; Nakajima, M.; Matsuyama, M.; Kiwamoto, T.; Morishima, Y.; Ishii, Y.; Hizawa, N. ROS-Nrf2 pathway mediates the development of TGF-beta1-induced epithelial-mesenchymal transition through the activation of Notch signaling. Eur. J. Cell Biol. 2021, 100, 151181. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gordon, G.B. A strategy for cancer prevention: Stimulation of the Nrf2-ARE signaling pathway. Mol. Cancer Ther. 2004, 3, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.M.; Maltagliati, A.J. Nrf2 at the heart of oxidative stress and cardiac protection. Physiol. Genom. 2018, 50, 77–97. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Gong, J.; Thimmulappa, R.K.; Kosmider, B.; Biswal, S.; Duh, E.J. Nrf2 acts cell-autonomously in endothelium to regulate tip cell formation and vascular branching. Proc. Natl. Acad. Sci. USA 2013, 110, E3910–E3918. [Google Scholar] [CrossRef]
- Baeyens, N.; Bandyopadhyay, C.; Coon, B.G.; Yun, S.; Schwartz, M.A. Endothelial fluid shear stress sensing in vascular health and disease. J. Clin. Investig. 2016, 126, 821–828. [Google Scholar] [CrossRef]
- Chiu, J.J.; Chien, S. Effects of disturbed flow on vascular endothelium: Pathophysiological basis and clinical perspectives. Physiol. Rev. 2011, 91, 327–387. [Google Scholar] [CrossRef]
- Niu, N.; Xu, S.; Xu, Y.; Little, P.J.; Jin, Z.G. Targeting Mechanosensitive Transcription Factors in Atherosclerosis. Trends Pharmacol. Sci. 2019, 40, 253–266. [Google Scholar] [CrossRef]
- Takabe, W.; Warabi, E.; Noguchi, N. Anti-atherogenic effect of laminar shear stress via Nrf2 activation. Antioxid. Redox Signal. 2011, 15, 1415–1426. [Google Scholar] [CrossRef]
- Nigro, P.; Abe, J.; Berk, B.C. Flow shear stress and atherosclerosis: A matter of site specificity. Antioxid. Redox Signal. 2011, 15, 1405–1414. [Google Scholar] [CrossRef]
- McSweeney, S.R.; Warabi, E.; Siow, R.C. Nrf2 as an Endothelial Mechanosensitive Transcription Factor: Going with the Flow. Hypertension 2016, 67, 20–29. [Google Scholar] [CrossRef]
- Chen, X.L.; Varner, S.E.; Rao, A.S.; Grey, J.Y.; Thomas, S.; Cook, C.K.; Wasserman, M.A.; Medford, R.M.; Jaiswal, A.K.; Kunsch, C. Laminar flow induction of antioxidant response element-mediated genes in endothelial cells. A novel anti-inflammatory mechanism. J. Biol. Chem. 2003, 278, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Dai, G.; Vaughn, S.; Zhang, Y.; Wang, E.T.; Garcia-Cardena, G.; Gimbrone, M.A., Jr. Biomechanical forces in atherosclerosis-resistant vascular regions regulate endothelial redox balance via phosphoinositol 3-kinase/Akt-dependent activation of Nrf2. Circ. Res. 2007, 101, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Hosoya, T.; Maruyama, A.; Kang, M.I.; Kawatani, Y.; Shibata, T.; Uchida, K.; Warabi, E.; Noguchi, N.; Itoh, K.; Yamamoto, M. Differential responses of the Nrf2-Keap1 system to laminar and oscillatory shear stresses in endothelial cells. J. Biol. Chem. 2005, 280, 27244–27250. [Google Scholar] [CrossRef]
- Cheng, X.; Siow, R.C.; Mann, G.E. Impaired redox signaling and antioxidant gene expression in endothelial cells in diabetes: A role for mitochondria and the nuclear factor-E2-related factor 2-Kelch-like ECH-associated protein 1 defense pathway. Antioxid. Redox Signal. 2011, 14, 469–487. [Google Scholar] [CrossRef] [PubMed]
- Zakkar, M.; Van der Heiden, K.; Luong le, A.; Chaudhury, H.; Cuhlmann, S.; Hamdulay, S.S.; Krams, R.; Edirisinghe, I.; Rahman, I.; Carlsen, H.; et al. Activation of Nrf2 in endothelial cells protects arteries from exhibiting a proinflammatory state. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1851–1857. [Google Scholar] [CrossRef] [PubMed]
- Mylroie, H.; Dumont, O.; Bauer, A.; Thornton, C.C.; Mackey, J.; Calay, D.; Hamdulay, S.S.; Choo, J.R.; Boyle, J.J.; Samarel, A.M.; et al. PKCepsilon-CREB-Nrf2 signalling induces HO-1 in the vascular endothelium and enhances resistance to inflammation and apoptosis. Cardiovasc. Res. 2015, 106, 509–519. [Google Scholar] [CrossRef]
- Xiao, Y.; Xia, J.; Wu, S.; Lv, Z.; Huang, S.; Huang, H.; Su, X.; Cheng, J.; Ke, Y. Curcumin Inhibits Acute Vascular Inflammation through the Activation of Heme Oxygenase-1. Oxid. Med. Cell. Longev. 2018, 2018, 3295807. [Google Scholar] [CrossRef]
- Li, P.; Guo, X.; Lei, P.; Shi, S.; Luo, S.; Cheng, X. PI3K/Akt/uncoupling protein 2 signaling pathway may be involved in cell senescence and apoptosis induced by angiotensin II in human vascular endothelial cells. Mol. Biol. Rep. 2014, 41, 6931–6937. [Google Scholar] [CrossRef]
- Bendall, J.K.; Rinze, R.; Adlam, D.; Tatham, A.L.; de Bono, J.; Wilson, N.; Volpi, E.; Channon, K.M. Endothelial Nox2 overexpression potentiates vascular oxidative stress and hemodynamic response to angiotensin II: Studies in endothelial-targeted Nox2 transgenic mice. Circ. Res. 2007, 100, 1016–1025. [Google Scholar] [CrossRef]
- Yang, Y.; Tian, T.; Wang, Y.; Li, Z.; Xing, K.; Tian, G. SIRT6 protects vascular endothelial cells from angiotensin II-induced apoptosis and oxidative stress by promoting the activation of Nrf2/ARE signaling. Eur. J. Pharmacol. 2019, 859, 172516. [Google Scholar] [CrossRef]
- Iizuka, T.; Ishii, Y.; Itoh, K.; Kiwamoto, T.; Kimura, T.; Matsuno, Y.; Morishima, Y.; Hegab, A.E.; Homma, S.; Nomura, A.; et al. Nrf2-deficient mice are highly susceptible to cigarette smoke-induced emphysema. Genes Cells 2005, 10, 1113–1125. [Google Scholar] [CrossRef] [PubMed]
