
The Role of S. cerevisiae Sub1/PC4 in Transcription Elongation Depends on the C-Terminal Region and Is Independent of the ssDNA Binding Domain
 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains, Media and Generation of Strains
2.2. Co-Immunoprecipitation and Western Blot Analysis
2.3. Chromatin Immunoprecipitation (ChIP)
2.4. Transcriptional Run-On (TRO)
2.5. RNA Isolation and RT-PCR
2.6. GLAM Assay
2.7. 6 -Azauracil Induction (6AU)
2.8. Statistical Analysis
2.9. Sequence Alignment, Prediction and Modelling of Sc and Sp Sub1 Structures
3. Results
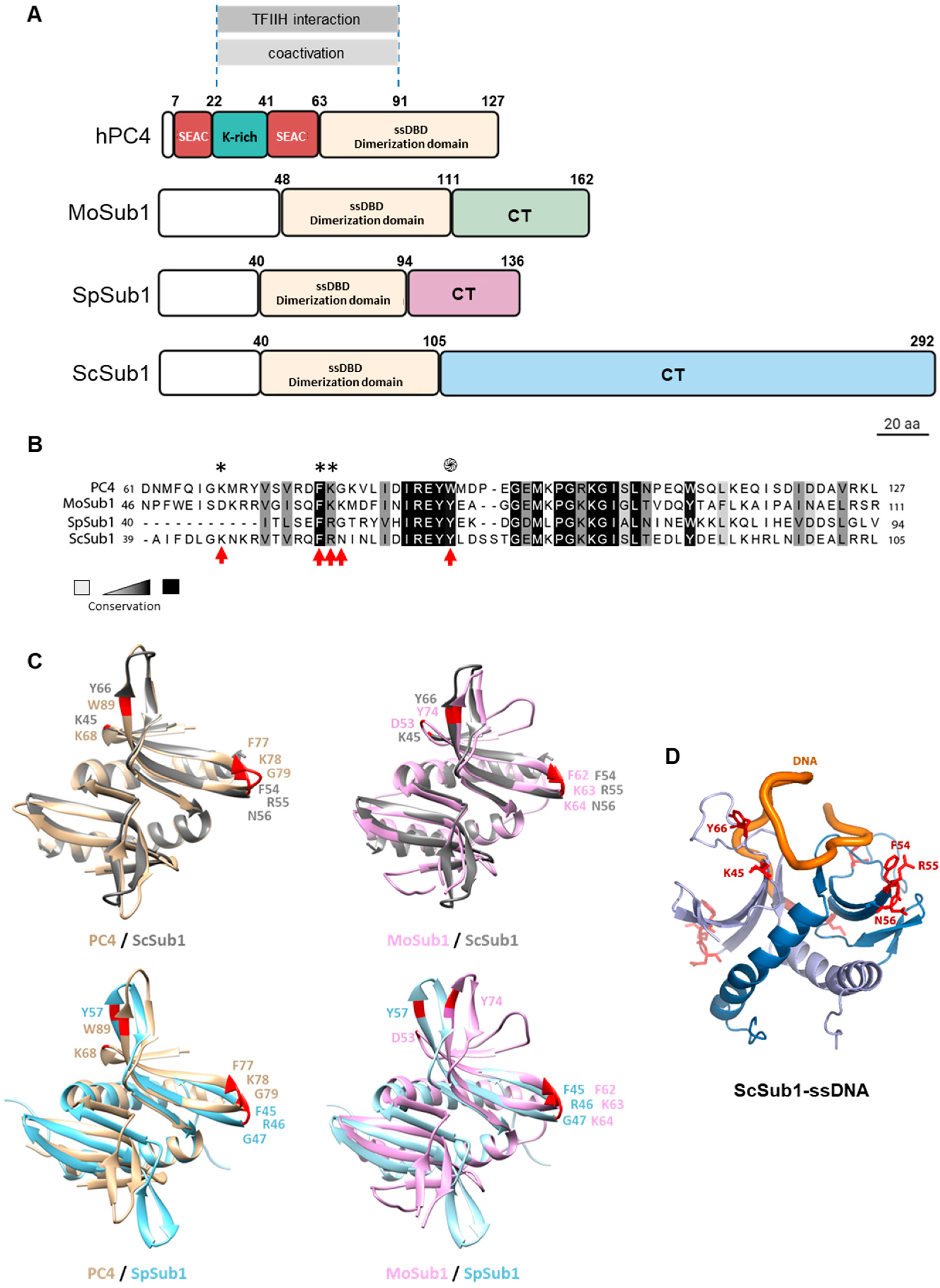
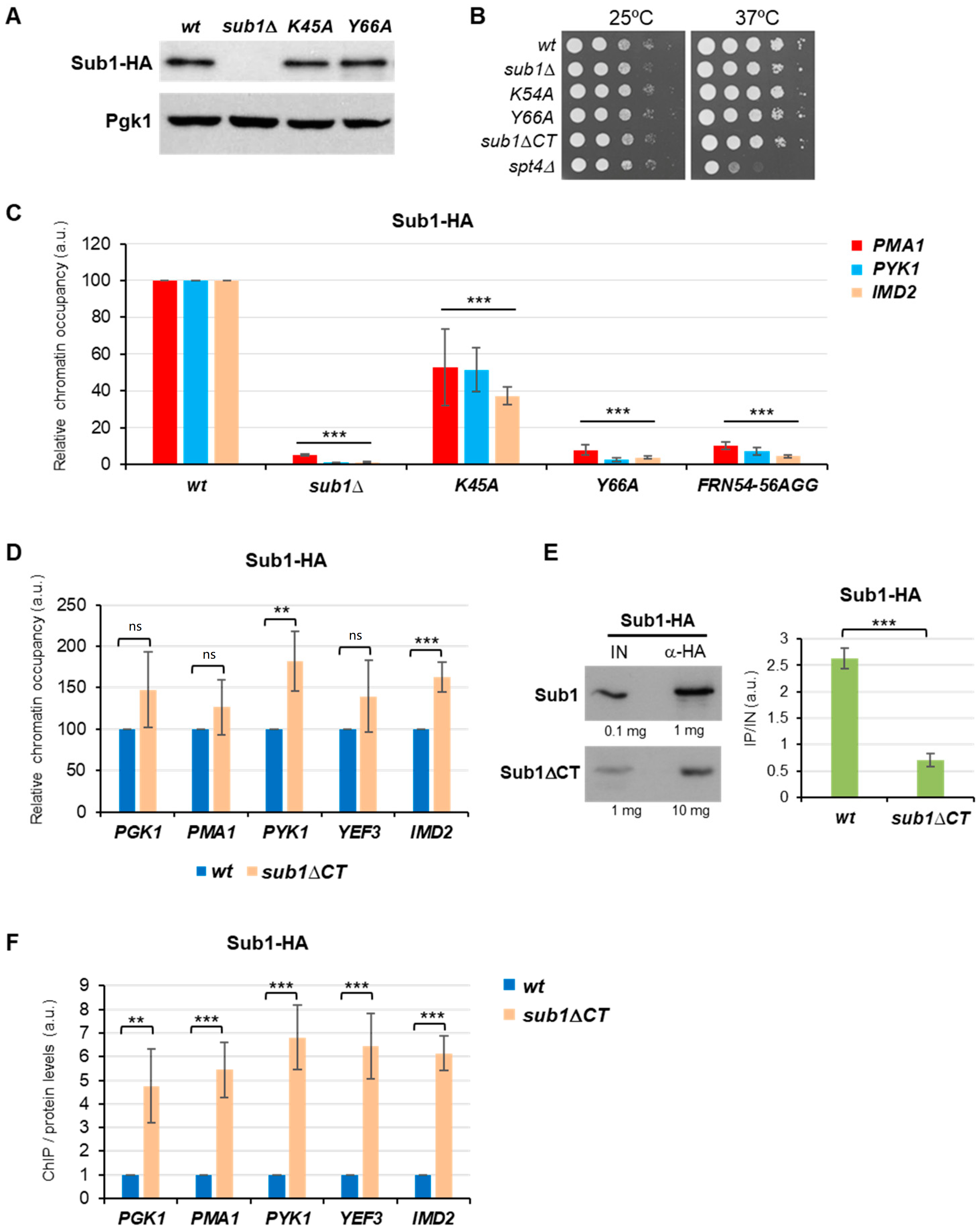
3.1. Sub1 ssDNA Binding Domain Shares Homology and Binding Capacity with Human PC4
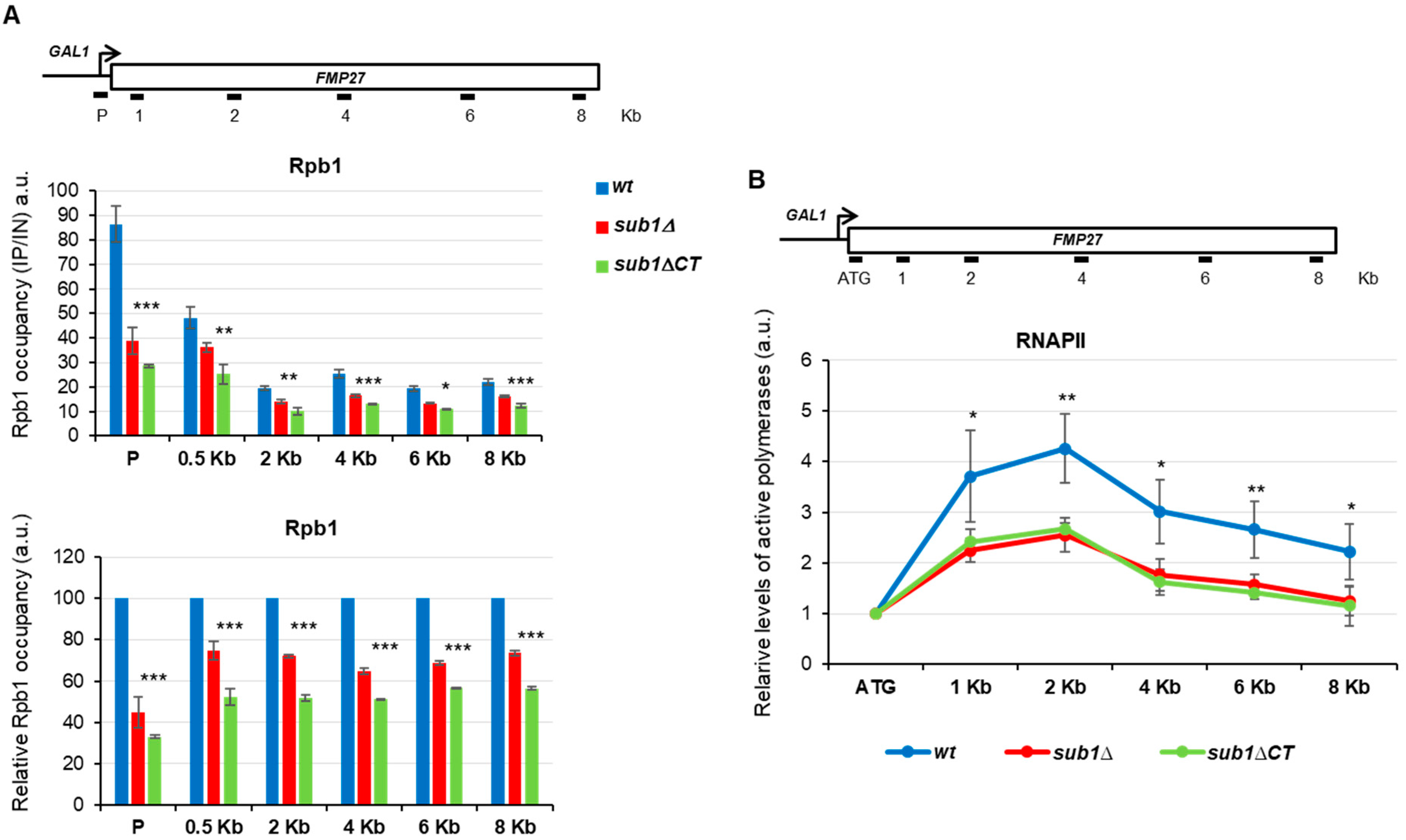
3.2. The C-Terminal Region of Sub1 Is Required for Proper Transcription Elongation
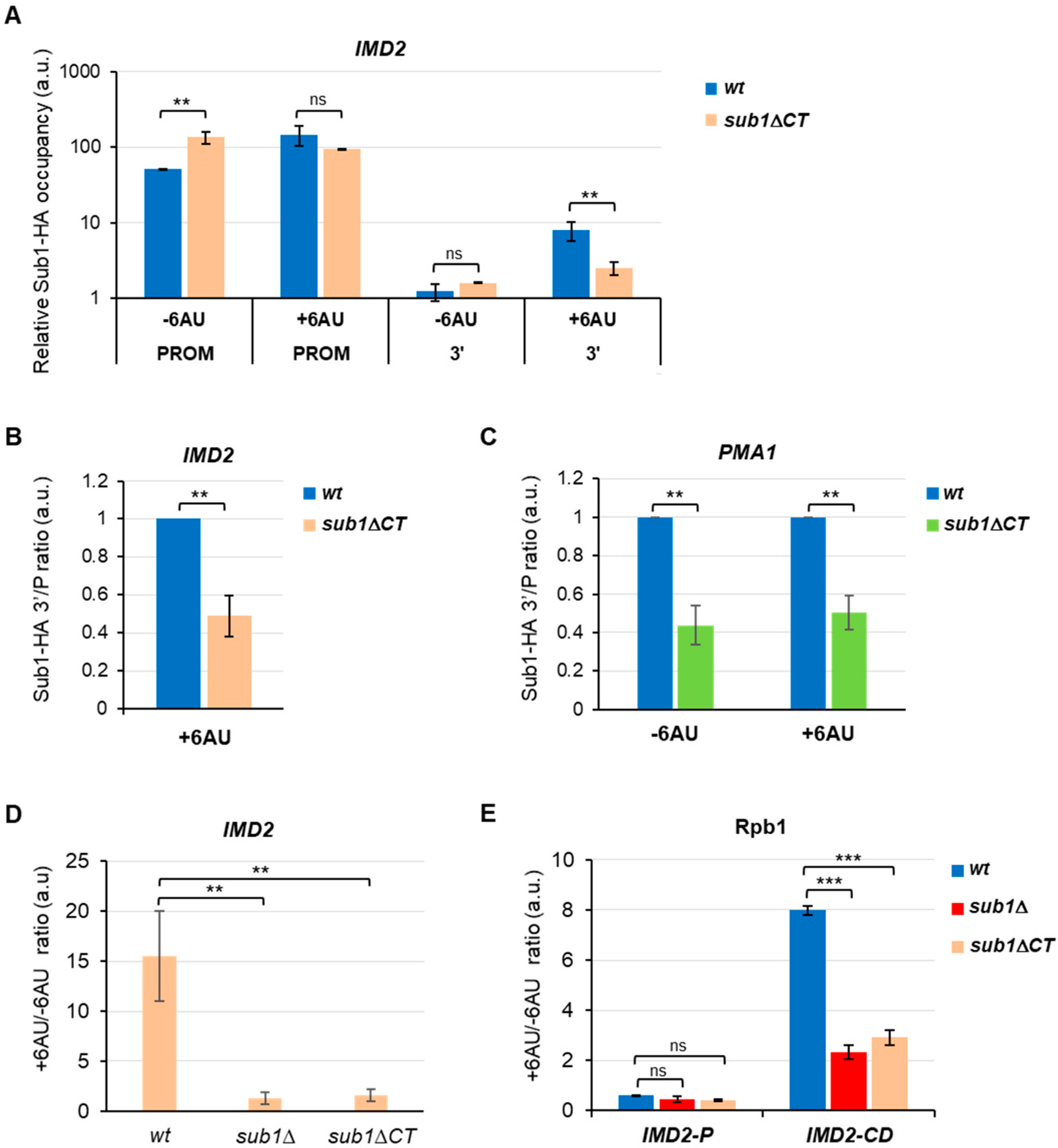
3.3. Sub1 Participates in the Expression of the IMD2 Gene trough the CT Region
4. Discussion
4.1. PC4/Sub1 ssDNA Binding Domain Is Functionally Conserved
4.2. S. cerevisiae Sub1 Contains an Unusual CT Extension with a Role in Transcription Elongation
4.3. Understanding the Mechanisms Underlying ScSub1 Functions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Petrenko, N.; Struhl, K. Comparison of transcriptional initiation by RNA polymerase II across eukaryotic species. eLife 2021, 10, e67964. [Google Scholar] [CrossRef]
- Malik, S.; Guermah, M.; Roeder, R.G. A dynamic model for PC4 coactivator function in RNA polymerase II transcription. Proc. Natl. Acad. Sci. USA 1998, 95, 2192–2197. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, K.; Meisterernst, M. The human general co-factors. Trends Biochem. Sci. 1996, 21, 342–345. [Google Scholar] [CrossRef]
- Ge, H.; Roeder, R.G. Purification, cloning, and characterization of a human coactivator, PC4, that mediates transcriptional activation of class II genes. Cell 1994, 78, 513–523. [Google Scholar] [CrossRef]
- Henry, N.L.; Bushnell, D.A.; Kornberg, R.D. A yeast transcriptional stimulatory protein similar to human PC4. J. Biol. Chem. 1996, 271, 21842–21847. [Google Scholar] [CrossRef] [Green Version]
- Knaus, R.; Pollock, R.; Guarente, L. Yeast SUB1 is a suppressor of TFIIB mutations and has homology to the human co-activator PC4. EMBO J. 1996, 15, 1933–1940. [Google Scholar] [CrossRef]
