
The Flavone Cirsiliol from Salvia x jamensis Binds the F1 Moiety of ATP Synthase, Modulating Free Radical Production

 ,
,  , , , ,
, , , ,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. General Experimental Procedures
2.1.1. Plant Material
2.1.2. Extraction and Isolation of Cirsiliol
2.2. Extraction of Retinas
2.3. Purified Bovine Rod OS Preparations and Treatments
2.4. Transmission Electron Microscopy
2.5. ATP Synthesis Assay
2.6. Assay of the ETC Complexes
2.7. Cytofluorimetric Assay
2.8. Molecular Modeling
2.9. Statistical Analysis
3. Results
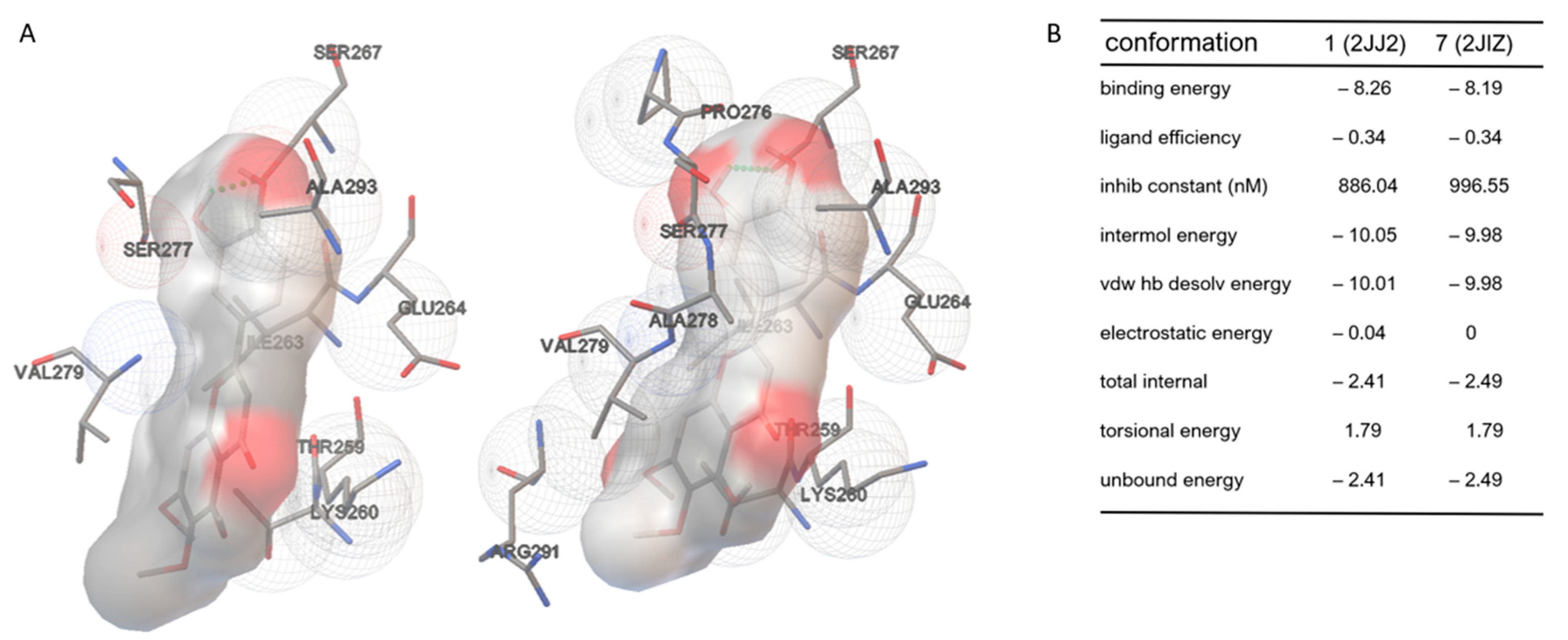
3.1. Cirsiliol Binds the F1Fo-ATP Synthase F1 Moiety
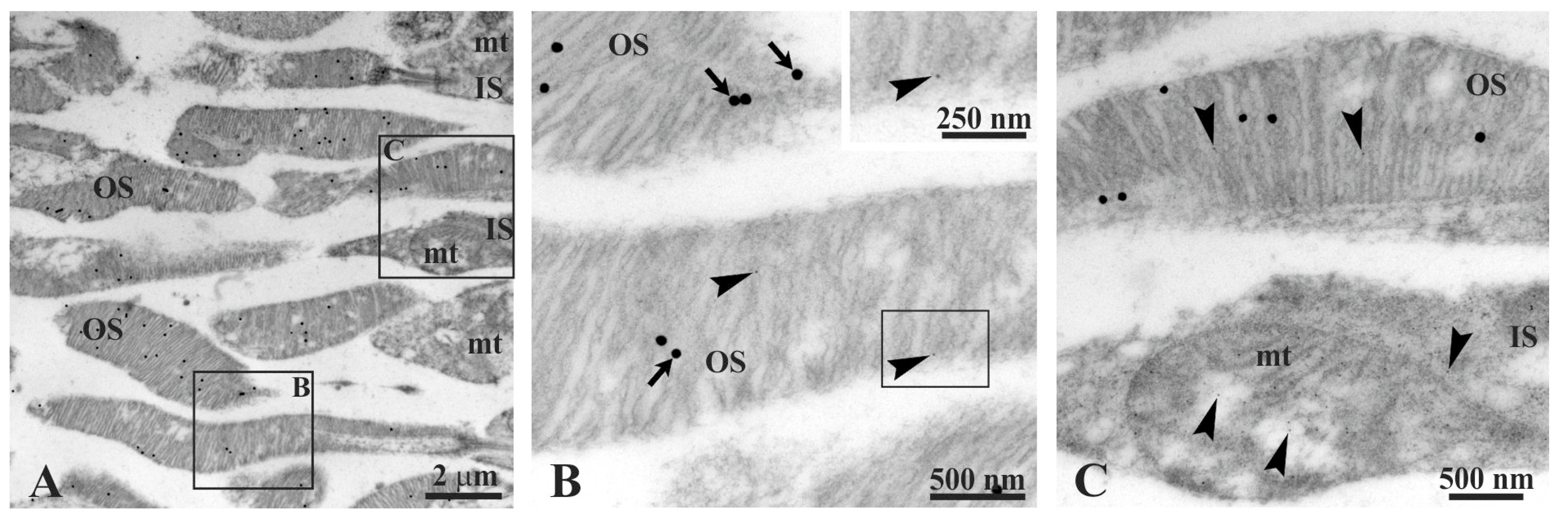
3.2. Rhodopsin and the β-Subunit of ATP Synthase Colocalize in the Rod Outer Segment
3.3. F1Fo-ATP Synthase Activity in Rod OS
3.4. Cirsiliol Inhibits OS ATP Synthesis
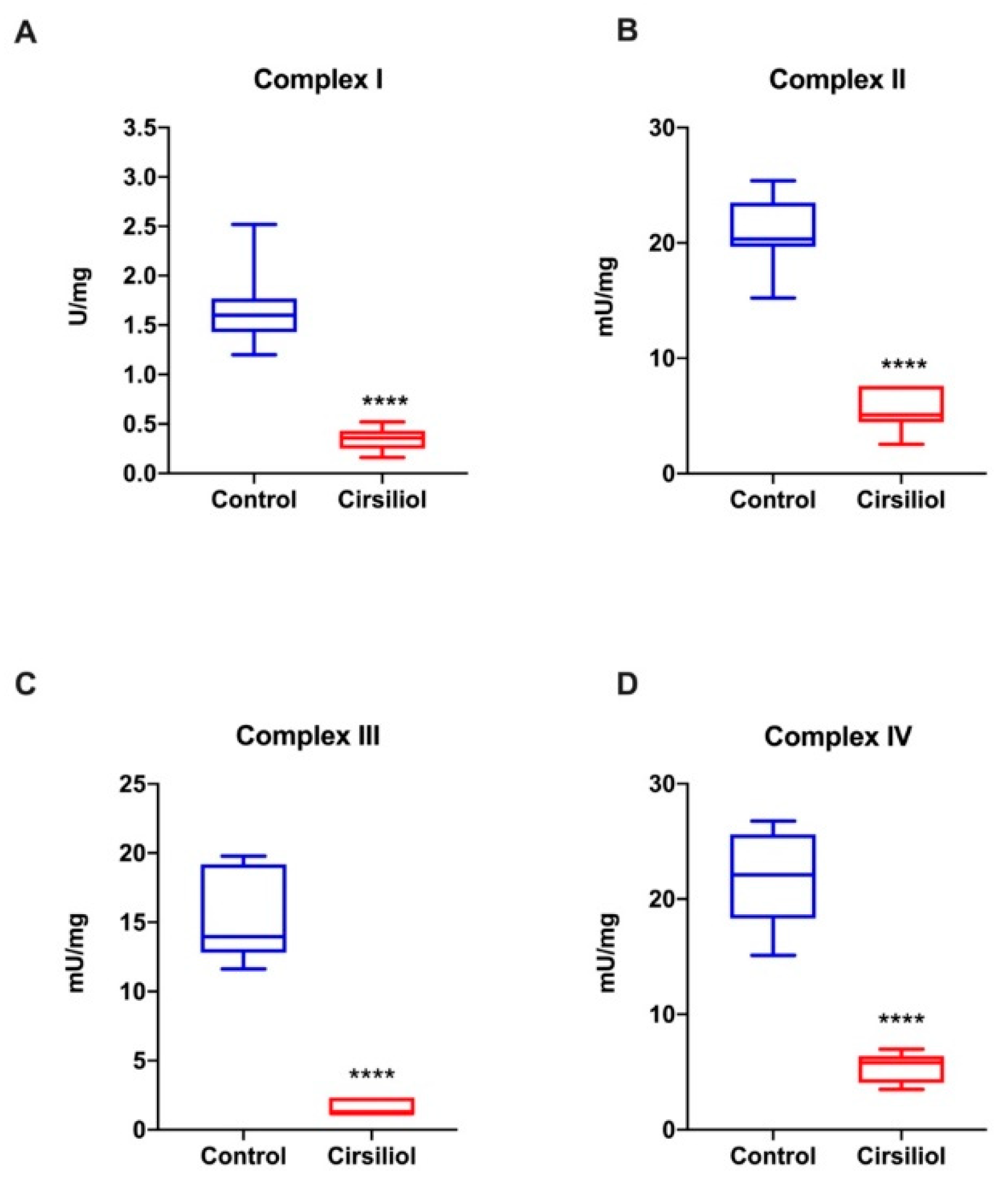
3.5. Cirsiliol Inhibits the Activity of the OS Respiratory Complexes Exclusively When in Coupled Conditions
3.6. Cirsiliol Decreased Reactive Oxygen Intermediates Production in the Light-Exposed Rod OS, Lowering the Oxidative and Structural Damage
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Pandey, K.B.; Rizvi, S.I. Plant Polyphenols as Dietary Antioxidants in Human Health and Disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scalbert, A.; Manach, C.; Morand, C.; Rémésy, C.; Jiménez, L. Dietary Polyphenols and the Prevention of Diseases. Crit. Rev. Food Sci. Nutr. 2005, 45, 287–306. [Google Scholar] [CrossRef] [PubMed]
- Bhullar, K.S.; Rupasinghe, H.P.V. Polyphenols: Multipotent Therapeutic Agents in Neurodegenerative Diseases. Oxid. Med. Cell. Longev. 2013, 2013, 891748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.B.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxid. Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattera, R.; Benvenuto, M.; Giganti, M.G.; Tresoldi, I.; Pluchinotta, F.R.; Bergante, S.; Tettamanti, G.; Masuelli, L.; Manzari, V.; Modesti, A.; et al. Effects of Polyphenols on Oxidative Stress-Mediated Injury in Cardiomyocytes. Nutrients 2017, 9, 523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bungau, S.; Abdel-Daim, M.M.; Tit, D.M.; Ghanem, E.; Sato, S.; Maruyama-Inoue, M.; Yamane, S.; Kadonosono, K. Health Benefits of Polyphenols and Carotenoids in Age-Related Eye Diseases. Oxidative Med. Cell. Longev. 2019, 2019, 9783429. [Google Scholar] [CrossRef] [PubMed]