- Rangasamy, T.; Cho, C.Y.; Thimmulappa, R.K.; Zhen, L.; Srisuma, S.S.; Kensler, T.W.; Yamamoto, M.; Petrache, I.; Tuder, R.M.; Biswal, S. Genetic ablation of Nrf2 enhances susceptibility to cigarette smoke-induced emphysema in mice. J. Clin. Investig. 2004, 114, 1248–1259. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Rangasamy, T.; Thimmulappa, R.K.; Lee, H.; Osburn, W.O.; Brigelius-Flohe, R.; Kensler, T.W.; Yamamoto, M.; Biswal, S. Glutathione peroxidase 2, the major cigarette smoke-inducible isoform of GPX in lungs, is regulated by Nrf2. Am. J. Respir. Cell. Mol. Biol. 2006, 35, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.L.; Wu, Y.L.; Tang, G.J.; Lee, T.S.; Kou, Y.R. Ginkgo biloba extract confers protection from cigarette smoke extract-induced apoptosis in human lung endothelial cells: Role of heme oxygenase-1. Pulm. Pharmacol. Ther. 2009, 22, 286–296. [Google Scholar] [CrossRef]
- Zhao, Z.; Wang, X.; Zhang, R.; Ma, B.; Niu, S.; Di, X.; Ni, L.; Liu, C. Melatonin attenuates smoking-induced atherosclerosis by activating the Nrf2 pathway via NLRP3 inflammasomes in endothelial cells. Aging (Albany NY) 2021, 13, 11363–11380. [Google Scholar] [CrossRef]
- Prasad, S.; Sajja, R.K.; Kaisar, M.A.; Park, J.H.; Villalba, H.; Liles, T.; Abbruscato, T.; Cucullo, L. Role of Nrf2 and protective effects of Metformin against tobacco smoke-induced cerebrovascular toxicity. Redox. Biol. 2017, 12, 58–69. [Google Scholar] [CrossRef]
- Li, T.; Wang, H.; Ding, Y.; Zhou, M.; Zhou, X.; Zhang, X.; Ding, K.; He, J.; Lu, X.; Xu, J.; et al. Genetic elimination of Nrf2 aggravates secondary complications except for vasospasm after experimental subarachnoid hemorrhage in mice. Brain Res. 2014, 1558, 90–99. [Google Scholar] [CrossRef]
- Li, M.; Liu, X.; He, Y.; Zheng, Q.; Wang, M.; Wu, Y.; Zhang, Y.; Wang, C. Celastrol attenuates angiotensin II mediated human umbilical vein endothelial cells damage through activation of Nrf2/ERK1/2/Nox2 signal pathway. Eur. J. Pharmacol. 2017, 797, 124–133. [Google Scholar] [CrossRef]
- Zhou, X.; Liang, L.; Zhao, Y.; Zhang, H. Epigallocatechin-3-Gallate Ameliorates Angiotensin II-Induced Oxidative Stress and Apoptosis in Human Umbilical Vein Endothelial Cells through the Activation of Nrf2/Caspase-3 Signaling. J. Vasc. Res. 2017, 54, 299–308. [Google Scholar] [CrossRef]
- Han, J.; Shi, X.; Du, Y.; Shi, F.; Zhang, B.; Zheng, Z.; Xu, J.; Jiang, L. Schisandrin C targets Keap1 and attenuates oxidative stress by activating Nrf2 pathway in Ang II-challenged vascular endothelium. Phytother. Res. 2019, 33, 779–790. [Google Scholar] [CrossRef]
- Tao, L.; Gu, X.; Xu, E.; Ren, S.; Zhang, L.; Liu, W.; Lin, X.; Yang, J.; Chen, C. Osthole protects against Ang II-induced endotheliocyte death by targeting NF-kappaB pathway and Keap-1/Nrf2 pathway. Am. J. Transl. Res. 2019, 11, 142–159. [Google Scholar] [PubMed]
- Chen, J.; Gong, F.; Chen, M.F.; Li, C.; Hong, P.; Sun, S.; Zhou, C.; Qian, Z.J. In Vitro Vascular-Protective Effects of a Tilapia By-Product Oligopeptide on Angiotensin II-Induced Hypertensive Endothelial Injury in HUVEC by Nrf2/NF-kappaB Pathways. Mar. Drugs 2019, 17, 431. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Li, Q.; Mao, S.; Qin, L.; Dong, P. The protective effects of memantine against inflammation and impairment of endothelial tube formation induced by oxygen-glucose deprivation/reperfusion. Aging (Albany NY) 2020, 12, 21469–21480. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.C.; Lai, Y.N.; Wang, W.H.; Xu, X.M.; Li, X.Q.; Wang, H.; Zheng, J.Y.; Zheng, J.Q. Metformin Ameliorates Gestational Diabetes Mellitus-Induced Endothelial Dysfunction via Downregulation of p65 and Upregulation of Nrf2. Front. Pharmacol. 2020, 11, 575390. [Google Scholar] [CrossRef] [PubMed]
- Gjyshi, O.; Flaherty, S.; Veettil, M.V.; Johnson, K.E.; Chandran, B.; Bottero, V. Kaposi’s sarcoma-associated herpesvirus induces Nrf2 activation in latently infected endothelial cells through SQSTM1 phosphorylation and interaction with polyubiquitinated Keap1. J. Virol. 2015, 89, 2268–2286. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Liao, Q.; Luo, M.; Hua, C.; Zhao, J.; Yu, G.; Chen, X.; Li, X.; Zhang, X.; Ran, R.; et al. Cigarette smoke-induced oxidative stress activates NRF2 to mediate fibronectin disorganization in vascular formation. Open Biol. 2022, 12, 210310. [Google Scholar] [CrossRef]
- Zhu, Z.; Li, J.; Zhang, X. Astragaloside IV Protects Against Oxidized Low-Density Lipoprotein (ox-LDL)-Induced Endothelial Cell Injury by Reducing Oxidative Stress and Inflammation. Med. Sci. Monit. 2019, 25, 2132–2140. [Google Scholar] [CrossRef]
- Guo, J.; Ma, J.; Cai, K.; Chen, H.; Xie, K.; Xu, B.; Quan, D.; Du, J. Isoflavones from Semen Sojae Preparatum Improve Atherosclerosis and Oxidative Stress by Modulating Nrf2 Signaling Pathway through Estrogen-Like Effects. Evid. Based. Complement. Altern. Med. 2022, 2022, 4242099. [Google Scholar] [CrossRef]
- Hou, J.R.; Wang, Y.H.; Zhong, Y.N.; Che, T.T.; Hu, Y.; Bao, J.; Meng, N. Protective Effect of Flavonoids from a Deep-Sea-Derived Arthrinium sp. against ox-LDL-Induced Oxidative Injury through Activating the AKT/Nrf2/HO-1 Pathway in Vascular Endothelial Cells. Mar. Drugs 2021, 19, 712. [Google Scholar] [CrossRef]
- He, L.; Liu, Y.Y.; Wang, K.; Li, C.; Zhang, W.; Li, Z.Z.; Huang, X.Z.; Xiong, Y. Tanshinone IIA protects human coronary artery endothelial cells from ferroptosis by activating the NRF2 pathway. Biochem. Biophys. Res. Commun. 2021, 575, 1–7. [Google Scholar] [CrossRef]
- Chen, L.W.; Tsai, M.C.; Chern, C.Y.; Tsao, T.P.; Lin, F.Y.; Chen, S.J.; Tsui, P.F.; Liu, Y.W.; Lu, H.J.; Wu, W.L.; et al. A chalcone derivative, 1m-6, exhibits atheroprotective effects by increasing cholesterol efflux and reducing inflammation-induced endothelial dysfunction. Br. J. Pharmacol. 2020, 177, 5375–5392. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Li, M.; Lu, Y.; Li, J.; Ke, Y.; Yang, J. Ilexgenin A inhibits mitochondrial fission and promote Drp1 degradation by Nrf2-induced PSMB5 in endothelial cells. Drug Dev. Res. 2019, 80, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Song, F.; Li, Y.; Li, J.; Cui, Y.; Hong, Y.; Han, W.; Wu, W.; Lakhani, I.; Li, G.; et al. Acacetin exerts antioxidant potential against atherosclerosis through Nrf2 pathway in ApoE−/− Mice. J. Cell. Mol. Med. 2021, 25, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Xu, Y.; Jiang, L. Irisin attenuates oxidized low-density lipoprotein impaired angiogenesis through AKT/mTOR/S6K1/Nrf2 pathway. J. Cell. Physiol. 2019, 234, 18951–18962. [Google Scholar] [CrossRef]