- Huang, J.; Zhao, Y.; Liu, H.; Huang, D.; Cheng, X.; Zhao, W.; Taylor, I.A.; Liu, J.; Peng, Y.L. Substitution of tryptophan 89 with tyrosine switches the DNA binding mode of PC4. Sci. Rep. 2015, 5, 8789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contreras-Levicoy, J.; Urbina, F.; Maldonado, E. Schizosaccharomyces pombe positive cofactor 4 stimulates basal transcription from TATA-containing and TATA-less promoters through Mediator and transcription factor IIA. FEBS J. 2008, 275, 2873–2883. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhao, Y.; Huang, D.; Liu, H.; Justin, N.; Zhao, W.; Liu, J.; Peng, Y. Structural features of the single-stranded DNA-binding protein MoSub1 from Magnaporthe oryzae. Acta Crystallogr. D Biol. Crystallogr. 2012, 68, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Rossi, P.; Barbieri, C.M.; Aramini, J.M.; Bini, E.; Lee, H.W.; Janjua, H.; Xiao, R.; Acton, T.B.; Montelione, G.T. Structures of apo- and ssDNA-bound YdbC from Lactococcus lactis uncover the function of protein domain family DUF2128 and expand the single-stranded DNA-binding domain proteome. Nucleic Acids Res. 2013, 41, 2756–2768. [Google Scholar] [CrossRef]
- Werten, S.; Kohler, C.; Bayer, N.J.; Steinmetz, I.; Hinrichs, W. Structural analysis and knock-out of a Burkholderia pseudomallei homolog of the eukaryotic transcription coactivator PC4. Gene 2016, 577, 140–147. [Google Scholar] [CrossRef]
- Steigemann, B.; Schulz, A.; Werten, S. Bacteriophage T5 encodes a homolog of the eukaryotic transcription coactivator PC4 implicated in recombination-dependent DNA replication. J. Mol. Biol. 2013, 425, 4125–4133. [Google Scholar] [CrossRef]
- Brandsen, J.; Werten, S.; van der Vliet, P.C.; Meisterernst, M.; Kroon, J.; Gros, P. C-terminal domain of transcription cofactor PC4 reveals dimeric ssDNA binding site. Nat. Struct. Biol. 1997, 4, 900–903. [Google Scholar] [CrossRef] [PubMed]
- Werten, S.; Stelzer, G.; Goppelt, A.; Langen, F.M.; Gros, P.; Timmers, H.T.; Van der Vliet, P.C.; Meisterernst, M. Interaction of PC4 with melted DNA inhibits transcription. EMBO J. 1998, 17, 5103–5111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werten, S.; Wechselberger, R.; Boelens, R.; van der Vliet, P.C.; Kaptein, R. Identification of the single-stranded DNA binding surface of the transcriptional coactivator PC4 by NMR. J. Biol. Chem. 1999, 274, 3693–3699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Zhang, Y.; Huang, J.; Wang, S.; Yi, L.; Zhang, X.; Xu, M.; Fang, X.; Liu, J. The effect of phosphate ion on the ssDNA binding mode of MoSub1, a Sub1/PC4 homolog from rice blast fungus. Proteins 2019, 87, 257–264. [Google Scholar] [CrossRef]