- Tungmunnithum, D.; Thongboonyou, A.; Pholboon, A.; Yangsabai, A. Flavonoids and Other Phenolic Compounds from Medicinal Plants for Pharmaceutical and Medical Aspects: An Overview. Medicines 2018, 5, 93. [Google Scholar] [CrossRef]
- Gledhill, J.R.; Montgomery, M.G.; Leslie, A.G.; Walker, J.E. Mechanism of Inhibition of Bovine F1-ATPase by Resveratrol and Related Polyphenols. Proc. Natl. Acad. Sci. USA 2007, 104, 13632–13637. [Google Scholar] [CrossRef] [Green Version]
- Panfoli, I.; Calzia, D.; Ravera, S.; Bruschi, M.; Tacchetti, C.; Candiani, S.; Morelli, A.; Candiano, G. Extramitochondrial Tricarboxylic Acid Cycle in Retinal Rod Outer Segments. Biochimie 2011, 93, 1565–1575. [Google Scholar] [CrossRef]
- Panfoli, I.; Calzia, D.; Bruschi, M.; Oneto, M.; Bianchini, P.; Ravera, S.; Petretto, A.; Diaspro, A.; Candiano, G. Functional Expression of Oxidative Phosphorylation Proteins in the Rod Outer Segment Disc. Cell Biochem. Funct. 2013, 31, 532–538. [Google Scholar] [CrossRef]
- Bruschi, M.; Bartolucci, M.; Petretto, A.; Calzia, D.; Caicci, F.; Manni, L.; Traverso, C.E.; Candiano, G.; Panfoli, I. Differential Expression of the Five Redox Complexes in the Retinal Mitochondria or Rod Outer Segment Disks Is Consistent with Their Different Functionality. FASEB Bioadv. 2020, 2, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Bruschi, M.; Petretto, A.; Caicci, F.; Bartolucci, M.; Calzia, D.; Santucci, L.; Manni, L.; Ramenghi, L.A.; Ghiggeri, G.; Traverso, C.E.; et al. Proteome of Bovine Mitochondria and Rod Outer Segment Disks: Commonalities and Differences. J. Proteome Res. 2018, 17, 918–925. [Google Scholar] [CrossRef] [PubMed]
- Panfoli, I.; Musante, L.; Bachi, A.; Ravera, S.; Calzia, D.; Cattaneo, A.; Bruschi, M.; Bianchini, P.; Diaspro, A.; Morelli, A.; et al. Proteomic Analysis of the Retinal Rod Outer Segment Disks. J. Proteome Res. 2008, 7, 2654–2669. [Google Scholar] [CrossRef]
- Ridge, K.D.; Abdulaev, N.G.; Sousa, M.; Palczewski, K. Phototransduction: Crystal Clear. Trends Biochem. Sci. 2003, 28, 479–487. [Google Scholar] [CrossRef]