- Pei, Y.; Lui, Y.; Cai, S.; Zhou, C.; Hong, P.; Qian, Z.J. A Novel Peptide Isolated from Microalgae Isochrysis zhanjiangensis Exhibits Anti-apoptosis and Anti-inflammation in Ox-LDL Induced HUVEC to Improve Atherosclerosis. Plant Foods Hum. Nutr. 2022, 77, 181–189. [Google Scholar] [CrossRef]
- Feng, Z.; Wang, C.; Yue, J.; Meng, Q.; Wu, J.; Sun, H. Kaempferol-induced GPER upregulation attenuates atherosclerosis via the PI3K/AKT/Nrf2 pathway. Pharm. Biol. 2021, 59, 1106–1116. [Google Scholar] [CrossRef]
- Yan, R.; Yan, J.; Chen, X.; Yu, Y.; Sun, T. Xanthoangelol Prevents Ox-LDL-Induced Endothelial Cell Injury by Activating Nrf2/ARE Signaling. J. Cardiovasc. Pharmacol. 2019, 74, 162–171. [Google Scholar] [CrossRef]
- Zhang, T.; Hu, Q.; Shi, L.; Qin, L.; Zhang, Q.; Mi, M. Equol Attenuates Atherosclerosis in Apolipoprotein E-Deficient Mice by Inhibiting Endoplasmic Reticulum Stress via Activation of Nrf2 in Endothelial Cells. PLoS ONE 2016, 11, e0167020. [Google Scholar] [CrossRef]
- Rajendran, P.; Alzahrani, A.M.; Ahmed, E.A.; Veeraraghavan, V.P. Kirenol Inhibits B[a]P-Induced Oxidative Stress and Apoptosis in Endothelial Cells via Modulation of the Nrf2 Signaling Pathway. Oxid. Med. Cell. Longev. 2021, 2021, 5585303. [Google Scholar] [CrossRef]
- Seo, Y.; Park, J.; Choi, W.; Ju Son, D.; Sung Kim, Y.; Kim, M.K.; Yoon, B.E.; Pyee, J.; Tae Hong, J.; Go, Y.M.; et al. Antiatherogenic Effect of Resveratrol Attributed to Decreased Expression of ICAM-1 (Intercellular Adhesion Molecule-1). Arterioscler. Thromb. Vasc. Biol. 2019, 39, 675–684. [Google Scholar] [CrossRef] [Green Version]
- Mao, H.; Tao, T.; Wang, X.; Liu, M.; Song, D.; Liu, X.; Shi, D. Zedoarondiol Attenuates Endothelial Cells Injury Induced by Oxidized Low-Density Lipoprotein via Nrf2 Activation. Cell. Physiol. Biochem. 2018, 48, 1468–1479. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Zhang, B.; Zhou, K.; Chen, M.; Wang, M.; Jia, Y.; Song, Y.; Li, Y.; Wen, A. Dietary ellagic acid improves oxidant-induced endothelial dysfunction and atherosclerosis: Role of Nrf2 activation. Int. J. Cardiol. 2014, 175, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Seo, M.; Lee, E.J. Salvianolic acid B inhibits atherogenesis of vascular cells through induction of Nrf2-dependent heme oxygenase-1. Curr. Med. Chem. 2014, 21, 3095–3106. [Google Scholar] [CrossRef]
- Luo, Y.; Lu, S.; Dong, X.; Xu, L.; Sun, G.; Sun, X. Dihydromyricetin protects human umbilical vein endothelial cells from injury through ERK and Akt mediated Nrf2/HO-1 signaling pathway. Apoptosis 2017, 22, 1013–1024. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, H.; Li, X.; Jia, B.; Yang, Y.; Zhou, P.; Li, P.; Chen, J. Miltirone protects human EA.hy926 endothelial cells from oxidized low-density lipoprotein-derived oxidative stress via a heme oxygenase-1 and MAPK/Nrf2 dependent pathway. Phytomedicine 2016, 23, 1806–1813. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zhang, Y.; Huang, X.; Xie, Y.; Qu, Y.; Long, H.; Gu, N.; Jiang, W. Z-Ligustilide protects vascular endothelial cells from oxidative stress and rescues high fat diet-induced atherosclerosis by activating multiple NRF2 downstream genes. Atherosclerosis 2019, 284, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.S.; Yoon, J.J.; Han, B.H.; Jeong, D.H.; Lee, Y.J.; Kang, D.G.; Lee, H.S. Ligustilide attenuates vascular inflammation and activates Nrf2/HO-1 induction and, NO synthesis in HUVECs. Phytomedicine 2018, 38, 12–23. [Google Scholar] [CrossRef]
- Zhang, S.; Jin, S.; Zhang, S.; Li, Y.Y.; Wang, H.; Chen, Y.; Lu, H. Vitexin protects against high glucose-induced endothelial cell apoptosis and oxidative stress via Wnt/beta-catenin and Nrf2 signalling pathway. Arch. Physiol. Biochem. 2022, 7, 1–10. [Google Scholar] [CrossRef]
- Zeng, J.; Deng, Z.; Zou, Y.; Liu, C.; Fu, H.; Gu, Y.; Chang, H. Theaflavin alleviates oxidative injury and atherosclerosis progress via activating microRNA-24-mediated Nrf2/HO-1 signal. Phytother. Res. 2021, 35, 3418–3427. [Google Scholar] [CrossRef]
- Chen, D.; Wu, Z.; Wu, L.N.; Jiang, J.; Hu, G.N. Theaflavin Attenuates TBHP-Induced Endothelial Cells Oxidative Stress by Activating PI3K/AKT/Nrf2 and Accelerates Wound Healing in Rats. Front. Bioeng. Biotechnol. 2022, 10, 830574. [Google Scholar] [CrossRef]
- Yan, C.; Zhang, X.; Miao, J.; Yuan, H.; Liu, E.; Liang, T.; Li, Q. Farrerol Directly Targets GSK-3beta to Activate Nrf2-ARE Pathway and Protect EA.hy926 Cells against Oxidative Stress-Induced Injuries. Oxid. Med. Cell. Longev. 2020, 2020, 5967434. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Liu, Y.; Liu, K.; Shu, Z.; Lv, T.; Chen, Z.; Feng, W.; Zhang, Y. A transcriptomics and molecular biology based investigation reveals the protective effect and mechanism of carnosol on t-BHP induced HRMECs via Nrf2 signaling pathway. Eur. J. Pharmacol. 2022, 923, 174933. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.Y.; Deng, L.H.; Huang, Z.J.; Daror, A.S.; Wang, Z.H.; Jin, W.J.; Zhuang, Z.; Tong, Q.; Zheng, G.Q.; Wang, Y. Buyang Huanwu Decoction Enhances Revascularization via Akt/GSK3beta/NRF2 Pathway in Diabetic Hindlimb Ischemia. Oxid. Med. Cell. Longev. 2021, 2021, 1470829. [Google Scholar] [CrossRef]