- Werten, S.; Moras, D. A global transcription cofactor bound to juxtaposed strands of unwound DNA. Nat. Struct. Mol. Biol. 2006, 13, 181–182. [Google Scholar] [CrossRef]
- Wang, Z.; Roeder, R.G. DNA topoisomerase I and PC4 can interact with human TFIIIC to promote both accurate termination and transcription reinitiation by RNA polymerase III. Mol. Cell 1998, 1, 749–757. [Google Scholar] [CrossRef]
- Garavis, M.; Calvo, O. Sub1/PC4, a multifaceted factor: From transcription to genome stability. Curr. Genet. 2017, 63, 1023–1035. [Google Scholar] [CrossRef]
- Calvo, O. Sub1 and RNAPII, until termination does them part. Transcription 2018, 9, 52–60. [Google Scholar] [CrossRef]
- Rosonina, E.; Willis, I.M.; Manley, J.L. Sub1 functions in osmoregulation and in transcription by both RNA polymerases II and III. Mol. Cell. Biol. 2009, 29, 2308–2321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyama, H.; Sumiya, E.; Nagata, M.; Ito, T.; Sekimizu, K. Transcriptional repression of the IMD2 gene mediated by the transcriptional co-activator Sub1. Genes Cells 2008, 13, 1113–1126. [Google Scholar] [CrossRef] [PubMed]
- Braberg, H.; Jin, H.; Moehle, E.A.; Chan, Y.A.; Wang, S.; Shales, M.; Benschop, J.J.; Morris, J.H.; Qiu, C.; Hu, F.; et al. From structure to systems: High-resolution, quantitative genetic analysis of RNA polymerase II. Cell 2013, 154, 775–788. [Google Scholar] [CrossRef] [Green Version]
- Sikorski, T.W.; Ficarro, S.B.; Holik, J.; Kim, T.; Rando, O.J.; Marto, J.A.; Buratowski, S. Sub1 and RPA Associate with RNA Polymerase II at Different Stages of Transcription. Mol. Cell 2011, 44, 397–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, A.; Rosonina, E.; Manley, J.L.; Calvo, O. Sub1 globally regulates RNA polymerase II C-terminal domain phosphorylation. Mol. Cell. Biol. 2010, 30, 5180–5193. [Google Scholar] [CrossRef] [Green Version]
- Garcia, A.; Collin, A.; Calvo, O. Sub1 associates with Spt5 and influences RNA polymerase II transcription elongation rate. Mol. Biol. Cell 2012, 23, 4297–4312. [Google Scholar] [CrossRef] [Green Version]
- Calvo, O.; Manley, J.L. Evolutionarily conserved interaction between CstF-64 and PC4 links transcription, polyadenylation, and termination. Mol. Cell 2001, 7, 1013–1023. [Google Scholar] [CrossRef]
- Calvo, O.; Manley, J.L. Strange bedfellows: Polyadenylation factors at the promoter. Genes Dev. 2003, 17, 1321–1327. [Google Scholar] [CrossRef] [Green Version]