- Palczewski, K. Chemistry and Biology of the Initial Steps in Vision: The Friedenwald Lecture. Investig. Ophthalmol. Vis. Sci. 2014, 55, 6651–6672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calzia, D.; Oneto, M.; Caicci, F.; Bianchini, P.; Ravera, S.; Bartolucci, M.; Diaspro, A.; Degan, P.; Manni, L.; Traverso, C.E.; et al. Effect of Polyphenolic Phytochemicals on Ectopic Oxidative Phosphorylation in Rod Outer Segments of Bovine Retina. Br. J. Pharmacol. 2015, 172, 3890–3903. [Google Scholar] [CrossRef] [Green Version]
- Calzia, D.; Degan, P.; Caicci, F.; Bruschi, M.; Manni, L.; Ramenghi, L.A.; Candiano, G.; Traverso, C.E.; Panfoli, I. Modulation of the Rod Outer Segment Aerobic Metabolism Diminishes the Production of Radicals Due to Light Absorption. Free Radic. Biol. Med. 2018, 117, 110–118. [Google Scholar] [CrossRef] [Green Version]
- Bisio, A.; Schito, A.M.; Pedrelli, F.; Danton, O.; Reinhardt, J.K.; Poli, G.; Tuccinardi, T.; Bürgi, T.; de Riccardis, F.; Giacomini, M.; et al. Antibacterial and ATP Synthesis Modulating Compounds from Salvia Tingitana. J. Nat. Prod. 2020, 83, 1027–1042. [Google Scholar] [CrossRef]
- Ravera, S.; Esposito, A.; Degan, P.; Caicci, F.; Manni, L.; Liguori, A.; Bisio, A.; Iobbi, V.; Schito, A.; Traverso, C.E.; et al. The Diterpene Manool Extracted from Salvia Tingitana Lowers Free Radical Production in Retinal Rod Outer Segments by Inhibiting the Extramitochondrial F1Fo ATP Synthase. Cell Biochem. Funct. 2021, 39, 528–535. [Google Scholar] [CrossRef]
- Ravera, S.; Esposito, A.; Degan, P.; Caicci, F.; Calzia, D.; Perrotta, E.; Manni, L.; Bisio, A.; Iobbi, V.; Schito, A.; et al. Sclareol Modulates Free Radical Production in the Retinal Rod Outer Segment by Inhibiting the Ectopic F1fo-Atp Synthase. Free Radic. Biol. Med. 2020, 160, 368–375. [Google Scholar] [CrossRef]
- Hu, G.X.; Takano, A.; Drew, B.T.; de Liu, E.; Soltis, D.E.; Soltis, P.S.; Peng, H.; Xiang, C.L. Phylogeny and Staminal Evolution of Salvia (Lamiaceae, Nepetoideae) in East Asia. Ann. Bot. 2018, 122, 649. [Google Scholar] [CrossRef] [PubMed]