- Senthil, K.K.J.; Gokila, V.M.; Wang, S.Y. Activation of Nrf2-mediated anti-oxidant genes by antrodin C prevents hyperglycemia-induced senescence and apoptosis in human endothelial cells. Oncotarget 2017, 8, 96568–96587. [Google Scholar] [CrossRef] [PubMed]
- Li, C.L.; Liu, X.H.; Qiao, Y.; Ning, L.N.; Li, W.J.; Sun, Y.S.; Liu, D.S.; Gao, W.; Ma, C.M. Allicin alleviates inflammation of diabetic macroangiopathy via the Nrf2 and NF-kB pathway. Eur. J. Pharmacol. 2020, 876, 173052. [Google Scholar] [CrossRef]
- Chen, J.; Tan, L.; Li, C.; Zhou, C.; Hong, P.; Sun, S.; Qian, Z.J. Mechanism Analysis of a Novel Angiotensin-I-Converting Enzyme Inhibitory Peptide from Isochrysis zhanjiangensis Microalgae for Suppressing Vascular Injury in Human Umbilical Vein Endothelial Cells. J. Agric. Food Chem. 2020, 68, 4411–4423. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Teng, X.; Jin, S.; Dong, J.; Guo, Q.; Tian, D.; Wu, Y. Hydrogen sulfide improves endothelial dysfunction by inhibiting the vicious cycle of NLRP3 inflammasome and oxidative stress in spontaneously hypertensive rats. J. Hypertens. 2019, 37, 1633–1643. [Google Scholar] [CrossRef]
- Fratantonio, D.; Speciale, A.; Ferrari, D.; Cristani, M.; Saija, A.; Cimino, F. Palmitate-induced endothelial dysfunction is attenuated by cyanidin-3-O-glucoside through modulation of Nrf2/Bach1 and NF-kappaB pathways. Toxicol. Lett. 2015, 239, 152–160. [Google Scholar] [CrossRef]
- Hu, S.; Wu, Y.; Zhao, B.; Hu, H.; Zhu, B.; Sun, Z.; Li, P.; Du, S. Panax notoginseng Saponins Protect Cerebral Microvascular Endothelial Cells against Oxygen-Glucose Deprivation/Reperfusion-Induced Barrier Dysfunction via Activation of PI3K/Akt/Nrf2 Antioxidant Signaling Pathway. Molecules 2018, 23, 2781. [Google Scholar] [CrossRef]
- Wang, X.; Chen, L.; Wang, T.; Jiang, X.; Zhang, H.; Li, P.; Lv, B.; Gao, X. Ginsenoside Rg3 antagonizes adriamycin-induced cardiotoxicity by improving endothelial dysfunction from oxidative stress via upregulating the Nrf2-ARE pathway through the activation of akt. Phytomedicine 2015, 22, 875–884. [Google Scholar] [CrossRef]
- Liu, L.; Wang, R.; Xu, R.; Chu, Y.; Gu, W. Procyanidin B2 ameliorates endothelial dysfunction and impaired angiogenesis via the Nrf2/PPARgamma/sFlt-1 axis in preeclampsia. Pharmacol. Res. 2022, 177, 106127. [Google Scholar] [CrossRef] [PubMed]
- Saji, N.; Francis, N.; Blanchard, C.L.; Schwarz, L.J.; Santhakumar, A.B. Rice Bran Phenolic Compounds Regulate Genes Associated with Antioxidant and Anti-Inflammatory Activity in Human Umbilical Vein Endothelial Cells with Induced Oxidative Stress. Int. J. Mol. Sci. 2019, 20, 4715. [Google Scholar] [CrossRef] [PubMed]
- Hada, Y.; Uchida, H.A.; Otaka, N.; Onishi, Y.; Okamoto, S.; Nishiwaki, M.; Takemoto, R.; Takeuchi, H.; Wada, J. The Protective Effect of Chlorogenic Acid on Vascular Senescence via the Nrf2/HO-1 Pathway. Int. J. Mol. Sci. 2020, 21, 4527. [Google Scholar] [CrossRef]
- Chen, Z.W.; Miu, H.F.; Wang, H.P.; Wu, Z.N.; Wang, W.J.; Ling, Y.J.; Xu, X.H.; Sun, H.J.; Jiang, X. Pterostilbene protects against uraemia serum-induced endothelial cell damage via activation of Keap1/Nrf2/HO-1 signaling. Int. Urol. Nephrol. 2018, 50, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Zheng, Y.; Hou, X. Lipoxin A4 restores oxidative stress-induced vascular endothelial cell injury and thrombosis-related factor expression by its receptor-mediated activation of Nrf2-HO-1 axis. Cell. Signal. 2019, 60, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Ling, K.; Xu, A.; Chen, Y.; Chen, X.; Li, Y.; Wang, W. Protective effect of a hydrogen sulfide donor on balloon injury-induced restenosis via the Nrf2/HIF-1alpha signaling pathway. Int. J. Mol. Med. 2019, 43, 1299–1310. [Google Scholar] [CrossRef]
- Jiang, J.; Dong, C.; Zhai, L.; Lou, J.; Jin, J.; Cheng, S.; Chen, Z.; Guo, X.; Lin, D.; Ding, J.; et al. Paeoniflorin Suppresses TBHP-Induced Oxidative Stress and Apoptosis in Human Umbilical Vein Endothelial Cells via the Nrf2/HO-1 Signaling Pathway and Improves Skin Flap Survival. Front. Pharmacol. 2021, 12, 735530. [Google Scholar] [CrossRef]
- Zhang, Q.; Xue, T.; Guan, J.; Wang, W.; Shi, J.; Lu, J.; Jiang, X. Irigenin alleviates angiotensin II-induced oxidative stress and apoptosis in HUVEC cells by activating Nrf2 pathway. Drug Dev. Res. 2021, 82, 999–1007. [Google Scholar] [CrossRef]
- Kim, N.Y.; Trinh, N.T.; Ahn, S.G.; Kim, S.A. Cinnamaldehyde protects against oxidative stress and inhibits the TNFalphainduced inflammatory response in human umbilical vein endothelial cells. Int. J. Mol. Med. 2020, 46, 449–457. [Google Scholar] [CrossRef]
- Priestley, J.R.C.; Fink, K.E.; McCord, J.M.; Lombard, J.H. NRF2 activation with Protandim attenuates salt-induced vascular dysfunction and microvascular rarefaction. Microcirculation 2019, 26, e12575. [Google Scholar] [CrossRef]
- Gao, F.; Li, J.M.; Xi, C.; Li, H.H.; Liu, Y.L.; Wang, Y.P.; Xuan, L.J. Magnesium lithospermate B protects the endothelium from inflammation-induced dysfunction through activation of Nrf2 pathway. Acta Pharmacol. Sin. 2019, 40, 867–878. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.S.; Huang, P.H.; Wang, C.H.; Lin, F.Y.; Tsai, H.Y.; Wu, T.C.; Lin, S.J.; Chen, J.W. Nrf-2 mediated heme oxygenase-1 expression, an antioxidant-independent mechanism, contributes to anti-atherogenesis and vascular protective effects of Ginkgo biloba extract. Atherosclerosis 2011, 214, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.; Chang, H.; Liu, X.; Wang, S.; Liu, H.; Xuan, H. Brazilian Green Propolis Inhibits Ox-LDL-Stimulated Oxidative Stress in Human Umbilical Vein Endothelial Cells Partly through PI3K/Akt/mTOR-Mediated Nrf2/HO-1 Pathway. Evid.-Based. Complement. Altern. Med. 2019, 2019, 5789574. [Google Scholar] [CrossRef] [PubMed]