- Garavis, M.; Gonzalez-Polo, N.; Allepuz-Fuster, P.; Louro, J.A.; Fernandez-Tornero, C.; Calvo, O. Sub1 contacts the RNA polymerase II stalk to modulate mRNA synthesis. Nucleic Acids Res. 2017, 45, 2458–2471. [Google Scholar] [CrossRef] [Green Version]
- Burke, D.; Dawson, D.; Stearns, T. Methods in Yeast Genetics; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2000. [Google Scholar]
- Allepuz-Fuster, P.; Martinez-Fernandez, V.; Garrido-Godino, A.I.; Alonso-Aguado, S.; Hanes, S.D.; Navarro, F.; Calvo, O. Rpb4/7 facilitates RNA polymerase II CTD dephosphorylation. Nucleic Acids Res. 2014, 42, 13674–13688. [Google Scholar] [CrossRef]
- Birse, C.E.; Minvielle-Sebastia, L.; Lee, B.A.; Keller, W.; Proudfoot, N.J. Coupling termination of transcription to messenger RNA maturation in yeast. Science 1998, 280, 298–301. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, M.E.; Brown, T.A.; Trumpower, B.L. A rapid and simple method for preparation of RNA from Saccharomyces cerevisiae. Nucleic Acids Res. 1990, 18, 3091–3092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morillo-Huesca, M.; Vanti, M.; Chavez, S. A simple in vivo assay for measuring the efficiency of gene length-dependent processes in yeast mRNA biogenesis. FEBS J. 2006, 273, 756–769. [Google Scholar] [CrossRef] [PubMed]
- Pandey, B.; Dev, A.; Chakravorty, D.; Bhandare, V.V.; Polley, S.; Roy, S.; Basu, G. Insights on the disruption of the complex between human positive coactivator 4 and p53 by small molecules. Biochem. Biophys. Res. Commun. 2021, 578, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Mirdita, M.; Schutze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making protein folding accessible to all. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Meng, E.C.; Couch, G.S.; Croll, T.I.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Structure visualization for researchers, educators, and developers. Protein Sci. 2021, 30, 70–82. [Google Scholar] [CrossRef]
- Hartzog, G.A.; Basrai, M.A.; Ricupero-Hovasse, S.L.; Hieter, P.; Winston, F. Identification and analysis of a functional human homolog of the SPT4 gene of Saccharomyces cerevisiae. Mol. Cell. Biol. 1996, 16, 2848–2856. [Google Scholar] [CrossRef] [Green Version]
- Hartzog, G.A.; Wada, T.; Handa, H.; Winston, F. Evidence that Spt4, Spt5, and Spt6 control transcription elongation by RNA polymerase II in Saccharomyces cerevisiae. Genes Dev. 1998, 12, 357–369. [Google Scholar] [CrossRef] [Green Version]
