Frontiers in Neurogenesis



{kind=link}
{kind=link}
Funding
Acknowledgments
Conflicts of Interest
References
- Altman, J. Are new neurons formed in the brains of adult mammals? Science 1962, 135, 1127–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altman, J.; Das, G.D. Autoradiographic and histological evidence of postnatal hippocampal neurogenesis in rats. J. Comp. Neurol. 1965, 124, 319–335. [Google Scholar] [CrossRef] [PubMed]
- Gross, C.G. Neurogenesis in the adult brain: Death of a dogma. Nat. Rev. Neurosci. 2000, 1, 67–73. [Google Scholar] [CrossRef]
- Kuhn, H.G.; Dickinson-Anson, H.; Gage, F.H. Neurogenesis in the dentate gyrus of the adult rat: Age-related decrease of neuronal progenitor proliferation. J. Neurosci. 1996, 16, 2027–2033. [Google Scholar] [CrossRef] [Green Version]
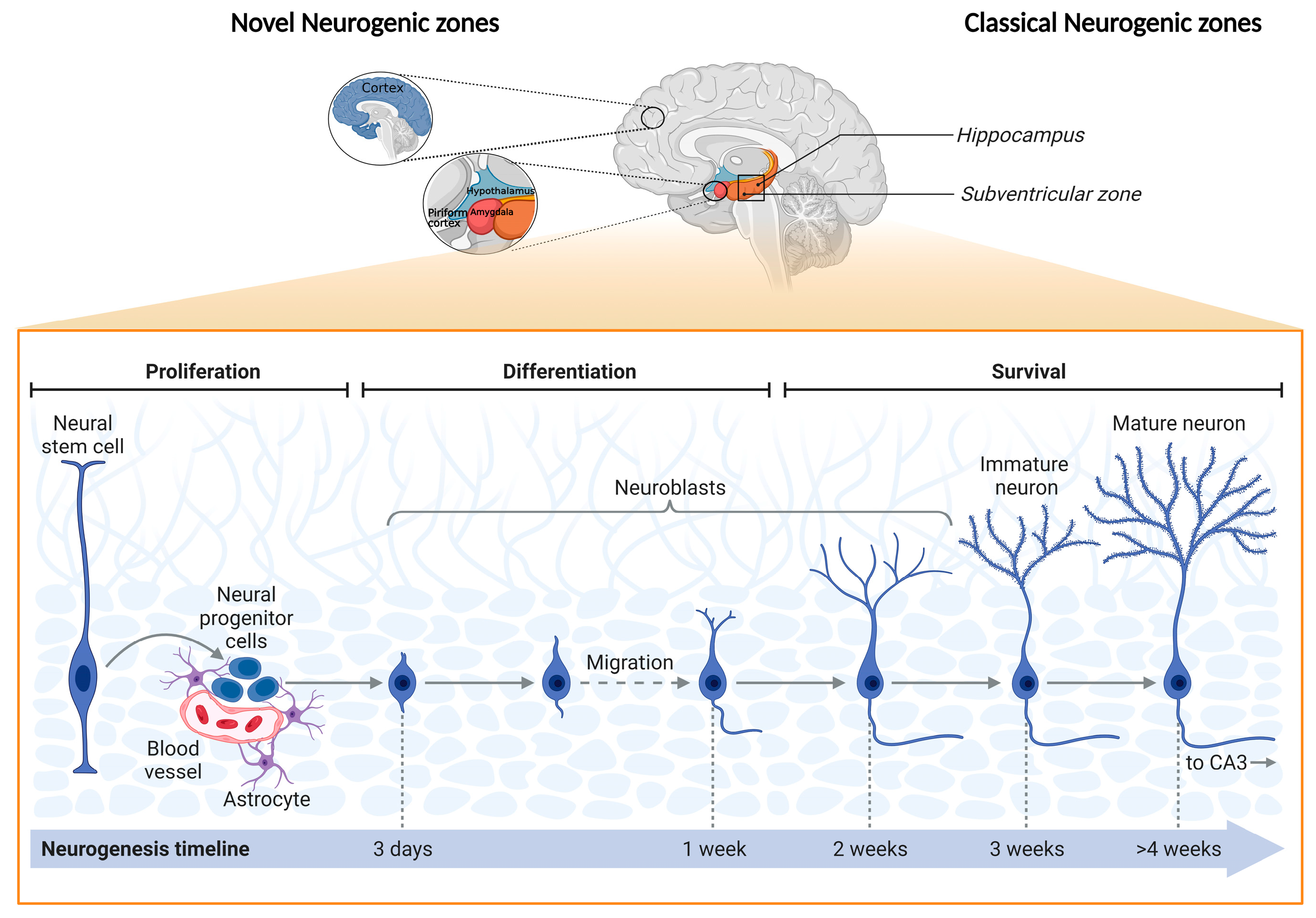
- Jurkowski, M.P.; Bettio, L.; Woo, E.K.; Patten, A.; Yau, S.-Y.; Gil-Mohapel, J. Beyond the Hippocampus and the SVZ: Adult Neurogenesis Throughout the Brain. Front. Cell. Neurosci. 2020, 14, 576444. [Google Scholar] [CrossRef] [PubMed]
- Gage, F.H. Adult neurogenesis in mammals. Science 2019, 364, 827–828. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, P.S.; Perfilieva, E.; Björk-Eriksson, T.; Alborn, A.-M.; Nordborg, C.; Peterson, D.A.; Gage, F.H. Neurogenesis in the adult human hippocampus. Nat. Med. 1998, 4, 1313–1317. [Google Scholar] [CrossRef] [PubMed]
- Benjelloun-Touimi, S.; Jacque, C.M.; Derer, P.; De Vitry, F.; Maunoury, R.; Dupouey, P. Evidence that mouse astrocytes may be derived from the radial glia. An immunohistochemical study of the cerebellum in the normal and reeler mouse. J. Neuroimmunol. 1985, 9, 87–97. [Google Scholar] [CrossRef]
- Cameron, H.A.; Woolley, C.S.; McEwen, B.S.; Gould, E. Differentiation of newly born neurons and glia in the dentate gyrus of the adult rat. Neuroscience 1993, 56, 337–344. [Google Scholar] [CrossRef]
- Dulken, B.W.; Buckley, M.T.; Navarro Negredo, P.; Saligrama, N.; Cayrol, R.; Leeman, D.S.; George, B.M.; Boutet, S.C.; Hebestreit, K.; Pluvinage, J.V.; et al. Single-cell analysis reveals T cell infiltration in old neurogenic niches. Nature 2019, 571, 205–210. [Google Scholar] [CrossRef]
- Kornack, D.R.; Rakic, P. Continuation of neurogenesis in the hippocampus of the adult macaque monkey. Proc. Natl. Acad. Sci. USA 1999, 96, 5768–5773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommer, I.; Schachner, M. Monoclonal antibodies (O1 to O4) to oligodendrocyte cell surfaces: An immunocytological study in the central nervous system. Dev. Biol. 1981, 83, 311–327. [Google Scholar] [CrossRef]
- van Praag, H.; Kempermann, G.; Gage, F.H. Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus. Nat. Neurosci. 1999, 2, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.Z.; McKanna, J.A. Gliogenesis in postnatal rat optic nerve: LC1 + microglia and S100-beta + astrocytes. Brain Res. Dev. Brain Res. 1997, 101, 27–36. [Google Scholar] [CrossRef]