- Jarisarapurin, W.; Sanrattana, W.; Chularojmontri, L.; Kunchana, K.; Wattanapitayakul, S.K. Antioxidant Properties of Unripe Carica papaya Fruit Extract and Its Protective Effects against Endothelial Oxidative Stress. Evid.-Based. Complement. Altern. Med. 2019, 2019, 4912631. [Google Scholar] [CrossRef]
- Sung, L.C.; Chao, H.H.; Chen, C.H.; Tsai, J.C.; Liu, J.C.; Hong, H.J.; Cheng, T.H.; Chen, J.J. Lycopene inhibits cyclic strain-induced endothelin-1 expression through the suppression of reactive oxygen species generation and induction of heme oxygenase-1 in human umbilical vein endothelial cells. Clin. Exp. Pharmacol. Physiol. 2015, 42, 632–639. [Google Scholar] [CrossRef]
- Oh, Y.; Ahn, C.B.; Je, J.Y. Cytoprotective Role of Edible Seahorse (Hippocampus abdominalis)-Derived Peptides in H2O2-Induced Oxidative Stress in Human Umbilical Vein Endothelial Cells. Mar. Drugs 2021, 19, 86. [Google Scholar] [CrossRef]
- Huang, M.Z.; Yang, Y.J.; Liu, X.W.; Qin, Z.; Li, J.Y. Aspirin eugenol ester attenuates oxidative injury of vascular endothelial cells by regulating NOS and Nrf2 signalling pathways. Br. J. Pharmacol. 2019, 176, 906–918. [Google Scholar] [CrossRef]
- Zhang, J.; Cai, W.; Fan, Z.; Yang, C.; Wang, W.; Xiong, M.; Ma, C.; Yang, J. MicroRNA-24 inhibits the oxidative stress induced by vascular injury by activating the Nrf2/Ho-1 signaling pathway. Atherosclerosis 2019, 290, 9–18. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, X.; Liu, X.; Du, A.; Yu, B. miR-200a mediates protection of thymosin beta-4 in cardiac microvascular endothelial cells as a novel mechanism under hypoxia-reoxygenation injury. J. Cell. Biochem. 2019, 120, 19098–19106. [Google Scholar] [CrossRef]
- Liu, Q.Q.; Ren, K.; Liu, S.H.; Li, W.M.; Huang, C.J.; Yang, X.H. MicroRNA-140-5p aggravates hypertension and oxidative stress of atherosclerosis via targeting Nrf2 and Sirt2. Int. J. Mol. Med. 2019, 43, 839–849. [Google Scholar] [CrossRef] [Green Version]
- Pan, H.; Xue, C.; Auerbach, B.J.; Fan, J.; Bashore, A.C.; Cui, J.; Yang, D.Y.; Trignano, S.B.; Liu, W.; Shi, J.; et al. Single-Cell Genomics Reveals a Novel Cell State During Smooth Muscle Cell Phenotypic Switching and Potential Therapeutic Targets for Atherosclerosis in Mouse and Human. Circulation 2020, 142, 2060–2075. [Google Scholar] [CrossRef] [PubMed]
- Herring, B.P.; Hoggatt, A.M.; Burlak, C.; Offermanns, S. Previously differentiated medial vascular smooth muscle cells contribute to neointima formation following vascular injury. Vasc. Cell. 2014, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhuang, W.; Xiong, T.; Park, W.S.; Zhang, S.; Zha, Y.; Yao, J.; Wang, F.; Yang, Y.; Chen, Y.; et al. Nrf2 deficiency attenuates atherosclerosis by reducing LOX-1-mediated proliferation and migration of vascular smooth muscle cells. Atherosclerosis 2022, 347, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Wang, C.; Zang, H.; Qi, L.; Azhar, M.; Nagarkatti, M.; Nagarkatti, P.; Cai, G.; Weiser-Evans, M.C.M.; Cui, T. Mature Vascular Smooth Muscle Cells, but Not Endothelial Cells, Serve as the Major Cellular Source of Intimal Hyperplasia in Vein Grafts. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 1870–1890. [Google Scholar] [CrossRef]
- Miano, J.M.; Fisher, E.A.; Majesky, M.W. Fate and State of Vascular Smooth Muscle Cells in Atherosclerosis. Circulation 2021, 143, 2110–2116. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Deng, J.; Yu, X.J.; Yang, S.; Yang, Y.; Zang, W.J. Activation of M3AChR (Type 3 Muscarinic Acetylcholine Receptor) and Nrf2 (Nuclear Factor Erythroid 2-Related Factor 2) Signaling by Choline Alleviates Vascular Smooth Muscle Cell Phenotypic Switching and Vascular Remodeling. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 2649–2664. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Li, X.X.; Wang, L.; Wang, M.; Zhang, Y.; Li, P.L. Contribution of Nrf2 to Atherogenic Phenotype Switching of Coronary Arterial Smooth Muscle Cells Lacking CD38 Gene. Cell. Physiol. Biochem. 2015, 37, 432–444. [Google Scholar] [CrossRef]
- Ashino, T.; Yamamoto, M.; Yoshida, T.; Numazawa, S. Redox-sensitive transcription factor Nrf2 regulates vascular smooth muscle cell migration and neointimal hyperplasia. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 760–768. [Google Scholar] [CrossRef]
- Chen, C.; Ma, J.; Xu, Z.; Chen, L.; Sun, B.; Shi, Y.; Miao, Y.; Wu, T.; Qin, M.; Zhang, Y.; et al. Rosmarinic Acid Inhibits Platelet Aggregation and Neointimal Hyperplasia In Vivo and Vascular Smooth Muscle Cell Dedifferentiation, Proliferation, and Migration In Vitro via Activation of the Keap1-Nrf2-ARE Antioxidant System. J. Agric. Food Chem. 2022, 70, 7420–7440. [Google Scholar] [CrossRef]
- Nie, X.; Shen, C.; Tan, J.; Yang, X.; Wang, W.; Dai, Y.; Sun, H.; Wu, Z.; Chen, J. Andrographolide Attenuates Established Pulmonary Hypertension via Rescue of Vascular Remodeling. Biomolecules 2021, 11, 1801. [Google Scholar] [CrossRef]
- Hwang, S.M.; Lee, Y.J.; Lee, Y.P.; Yoon, J.J.; Lee, S.M.; Cha, J.D.; Choi, K.M.; Kang, D.G.; Lee, H.S. Anti-Proliferative Effect of an Aqueous Extract of Prunella vulgaris in Vascular Smooth Muscle Cells. Evid.-Based. Complement. Altern. Med. 2013, 2013, 936463. [Google Scholar] [CrossRef] [PubMed]
- Buglak, N.E.; Jiang, W.; Bahnson, E.S.M. Cinnamic aldehyde inhibits vascular smooth muscle cell proliferation and neointimal hyperplasia in Zucker Diabetic Fatty rats. Redox Biol. 2018, 19, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Xu, Y.; Qiu, Z.; Jiang, L. Sulforaphane Attenuates Angiotensin II-Induced Vascular Smooth Muscle Cell Migration via Suppression of NOX4/ROS/Nrf2 Signaling. Int. J. Biol. Sci. 2019, 15, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Zghonda, N.; Yoshida, S.; Araki, M.; Kusunoki, M.; Mliki, A.; Ghorbel, A.; Miyazaki, H. Greater effectiveness of epsilon-viniferin in red wine than its monomer resveratrol for inhibiting vascular smooth muscle cell proliferation and migration. Biosci. Biotechnol. Biochem. 2011, 75, 1259–1267. [Google Scholar] [CrossRef]