- Hartzog, G.A.; Fu, J. The Spt4-Spt5 complex: A multi-faceted regulator of transcription elongation. Biochim. Biophys. Acta 2013, 1829, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Wada, T.; Takagi, T.; Yamaguchi, Y.; Ferdous, A.; Imai, T.; Hirose, S.; Sugimoto, S.; Yano, K.; Hartzog, G.A.; Winston, F.; et al. DSIF, a novel transcription elongation factor that regulates RNA polymerase II processivity, is composed of human Spt4 and Spt5 homologs. Genes Dev. 1998, 12, 343–356. [Google Scholar] [CrossRef]
- Swanson, M.S.; Malone, E.A.; Winston, F. SPT5, an essential gene important for normal transcription in Saccharomyces cerevisiae, encodes an acidic nuclear protein with a carboxy-terminal repeat. Mol. Cell. Biol. 1991, 11, 3009–3019. [Google Scholar] [CrossRef] [PubMed]
- Winston, F.; Chaleff, D.T.; Valent, B.; Fink, G.R. Mutations affecting Ty-mediated expression of the HIS4 gene of Saccharomyces cerevisiae. Genetics 1984, 107, 179–197. [Google Scholar] [CrossRef] [PubMed]
- Mason, P.B.; Struhl, K. Distinction and relationship between elongation rate and processivity of RNA polymerase II in vivo. Mol. Cell 2005, 17, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Hirayoshi, K.; Lis, J.T. Nuclear run-on assays: Assessing transcription by measuring density of engaged RNA polymerases. Methods Enzymol. 1999, 304, 351–362. [Google Scholar] [CrossRef]
- Exinger, F.; Lacroute, F. 6-Azauracil inhibition of GTP biosynthesis in Saccharomyces cerevisiae. Curr. Genet. 1992, 22, 9–11. [Google Scholar] [CrossRef]
- Kwapisz, M.; Wery, M.; Despres, D.; Ghavi-Helm, Y.; Soutourina, J.; Thuriaux, P.; Lacroute, F. Mutations of RNA polymerase II activate key genes of the nucleoside triphosphate biosynthetic pathways. EMBO J. 2008, 27, 2411–2421. [Google Scholar] [CrossRef] [Green Version]
- Shaw, R.J.; Reines, D. Saccharomyces cerevisiae transcription elongation mutants are defective in PUR5 induction in response to nucleotide depletion. Mol. Cell. Biol. 2000, 20, 7427–7437. [Google Scholar] [CrossRef] [Green Version]
- Riles, L.; Shaw, R.J.; Johnston, M.; Reines, D. Large-scale screening of yeast mutants for sensitivity to the IMP dehydrogenase inhibitor 6-azauracil. Yeast 2004, 21, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Malagon, F.; Kireeva, M.L.; Shafer, B.K.; Lubkowska, L.; Kashlev, M.; Strathern, J.N. Mutations in the Saccharomyces cerevisiae RPB1 gene conferring hypersensitivity to 6-azauracil. Genetics 2006, 172, 2201–2209. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Liu, Q.; Shi, T.; Yu, Y.; Lu, H. Genome-wide screen of fission yeast mutants for sensitivity to 6-azauracil, an inhibitor of transcriptional elongation. Yeast 2015, 32, 643–655. [Google Scholar] [CrossRef]