- Drew, B.T.; González-Gallegos, J.G.; Xiang, C.L.; Kriebel, R.; Drummond, C.P.; Walker, J.B.; Sytsma, K.J. Salvia United: The Greatest Good for the Greatest Number. Taxon 2017, 66, 133–145. [Google Scholar] [CrossRef] [Green Version]
- Will, M.; Claßen-Bockhoff, R. Time to Split Salvia s.l. (Lamiaceae)—New Insights from Old World Salvia Phylogeny. Mol. Phylogenet. Evol. 2017, 109, 33–58. [Google Scholar] [CrossRef] [PubMed]
- Will, M.; Schmalz, N.; Classen-Bockhoff, R. Towards a New Classification of Salvia s.l.: (Re)Establishing the Genus Pleudia Raf. Turk. J. Bot. 2015, 39, 693–707. [Google Scholar] [CrossRef]
- Lu, Y.; Yeap Foo, L. Polyphenolics of Salvia—A Review. Phytochemistry 2002, 59, 117–140. [Google Scholar] [CrossRef]
- Mustafa, E.H.; Abu Zarga, M.; Abdalla, S. Effects of Cirsiliol, a Flavone Isolated from Achillea Fragrantissima, on Rat Isolated Ileum. Gen. Pharmacol. 1992, 23, 555–560. [Google Scholar] [CrossRef]
- Bisio, A.; Fraternale, D.; Damonte, G.; Millo, E.; Lanteri, A.P.; Russo, E.; Romussi, G.; Parodi, B.; Ricci, D.; de Tommasi, N. Phytotoxic Activity of Salvia x Jamensis. Nat. Prod. Commun. 2009, 4, 1621–1630. [Google Scholar] [CrossRef] [Green Version]
- Lim, H.J.; Jang, H.J.; Bak, S.G.; Lee, S.; Lee, S.W.; Lee, K.M.; Lee, S.J.; Rho, M.C. In Vitro Inhibitory Effects of Cirsiliol on IL-6-Induced STAT3 Activation through Anti-Inflammatory Activity. Bioorg. Med. Chem. Lett. 2019, 29, 1586–1592. [Google Scholar] [CrossRef]
- Calixto, J.B.; Otuki, M.F.; Santos, A.R.S. Anti-Inflammatory Compounds of Plant Origin. Part I. Action on Arachidonic Acid Pathway, Nitric Oxide and Nuclear Factor Kappa B (NF-KappaB). Planta Med. 2003, 69, 973–983. [Google Scholar] [CrossRef]
- Forgo, P.; Zupkó, I.; Molnár, J.; Vasas, A.; Dombi, G.; Hohmann, J. Bioactivity-Guided Isolation of Antiproliferative Compounds from Centaurea Jacea L. Fitoterapia 2012, 83, 921–925. [Google Scholar] [CrossRef]
- Prasad, P.; Vasas, A.; Hohmann, J.; Bishayee, A.; Sinha, D. Cirsiliol Suppressed Epithelial to Mesenchymal Transition in B16F10 Malignant Melanoma Cells through Alteration of the PI3K/Akt/NF-ΚB Signaling Pathway. Int. J. Mol. Sci. 2019, 20, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, X.; Huang, C.; Hu, Y.; Wu, Q.; Liu, F.; Nie, W.; Chen, H.; Li, X.; Dong, Z.; Liu, K. Cirsiliol Targets Tyrosine Kinase 2 to Inhibit Esophageal Squamous Cell Carcinoma Growth in Vitro and in Vivo. J. Exp. Clin. Cancer Res. 2021, 40, 105. [Google Scholar] [CrossRef] [PubMed]