- Kim, J.W.; Lim, S.C.; Lee, M.Y.; Lee, J.W.; Oh, W.K.; Kim, S.K.; Kang, K.W. Inhibition of neointimal formation by trans-resveratrol: Role of phosphatidyl inositol 3-kinase-dependent Nrf2 activation in heme oxygenase-1 induction. Mol. Nutr. Food Res. 2010, 54, 1497–1505. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Zhao, Y.; Feng, P.; Liu, Y. Sulfiredoxin-1 Inhibits PDGF-BB-Induced Vascular Smooth Muscle Cell Proliferation and Migration by Enhancing the Activation of Nrf2/ARE Signaling. Int. Heart J. 2022, 63, 113–121. [Google Scholar] [CrossRef]
- Qiu, L.; Hu, L.; Liu, X.; Li, W.; Zhang, X.; Xia, H.; Zhang, C. Physalin B inhibits PDGF-BB-induced VSMC proliferation, migration and phenotypic transformation by activating the Nrf2 pathway. Food Funct. 2021, 12, 10950–10966. [Google Scholar] [CrossRef]
- Choi, S.H.; Park, S.; Oh, C.J.; Leem, J.; Park, K.G.; Lee, I.K. Dipeptidyl peptidase-4 inhibition by gemigliptin prevents abnormal vascular remodeling via NF-E2-related factor 2 activation. Vascul. Pharmacol. 2015, 73, 11–19. [Google Scholar] [CrossRef]
- Kim, N.; Hwangbo, C.; Lee, S.; Lee, J.H. Eupatolide inhibits PDGF-induced proliferation and migration of aortic smooth muscle cells through ROS-dependent heme oxygenase-1 induction. Phytother. Res. 2013, 27, 1700–1707. [Google Scholar] [CrossRef]
- Lee, S.; Seo, J.; Ryoo, S.; Cuong, T.D.; Min, B.S.; Lee, J.H. Malabaricone C inhibits PDGF-induced proliferation and migration of aortic smooth muscle cells through induction of heme oxygenase-1. J. Cell. Biochem. 2012, 113, 2866–2876. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, H.; You, L.; Zhang, J.; Jiang, Z. Coenzyme Q10 inhibits intracranial aneurysm formation and progression in a mouse model. Pediatr. Res. 2022, 91, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Behnammanesh, G.; Durante, G.L.; Khanna, Y.P.; Peyton, K.J.; Durante, W. Canagliflozin inhibits vascular smooth muscle cell proliferation and migration: Role of heme oxygenase-1. Redox. Biol. 2020, 32, 101527. [Google Scholar] [CrossRef] [PubMed]
- Hwang, A.R.; Han, J.H.; Lim, J.H.; Kang, Y.J.; Woo, C.H. Fluvastatin inhibits AGE-induced cell proliferation and migration via an ERK5-dependent Nrf2 pathway in vascular smooth muscle cells. PLoS ONE 2017, 12, e0178278. [Google Scholar] [CrossRef] [PubMed]
- Aghagolzadeh, P.; Radpour, R.; Bachtler, M.; van Goor, H.; Smith, E.R.; Lister, A.; Odermatt, A.; Feelisch, M.; Pasch, A. Hydrogen sulfide attenuates calcification of vascular smooth muscle cells via KEAP1/NRF2/NQO1 activation. Atherosclerosis 2017, 265, 78–86. [Google Scholar] [CrossRef]
- Ma, W.Q.; Sun, X.J.; Zhu, Y.; Liu, N.F. Metformin attenuates hyperlipidaemia-associated vascular calcification through anti-ferroptotic effects. Free Radic. Biol. Med. 2021, 165, 229–242. [Google Scholar] [CrossRef]
- Cui, L.; Zhou, Q.; Zheng, X.; Sun, B.; Zhao, S. Mitoquinone attenuates vascular calcification by suppressing oxidative stress and reducing apoptosis of vascular smooth muscle cells via the Keap1/Nrf2 pathway. Free Radic. Biol. Med. 2020, 161, 23–31. [Google Scholar] [CrossRef]
- Chen, Y.; Huang, C.; Zhu, S.Y.; Zou, H.C.; Xu, C.Y.; Chen, Y.X. Overexpression of HOTAIR attenuates Pi-induced vascular calcification by inhibiting Wnt/beta-catenin through regulating miR-126/Klotho/SIRT1 axis. Mol. Cell. Biochem. 2021, 476, 3551–3561. [Google Scholar] [CrossRef]
- Ji, R.; Sun, H.; Peng, J.; Ma, X.; Bao, L.; Fu, Y.; Zhang, X.; Luo, C.; Gao, C.; Jin, Y.; et al. Rosmarinic acid exerts an antagonistic effect on vascular calcification by regulating the Nrf2 signalling pathway. Free Radic. Res. 2019, 53, 187–197. [Google Scholar] [CrossRef]
- Kim, J.Y.; Cho, H.J.; Sir, J.J.; Kim, B.K.; Hur, J.; Youn, S.W.; Yang, H.M.; Jun, S.I.; Park, K.W.; Hwang, S.J.; et al. Sulfasalazine induces haem oxygenase-1 via ROS-dependent Nrf2 signalling, leading to control of neointimal hyperplasia. Cardiovasc. Res. 2009, 82, 550–560. [Google Scholar] [CrossRef]
- Durham, A.L.; Speer, M.Y.; Scatena, M.; Giachelli, C.M.; Shanahan, C.M. Role of smooth muscle cells in vascular calcification: Implications in atherosclerosis and arterial stiffness. Cardiovasc. Res. 2018, 114, 590–600. [Google Scholar] [CrossRef]
- Jin, D.; Lin, L.; Xie, Y.; Jia, M.; Qiu, H.; Xun, K. NRF2-suppressed vascular calcification by regulating the antioxidant pathway in chronic kidney disease. FASEB J. 2022, 36, e22098. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Enaka, M.; Muragaki, Y. Activation of KEAP1/NRF2/P62 signaling alleviates high phosphate-induced calcification of vascular smooth muscle cells by suppressing reactive oxygen species production. Sci. Rep. 2019, 9, 10366. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.H.; Du, Y.; Sheng, Z.; Li, Y.; Qiu, X.; Tian, B.; Yao, L. OGT-Mediated KEAP1 Glycosylation Accelerates NRF2 Degradation Leading to High Phosphate-Induced Vascular Calcification in Chronic Kidney Disease. Front. Physiol. 2020, 11, 1092. [Google Scholar] [CrossRef] [PubMed]
- Girona, J.; Rosales, R.; Saavedra, P.; Masana, L.; Vallve, J.C. Palmitate decreases migration and proliferation and increases oxidative stress and inflammation in smooth muscle cells: Role of the Nrf2 signaling pathway. Am. J. Physiol. Cell. Physiol. 2019, 316, C888–C897. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Pineiro, J.A.; Gonzalez-Rovira, A.; Sanchez-Gomar, I.; Moreno, J.A.; Duran-Ruiz, M.C. Nrf2 and Heme Oxygenase-1 Involvement in Atherosclerosis Related Oxidative Stress. Antioxidants 2021, 10, 1463. [Google Scholar] [CrossRef]