- Braberg, H.; Moehle, E.A.; Shales, M.; Guthrie, C.; Krogan, N.J. Genetic interaction analysis of point mutations enables interrogation of gene function at a residue-level resolution: Exploring the applications of high-resolution genetic interaction mapping of point mutations. Bioessays 2014, 36, 706–713. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Sarker, A.H.; Cooper, P.K.; Volkert, M.R. The single-strand DNA binding activity of human PC4 prevents mutagenesis and killing by oxidative DNA damage. Mol. Cell. Biol. 2004, 24, 6084–6093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Giles, C.; Li, S. Insights into how Spt5 functions in transcription elongation and repressing transcription coupled DNA repair. Nucleic Acids Res. 2014, 42, 7069–7083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Rucobo, F.W.; Sainsbury, S.; Cheung, A.C.; Cramer, P. Architecture of the RNA polymerase-Spt4/5 complex and basis of universal transcription processivity. EMBO J. 2011, 30, 1302–1310. [Google Scholar] [CrossRef]
- Jonker, H.R.; Wechselberger, R.W.; Pinkse, M.; Kaptein, R.; Folkers, G.E. Gradual phosphorylation regulates PC4 coactivator function. FEBS J. 2006, 273, 1430–1444. [Google Scholar] [CrossRef] [Green Version]
- Schang, L.M.; Hwang, G.J.; Dynlacht, B.D.; Speicher, D.W.; Bantly, A.; Schaffer, P.A.; Shilatifard, A.; Ge, H.; Shiekhattar, R. Human PC4 is a substrate-specific inhibitor of RNA polymerase II phosphorylation. J. Biol. Chem. 2000, 275, 6071–6074. [Google Scholar] [CrossRef] [Green Version]
- Rojas, D.A.; Urbina, F.; Solari, A.; Maldonado, E. The catalytic subunit of Schizosaccharomyces pombe CK2 (Cka1) negatively regualtes RNA polymerase II transcription through phosphorylation of positive cofactor (PC4). Int. J. Mol. Sci. 2022, 23, 9499. [Google Scholar] [CrossRef]
- Albuquerque, C.P.; Smolka, M.B.; Payne, S.H.; Bafna, V.; Eng, J.; Zhou, H. A multidimensional chromatography technology for in-depth phosphoproteome analysis. Mol. Cell. Proteom. MCP 2008, 7, 1389–1396. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.C.; Jiang, P.H.; Chen, H.M.; Chen, C.H.; Wang, Y.T.; Chen, Y.J.; Yu, C.J.; Teng, S.C. Glucose intake hampers PKA-regulated HSP90 chaperone activity. eLife 2018, 7, e39925. [Google Scholar] [CrossRef]