- Marder, M.; Viola, H.; Wasowski, C.; Wolfman, C.; Waterman, P.G.; Medina, J.H.; Paladini, A.C. Cirsiliol and Caffeic Acid Ethyl Ester, Isolated from Salvia Guaranitica, Are Competitive Ligands for the Central Benzodiazepine Receptors. Phytomedicine 1996, 3, 29–31. [Google Scholar] [CrossRef]
- Iinuma, M.; Matsuura, S.; Kusuda, K. ^<13>C-Nuclear Magnetic Resonance (NMR) Spectral Studies on Polysubstituted Flavonoids. I. 13C-NMR Spectra of Flavones. Chem. Pharm. Bull. 1980, 28, 708–716. [Google Scholar] [CrossRef] [Green Version]
- Panfoli, I.; Calzia, D.; Ravera, S.; Bianchini, P.; Diaspro, A. Maximizing the Rod Outer Segment Yield in Retinas Extracted from Cattle Eyes. Bio. Protoc. 2022, 12, e4474. [Google Scholar] [CrossRef]
- Calzia, D.; Barabino, S.; Bianchini, P.; Garbarino, G.; Oneto, M.; Caicci, F.; Diaspro, A.; Tacchetti, C.; Manni, L.; Candiani, S.; et al. New Findings in ATP Supply in Rod Outer Segments: Insights for Retinopathies. Biol. Cell 2013, 105, 345–358. [Google Scholar] [CrossRef]
- Schnetkamp, P.P.; Daemen, F.J. Isolation and Characterization of Osmotically Sealed Bovine Rod Outer Segments. Methods. Enzymol. 1982, 81, 110–116. [Google Scholar]
- Panfoli, I.; Calzia, D.; Bianchini, P.; Ravera, S.; Diaspro, A.; Candiano, G.; Bachi, A.; Monticone, M.; Aluigi, M.G.; Barabino, S.; et al. Evidence for Aerobic Metabolism in Retinal Rod Outer Segment Disks. Int. J. Biochem. Cell Biol. 2009, 41, 2555–2565. [Google Scholar] [CrossRef]
- Calzia, D.; Candiani, S.; Garbarino, G.; Caicci, F.; Ravera, S.; Bruschi, M.; Manni, L.; Morelli, A.; Traverso, C.E.; Candiano, G.; et al. Are Rod Outer Segment ATP-Ase and ATP-Synthase Activity Expression of the Same Protein? Cell Mol Neurobiol 2013, 33, 637–649. [Google Scholar] [CrossRef]
- Pinzi, L.; Rastelli, G. Molecular Docking: Shifting Paradigms in Drug Discovery. Int. J. Mol. Sci. 2019, 20, 4331. [Google Scholar] [CrossRef] [Green Version]
- McGann, M. FRED Pose Prediction and Virtual Screening Accuracy. J. Chem. Inf. Model. 2011, 51, 578–596. [Google Scholar] [CrossRef] [PubMed]