- Ishii, T.; Itoh, K.; Ruiz, E.; Leake, D.S.; Unoki, H.; Yamamoto, M.; Mann, G.E. Role of Nrf2 in the regulation of CD36 and stress protein expression in murine macrophages: Activation by oxidatively modified LDL and 4-hydroxynonenal. Circ. Res. 2004, 94, 609–616. [Google Scholar] [CrossRef]
- Sussan, T.E.; Jun, J.; Thimmulappa, R.; Bedja, D.; Antero, M.; Gabrielson, K.L.; Polotsky, V.Y.; Biswal, S. Disruption of Nrf2, a key inducer of antioxidant defenses, attenuates ApoE-mediated atherosclerosis in mice. PLoS ONE 2008, 3, e3791. [Google Scholar] [CrossRef] [PubMed]
- Barajas, B.; Che, N.; Yin, F.; Rowshanrad, A.; Orozco, L.D.; Gong, K.W.; Wang, X.; Castellani, L.W.; Reue, K.; Lusis, A.J.; et al. NF-E2-related factor 2 promotes atherosclerosis by effects on plasma lipoproteins and cholesterol transport that overshadow antioxidant protection. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 58–66. [Google Scholar] [CrossRef]
- Freigang, S.; Ampenberger, F.; Spohn, G.; Heer, S.; Shamshiev, A.T.; Kisielow, J.; Hersberger, M.; Yamamoto, M.; Bachmann, M.F.; Kopf, M. Nrf2 is essential for cholesterol crystal-induced inflammasome activation and exacerbation of atherosclerosis. Eur. J. Immunol. 2011, 41, 2040–2051. [Google Scholar] [CrossRef]
- Harada, N.; Ito, K.; Hosoya, T.; Mimura, J.; Maruyama, A.; Noguchi, N.; Yagami, K.; Morito, N.; Takahashi, S.; Maher, J.M.; et al. Nrf2 in bone marrow-derived cells positively contributes to the advanced stage of atherosclerotic plaque formation. Free Radic. Biol. Med. 2012, 53, 2256–2262. [Google Scholar] [CrossRef]
- Liu, J.; Yang, B.; Wang, Y.; Wu, Y.; Fan, B.; Zhu, S.; Song, E.; Song, Y. Polychlorinated biphenyl quinone promotes macrophage polarization to CD163(+) cells through Nrf2 signaling pathway. Environ. Pollut. 2020, 257, 113587. [Google Scholar] [CrossRef] [PubMed]
- Bozaykut, P.; Karademir, B.; Yazgan, B.; Sozen, E.; Siow, R.C.; Mann, G.E.; Ozer, N.K. Effects of vitamin E on peroxisome proliferator-activated receptor gamma and nuclear factor-erythroid 2-related factor 2 in hypercholesterolemia-induced atherosclerosis. Free Radic. Biol. Med. 2014, 70, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Ruotsalainen, A.K.; Lappalainen, J.P.; Heiskanen, E.; Merentie, M.; Sihvola, V.; Napankangas, J.; Lottonen-Raikaslehto, L.; Kansanen, E.; Adinolfi, S.; Kaarniranta, K.; et al. Nuclear factor E2-related factor 2 deficiency impairs atherosclerotic lesion development but promotes features of plaque instability in hypercholesterolaemic mice. Cardiovasc. Res. 2019, 115, 243–254. [Google Scholar] [CrossRef]
- McCormick, M.L.; Gavrila, D.; Weintraub, N.L. Role of oxidative stress in the pathogenesis of abdominal aortic aneurysms. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Jreije, A.; Medlej-Hashim, M.; Hajal, J.; Saliba, Y.; Chacar, S.; Fares, N.; Khouzami, L. Calcitriol Supplementation Protects Against Apoptosis and Alleviates the Severity of Abdominal Aortic Aneurysm Induced by Angiotensin II and Anti-TGFβ. J. Cardiovasc. Transl. Res. 2022; ahead of print. [Google Scholar] [CrossRef]
- Qiu, R.; Chen, S.; Hua, F.; Bian, S.; Chen, J.; Li, G.; Wu, X. Betanin Prevents Experimental Abdominal Aortic Aneurysm Progression by Modulating the TLR4/NF-kappaB and Nrf2/HO-1 Pathways. Biol. Pharm. Bull. 2021, 44, 1254–1262. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Xu, T.; Feng, X.; Lai, Y.; Yang, Y.; Zheng, H.; He, X.; Wei, G.; Liao, W.; Liao, Y.; et al. Itaconate prevents abdominal aortic aneurysm formation through inhibiting inflammation via activation of Nrf2. EBioMedicine 2020, 57, 102832. [Google Scholar] [CrossRef] [PubMed]
- Xiao, T.; Zhang, L.; Huang, Y.; Shi, Y.; Wang, J.; Ji, Q.; Ye, J.; Lin, Y.; Liu, H. Sestrin2 increases in aortas and plasma from aortic dissection patients and alleviates angiotensin II-induced smooth muscle cell apoptosis via the Nrf2 pathway. Life Sci. 2019, 218, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.Y.; Vo, T.T.T.; Lee, C.W.; Chen, Y.L.; Lin, W.N.; Cheng, H.C.; Vo, Q.C.; Lee, I.T. Carbon monoxide releasing molecule-2 attenuates angiotensin II-induced IL-6/Jak2/Stat3-associated inflammation by inhibiting NADPH oxidase- and mitochondria-derived ROS in human aortic smooth muscle cells. Biochem. Pharmacol. 2022, 198, 114978. [Google Scholar] [CrossRef]
- Piechota-Polanczyk, A.; Kopacz, A.; Kloska, D.; Zagrapan, B.; Neumayer, C.; Grochot-Przeczek, A.; Huk, I.; Brostjan, C.; Dulak, J.; Jozkowicz, A. Simvastatin Treatment Upregulates HO-1 in Patients with Abdominal Aortic Aneurysm but Independently of Nrf2. Oxid. Med. Cell. Longev. 2018, 2018, 2028936. [Google Scholar] [CrossRef] [Green Version]
- Kopacz, A.; Werner, E.; Grochot-Przeczek, A.; Kloska, D.; Hajduk, K.; Neumayer, C.; Jozkowicz, A.; Piechota-Polanczyk, A. Simvastatin Attenuates Abdominal Aortic Aneurysm Formation Favoured by Lack of Nrf2 Transcriptional Activity. Oxid. Med. Cell. Longev. 2020, 2020, 6340190. [Google Scholar] [CrossRef]
- Pan, B.; Zhang, H.; Cui, T.; Wang, X. TFEB activation protects against cardiac proteotoxicity via increasing autophagic flux. J. Mol. Cell. Cardiol. 2017, 113, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.; Wang, B.; Li, W.; Wang, L.; Song, X.; Guo, C.; Li, Y.; Liu, F.; Zhu, F.; Wang, Q.; et al. Systemic application of 3-methyladenine markedly inhibited atherosclerotic lesion in ApoE−/− mice by modulating autophagy, foam cell formation and immune-negative molecules. Cell Death Dis. 2016, 7, e2498. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Popal, M.S.; Liu, Y.; Gao, R.; Lyu, S.; Chen, K.; Liu, Y. Ginkgo Biloba Leaf Extract Attenuates Atherosclerosis in Streptozotocin-Induced Diabetic ApoE−/− Mice by Inhibiting Endoplasmic Reticulum Stress via Restoration of Autophagy through the mTOR Signaling Pathway. Oxid. Med. Cell. Longev. 2019, 2019, 8134678. [Google Scholar] [CrossRef] [PubMed]