- Holt, L.J.; Tuch, B.B.; Villen, J.; Johnson, A.D.; Gygi, S.P.; Morgan, D.O. Global analysis of Cdk1 substrate phosphorylation sites provides insights into evolution. Science 2009, 325, 1682–1686. [Google Scholar] [CrossRef]
- Lanz, M.C.; Yugandhar, K.; Gupta, S.; Sanford, E.J.; Faca, V.M.; Vega, S.; Joiner, A.M.N.; Fromme, J.C.; Yu, H.; Smolka, M.B. In-depth and 3-dimensional exploration of the budding yeast phosphoproteome. EMBO Rep. 2021, 22, e51121. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Gerber, S.A.; Rudner, A.D.; Beausoleil, S.A.; Haas, W.; Villen, J.; Elias, J.E.; Gygi, S.P. Large-scale phosphorylation analysis of alpha-factor-arrested Saccharomyces cerevisiae. J. Proteome Res. 2007, 6, 1190–1197. [Google Scholar] [CrossRef] [PubMed]
- MacGilvray, M.E.; Shishkova, E.; Place, M.; Wagner, E.R.; Coon, J.J.; Gasch, A.P. Phosphoproteome Response to Dithiothreitol Reveals Unique Versus Shared Features of Saccharomyces cerevisiae Stress Responses. J. Proteome Res. 2020, 19, 3405–3417. [Google Scholar] [CrossRef] [PubMed]
- Pultz, D.; Bennetzen, M.V.; Rodkaer, S.V.; Zimmermann, C.; Enserink, J.M.; Andersen, J.S.; Faergeman, N.J. Global mapping of protein phosphorylation events identifies Ste20, Sch9 and the cell-cycle regulatory kinases Cdc28/Pho85 as mediators of fatty acid starvation responses in Saccharomyces cerevisiae. Mol. Biosyst. 2012, 8, 796–803. [Google Scholar] [CrossRef] [PubMed]
- Soulard, A.; Cremonesi, A.; Moes, S.; Schutz, F.; Jeno, P.; Hall, M.N. The rapamycin-sensitive phosphoproteome reveals that TOR controls protein kinase A toward some but not all substrates. Mol. Biol. Cell 2010, 21, 3475–3486. [Google Scholar] [CrossRef] [Green Version]
- Swaney, D.L.; Beltrao, P.; Starita, L.; Guo, A.; Rush, J.; Fields, S.; Krogan, N.J.; Villen, J. Global analysis of phosphorylation and ubiquitylation cross-talk in protein degradation. Nat. Methods 2013, 10, 676–682. [Google Scholar] [CrossRef]
- Calvo, O.; Manley, J.L. The transcriptional coactivator PC4/Sub1 has multiple functions in RNA polymerase II transcription. EMBO J. 2005, 24, 1009–1020. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Warfield, L.; Zhang, C.; Luo, J.; Allen, J.; Lang, W.H.; Ranish, J.; Shokat, K.M.; Hahn, S. Phosphorylation of the transcription elongation factor Spt5 by yeast Bur1 kinase stimulates recruitment of the PAF complex. Mol. Cell. Biol. 2009, 29, 4852–4863. [Google Scholar] [CrossRef] [Green Version]