- Mangiullo, R.; Gnoni, A.; Leone, A.; Gnoni, G.V.; Papa, S.; Zanotti, F. Structural and Functional Characterization of FoF1-ATP Synthase on the Extracellular Surface of Rat Hepatocytes. Biochim. Biophys. Acta. Bioenerg. 2008, 1777, 1326–1335. [Google Scholar] [CrossRef] [Green Version]
- Arakaki, N.; Nagao, T.; Niki, R.; Toyofuku, A.; Tanaka, H.; Kuramoto, Y.; Emoto, Y.; Shibata, H.; Magota, K.; Higuti, T. Possible Role of Cell Surface H+ -ATP Synthase in the Extracellular ATP Synthesis and Proliferation of Human Umbilical Vein Endothelial Cells. Mol. Cancer Res. 2003, 1, 931–939. [Google Scholar] [PubMed]
- Kita, T.; Arakaki, N. Contribution of Extracellular ATP on the Cell-Surface F1F0-ATP Synthase-Mediated Intracellular Triacylglycerol Accumulation. Biomed. Res. 2015, 36, 115–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravera, S.; Bartolucci, M.; Garbati, P.; Ferrando, S.; Calzia, D.; Ramoino, P.; Balestrino, M.; Morelli, A.; Panfoli, I. Evaluation of the Acquisition of the Aerobic Metabolic Capacity by Myelin, during Its Development. Mol. Neurobiol. 2015, 53, 7048–7056. [Google Scholar] [CrossRef] [PubMed]
- Panfoli, I.; Ravera, S.; Podestà, M.; Cossu, C.; Santucci, L.; Bartolucci, M.; Bruschi, M.; Calzia, D.; Sabatini, F.; Bruschettini, M.; et al. Exosomes from Human Mesenchymal Stem Cells Conduct Aerobic Metabolism in Term and Preterm Newborn Infants. FASEB J. 2015, 30, 1416–1424. [Google Scholar] [CrossRef] [Green Version]
- Bruschi, M.; Santucci, L.; Ravera, S.; Bartolucci, M.; Petretto, A.; Calzia, D.; Ghiggeri, G.M.; Ramenghi, L.A.; Candiano, G.; Panfoli, I. Metabolic Signature of Microvesicles from Umbilical Cord Mesenchymal Stem Cells of Preterm and Term Infants. Proteom. Clin. Appl. 2018, 12, e1700082. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.-H.; Song, S.-H.; Paik, J.-Y.; Koh, B.-H.; Choe, Y.S.; Lee, E.J.; Kim, B.-T.; Lee, K.-H. Direct Targeting of Tumor Cell F(1)F(0) ATP-Synthase by Radioiodine Angiostatin in Vitro and in Vivo. Cancer Biother. Radiopharm. 2007, 22, 704–712. [Google Scholar] [CrossRef]