- de Zeeuw, D.; Akizawa, T.; Audhya, P.; Bakris, G.L.; Chin, M.; Christ-Schmidt, H.; Goldsberry, A.; Houser, M.; Krauth, M.; Lambers Heerspink, H.J.; et al. Bardoxolone methyl in type 2 diabetes and stage 4 chronic kidney disease. N. Engl. J. Med. 2013, 369, 2492–2503. [Google Scholar] [CrossRef] [PubMed]
- Mitsuishi, Y.; Taguchi, K.; Kawatani, Y.; Shibata, T.; Nukiwa, T.; Aburatani, H.; Yamamoto, M.; Motohashi, H. Nrf2 redirects glucose and glutamine into anabolic pathways in metabolic reprogramming. Cancer Cell 2012, 22, 66–79. [Google Scholar] [CrossRef] [PubMed]
- Robledinos-Anton, N.; Fernandez-Gines, R.; Manda, G.; Cuadrado, A. Activators and Inhibitors of NRF2: A Review of Their Potential for Clinical Development. Oxid. Med. Cell. Longev. 2019, 2019, 9372182. [Google Scholar] [CrossRef] [Green Version]
| Approach | Nrf2 Signaling | Finding | Reference |
|---|---|---|---|
| Compounds | Atherosclerosis (AS) Related | ||
| Salvianolic acid B | Nrf2/HO-1 | Inhibits PDGF-induced proliferation and migration of VSMCs Activates Nrf2-mediated HO-1 expression and suppression of PDGF-induced neointimal hyperplasia in cultured arterial rings isolated from mice ex vivo | [103] |
| Eupatolide | Nrf2/HO-1 | Inhibits PDGF-induced proliferation and migration of aortic SMCs in vitro | [159] |
| Compounds | Diabetes Related | ||
| Aqueous extract of Prunella vulgaris | Nrf2/HO-1 | Exhibits inhibitory effects on high glucose-stimulated VSMCs proliferation and migration, and invasion activities in vitro | [151] |
| Cinnamic aldehyde | Nrf2 | Inhibits neointimal hyperplasia after carotid artery balloon injury in the Zucker Diabetic Fatty (ZDF) rats and inhibits proliferation of ZDF VSMCs in vitro | [152] |
| Canagliflozin | Nrf2/HO-1 | Stimulates HO-1 expression in mice and human VSMCs through the ROS-Nrf2 pathway, and inhibits VSMCs proliferation and migration in vitro | [162] |
| Fluvastatin | ERK5/Nrf2 | Activates ERK5-dependent Nrf2 pathway and inhibits cellular proliferation and migration in VSMCs in vitro | [163] |
| Compounds | Vascular Calcification Related | ||
| Hydrogen sulfide | Keap1/Nrf2/NQO1 | Attenuates circulating calciprotein particles (CPP)-induced VSMCs calcification in vitro | [164] |
| Metformin | Nrf2 | Inhibits hyperlipidemia-associated calcium deposition in the rat aortic tissue of hyperlipidemia-related vascular calcification model in vivo and attenuates ferroptosis with increased calcium deposition in VSMCs with PA treatment | [165] |
| Mitoquinone | Keap1/Nrf2 | Attenuates vascular calcification by suppressing oxidative stress and reducing apoptosis in adenine-induced calcification in rats and inorganic phosphate-induced calcification in VSMCs in vitro | [166] |
| miR-126 | Sirt1/Nrf2 | Attenuates calcification, in human aortic smooth muscle cells (HASMCs) in vitro and in a mouse calcification model in vivo | [167] |
| Rosmarinic acid | Keap1/Nrf2/ARE | Inhibits VSMCs proliferation, migration, and calcification in a rat model of vascular calcification model induced by high-fat diet and vitamin D3 injection as well as β-glyerophosphate-induced calcification in rat aortic SMCs in vitro | [168] |
| Compounds | Vascular Occlusive Disease and Other Diseases Related | ||
| Rosmarinic acid | Keap1/Nrf2/ARE | Inhibits platelet aggregation and neointimal hyperplasia in vivo and VSMCs dedifferentiation, proliferation, and migration in vitro | [149] |
| Andrographolide | NOX/Nrf2 | Reverses pulmonary vascular remodeling through modulation of NOX/Nrf2-mediated oxidative stress and NF-κB-mediated inflammation in both chronic hypoxia and Sugen5416/hypoxia mouse pulmonary hypertension (PH) models and in cultured human PASMCs isolated from either healthy donors or PH patients | [150] |
| Sulforaphane | NOX4/ROS/Nrf2 | Attenuates Ang II-induced human VSMCs migration in vitro | [153] |
| Trans-resveratrol | Nrf2/HO-1 | Suppresses intimal hyperplasia in a mouse model of wire-injured femoral artery injury by oral administration Inhibits PDGF-stimulated DNA synthesis and cell proliferation in VSMCs in vitro | [155] |
| Sulfiredoxin-1 | Nrf2/ARE | Inhibits PDGF-BB-induced VSMCs proliferation and migration in vitro | [156] |
| Physalin B | Nrf2/HO-1 | Inhibits PDGF-BB-induced VSMCs proliferation, migration, and phenotypic transformation in vitro and prevents intimal hyperplasia in a mouse model of carotid artery injury induced by ligation | [157] |
| Gemigliptin | Nrf2/HO-1 | Exerts a preventative effect on ligation injury-induced neointimal hyperplasia in vivo and inhibits VSMCs proliferation and migration in vitro | [158] |
| Malabaricone C | Nrf2/HO-1 | Inhibits PDGF-induced proliferation and migration of aortic SMCs in vitro | [160] |
| Coenzyme Q10 | Nrf2/HO-1 | Attenuates intracranial aneurysm formation and rupture in mice and reduces H2O2-induced oxidative stress in VSMCs in vitro | [161] |
| Sulfasalazine | Nrf2/HO-1 | Suppresses VSMCs growth in vitro and prevents neointimal hyperplasia in rat carotid arteries induced by balloon in vivo | [169] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, W.; Hendrix, A.; Nair, S.; Cui, T. Nrf2-Mediated Dichotomy in the Vascular System: Mechanistic and Therapeutic Perspective. Cells 2022, 11, 3042. https://doi.org/10.3390/cells11193042
Wu W, Hendrix A, Nair S, Cui T. Nrf2-Mediated Dichotomy in the Vascular System: Mechanistic and Therapeutic Perspective. Cells. 2022; 11(19):3042. https://doi.org/10.3390/cells11193042
Chicago/Turabian StyleWu, Weiwei, Andrew Hendrix, Sharad Nair, and Taixing Cui. 2022. "Nrf2-Mediated Dichotomy in the Vascular System: Mechanistic and Therapeutic Perspective" Cells 11, no. 19: 3042. https://doi.org/10.3390/cells11193042