- Zhou, K.; Kuo, W.H.; Fillingham, J.; Greenblatt, J.F. Control of transcriptional elongation and cotranscriptional histone modification by the yeast BUR kinase substrate Spt5. Proc. Natl. Acad. Sci. USA 2009, 106, 6956–6961. [Google Scholar] [CrossRef] [Green Version]
- Bataille, A.R.; Jeronimo, C.; Jacques, P.E.; Laramee, L.; Fortin, M.E.; Forest, A.; Bergeron, M.; Hanes, S.D.; Robert, F. A universal RNA polymerase II CTD cycle is orchestrated by complex interplays between kinase, phosphatase, and isomerase enzymes along genes. Mol. Cell 2012, 45, 158–170. [Google Scholar] [CrossRef]
- Lindstrom, D.L.; Squazzo, S.L.; Muster, N.; Burckin, T.A.; Wachter, K.C.; Emigh, C.A.; McCleery, J.A.; Yates, J.R., 3rd; Hartzog, G.A. Dual roles for Spt5 in pre-mRNA processing and transcription elongation revealed by identification of Spt5-associated proteins. Mol. Cell. Biol. 2003, 23, 1368–1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimeno-Gonzalez, S.; Haaning, L.L.; Malagon, F.; Jensen, T.H. The yeast 5′-3′ exonuclease Rat1p functions during transcription elongation by RNA polymerase II. Mol. Cell 2010, 37, 580–587. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Collin, A.; González-Jiménez, A.; González-Jiménez, M.d.C.; Alfonso, M.J.; Calvo, O. The Role of S. cerevisiae Sub1/PC4 in Transcription Elongation Depends on the C-Terminal Region and Is Independent of the ssDNA Binding Domain. Cells 2022, 11, 3320. https://doi.org/10.3390/cells11203320
Collin A, González-Jiménez A, González-Jiménez MdC, Alfonso MJ, Calvo O. The Role of S. cerevisiae Sub1/PC4 in Transcription Elongation Depends on the C-Terminal Region and Is Independent of the ssDNA Binding Domain. Cells. 2022; 11(20):3320. https://doi.org/10.3390/cells11203320
Chicago/Turabian StyleCollin, Alejandro, Araceli González-Jiménez, María del Carmen González-Jiménez, Manuel J. Alfonso, and Olga Calvo. 2022. "The Role of S. cerevisiae Sub1/PC4 in Transcription Elongation Depends on the C-Terminal Region and Is Independent of the ssDNA Binding Domain" Cells 11, no. 20: 3320. https://doi.org/10.3390/cells11203320
APA StyleCollin, A., González-Jiménez, A., González-Jiménez, M. d. C., Alfonso, M. J., & Calvo, O. (2022). The Role of S. cerevisiae Sub1/PC4 in Transcription Elongation Depends on the C-Terminal Region and Is Independent of the ssDNA Binding Domain. Cells, 11(20), 3320. https://doi.org/10.3390/cells11203320