- Lu, Z.J.; Song, Q.F.; Jiang, S.S.; Song, Q.; Wang, W.; Zhang, G.H.; Kan, B.; Chen, L.J.; Yang, J.L.; Luo, F.; et al. Identification of ATP Synthase Beta Subunit (ATPB) on the Cell Surface as a Non-Small Cell Lung Cancer (NSCLC) Associated Antigen. BMC Cancer 2009, 9, 16. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Li, Y.; Li, G.; Ng Zhou, Z.; Chang, X.; Xia, Y.; Dong, X.; Liu, Z.; Ren, B.; Liu, W.; et al. Ectopic Expression of the ATP Synthase β Subunit on the Membrane of PC-3M Cells Supports Its Potential Role in Prostate Cancer Metastasis. Int. J. Oncol. 2017, 50, 1312–1320. [Google Scholar] [CrossRef] [Green Version]
- Panfoli, I.; Ravera, S.; Bruschi, M.; Candiano, G.; Morelli, A. Proteomics Unravels the Exportability of Mitochondrial Respiratory Chains. Expert Rev. Proteom. 2011, 8, 231–239. [Google Scholar] [CrossRef]
- Wood dos Santos, T.; Cristina Pereira, Q.; Teixeira, L.; Gambero, A.; A. Villena, J.; Lima Ribeiro, M. Effects of Polyphenols on Thermogenesis and Mitochondrial Biogenesis. Int. J. Mol. Sci. 2018, 19, 2757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panfoli, I. Extramitochondrial aerobic metabolism in retinal rod outer segment disks. Med. Hopytheses 2012, 78, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Genova, M.L.; Baracca, A.; Biondi, A.; Casalena, G.; Faccioli, M.; Falasca, A.I.; Formiggini, G.; Sgarbi, G.; Solaini, G.; Lenaz, G. Is Supercomplex Organization of the Respiratory Chain Required for Optimal Electron Transfer Activity? Biochim. Biophys. Acta 2008, 1777, 740–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roehlecke, C.; Schumann, U.; Ader, M.; Brunssen, C.; Bramke, S.; Morawietz, H.; Funk, R.H. Stress Reaction in Outer Segments of Photoreceptors after Blue Light Irradiation. PLoS ONE 2013, 8, e71570. [Google Scholar] [CrossRef] [PubMed]
- Calzia, D.; Panfoli, I.; Heinig, N.; Schumann, U.; Ader, M.; Traverso, C.E.; Funk, R.H.W.; Roehlecke, C. Impairment of Extramitochondrial Oxidative Phosphorylation in Mouse Rod Outer Segments by Blue Light Irradiation. Biochimie 2016, 125, 171–178. [Google Scholar] [CrossRef]
- Shaw, P.X.; Stiles, T.; Douglas, C.; Ho, D.; Fan, W.; Du, H.; Xiao, X. Oxidative Stress, Innate Immunity, and Age-Related Macular Degeneration. AIMS Mol. Sci. 2016, 3, 196–221. [Google Scholar] [CrossRef]
- Tan, B.L.; Norhaizan, M.E.; Liew, W.P.P.; Rahman, H.S. Antioxidant and Oxidative Stress: A Mutual Interplay in Age-Related Diseases. Front. Pharmacol. 2018, 9, 1162. [Google Scholar] [CrossRef] [Green Version]
- Bellezza, I. Oxidative Stress in Age-Related Macular Degeneration: NRF2 as Therapeutic Target. Front. Pharmacol. 2018, 9, 1280. [Google Scholar] [CrossRef]
- Domènech, E.B.; Marfany, G. The Relevance of Oxidative Stress in the Pathogenesis and Therapy of Retinal Dystrophies. Antioxidants 2020, 9. [Google Scholar] [CrossRef] [Green Version]
- Roehlecke, C.; Schumann, U.; Ader, M.; Knels, L.; Funk, R.H. Influence of Blue Light on Photoreceptors in a Live Retinal Explant System. Mol. Vis. 2011, 17, 876–884. [Google Scholar] [PubMed]
- Braun, R.D.; Linsenmeier, R.A.; Goldstick, T.K. Oxygen Consumption in the Inner and Outer Retina of the Cat. Investig. Ophthalmol. Vis. Sci. 1995, 36, 542–554. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carlini, L.; Tancreda, G.; Iobbi, V.; Caicci, F.; Bruno, S.; Esposito, A.; Calzia, D.; Benini, S.; Bisio, A.; Manni, L.; et al. The Flavone Cirsiliol from Salvia x jamensis Binds the F1 Moiety of ATP Synthase, Modulating Free Radical Production. Cells 2022, 11, 3169. https://doi.org/10.3390/cells11193169
Carlini L, Tancreda G, Iobbi V, Caicci F, Bruno S, Esposito A, Calzia D, Benini S, Bisio A, Manni L, et al. The Flavone Cirsiliol from Salvia x jamensis Binds the F1 Moiety of ATP Synthase, Modulating Free Radical Production. Cells. 2022; 11(19):3169. https://doi.org/10.3390/cells11193169
Chicago/Turabian StyleCarlini, Lavinia, Gabriele Tancreda, Valeria Iobbi, Federico Caicci, Silvia Bruno, Alfonso Esposito, Daniela Calzia, Stefano Benini, Angela Bisio, Lucia Manni, and et al. 2022. "The Flavone Cirsiliol from Salvia x jamensis Binds the F1 Moiety of ATP Synthase, Modulating Free Radical Production" Cells 11, no. 19: 3169. https://doi.org/10.3390/cells11193169
APA StyleCarlini, L., Tancreda, G., Iobbi, V., Caicci, F., Bruno, S., Esposito, A., Calzia, D., Benini, S., Bisio, A., Manni, L., Schito, A., Traverso, C. E., Ravera, S., & Panfoli, I. (2022). The Flavone Cirsiliol from Salvia x jamensis Binds the F1 Moiety of ATP Synthase, Modulating Free Radical Production. Cells, 11(19), 3169. https://doi.org/10.3390/cells11193169