- Baptista, P.; Andrade, J.P. Adult Hippocampal Neurogenesis: Regulation and Possible Functional and Clinical Correlates. Front. Neuroanat. 2018, 12, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saraulli, D.; Costanzi, M.; Mastrorilli, V.; Farioli-Vecchioli, S. The Long Run: Neuroprotective Effects of Physical Exercise on Adult Neurogenesis from Youth to Old Age. Curr. Neuropharmacol. 2017, 15, 519–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sefiani, A.; Geoffroy, C.G. The Potential Role of Inflammation in Modulating Endogenous Hippocampal Neurogenesis After Spinal Cord Injury. Front. Neurosci. 2021, 15, 682259. [Google Scholar] [CrossRef]
- Eze, U.C.; Bhaduri, A.; Haeussler, M.; Nowakowski, T.J.; Kriegstein, A.R. Single-cell atlas of early human brain development highlights heterogeneity of human neuroepithelial cells and early radial glia. Nat. Neurosci. 2021, 24, 584–594. [Google Scholar] [CrossRef]
- Götz, M.; Hartfuss, E.; Malatesta, P. Radial glial cells as neuronal precursors: A new perspective on the correlation of morphology and lineage restriction in the developing cerebral cortex of mice. Brain Res. Bull. 2002, 57, 777–788. [Google Scholar] [CrossRef]
- Götz, M.; Nakafuku, M.; Petrik, D. Neurogenesis in the Developing and Adult Brain-Similarities and Key Differences. Cold Spring Harb. Perspect Biol. 2016, 8, a018853. [Google Scholar] [CrossRef]
- Malatesta, P.; Götz, M. Radial glia-from boring cables to stem cell stars. Development 2013, 140, 483–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, F.D.; Gauthier, A.S. Timing Is Everything: Making Neurons versus Glia in the Developing Cortex. Neuron 2007, 54, 357–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lago-Baldaia, I.; Fernandes, V.M.; Ackerman, S.D. More Than Mortar: Glia as Architects of Nervous System Development and Disease. Front. Cell Dev. Biol. 2020, 8, 611269. [Google Scholar] [CrossRef] [PubMed]
- Martins-Macedo, J.; Salgado, A.J.; Gomes, E.D.; Pinto, L. Adult brain cytogenesis in the context of mood disorders: From neurogenesis to the emergent role of gliogenesis. Neurosci. Biobehav. Rev. 2021, 131, 411–428. [Google Scholar] [CrossRef]
- Bicker, F.; Nardi, L.; Maier, J.; Vasic, V.; Schmeisser, M.J. Criss-crossing autism spectrum disorder and adult neurogenesis. J. Neurochem. 2021, 159, 452–478. [Google Scholar] [CrossRef]
- Castilla-Ortega, E.; Serrano, A.; Blanco, E.; Araos, P.; Suárez, J.; Pavón, F.J.; Rodríguez de Fonseca, F.; Santín, L.J. A place for the hippocampus in the cocaine addiction circuit: Potential roles for adult hippocampal neurogenesis. Neurosci. Biobehav. Rev. 2016, 66, 15–32. [Google Scholar] [CrossRef]
- Chen, P.; Chen, F.; Wu, Y.; Zhou, B. New Insights Into the Role of Aberrant Hippocampal Neurogenesis in Epilepsy. Front. Neurol. 2021, 12, 727065. [Google Scholar] [CrossRef]
- Iannitelli, A.; Quartini, A.; Tirassa, P.; Bersani, G. Schizophrenia and neurogenesis: A stem cell approach. Neurosci. Biobehav. Rev. 2017, 80, 414–442. [Google Scholar] [CrossRef]
- Peyton, L.; Oliveros, A.; Choi, D.-S.; Jang, M.-H. Hippocampal regenerative medicine: Neurogenic implications for addiction and mental disorders. Exp. Mol. Med. 2021, 53, 358–368. [Google Scholar] [CrossRef]
- Toda, T.; Parylak, S.L.; Linker, S.B.; Gage, F.H. The role of adult hippocampal neurogenesis in brain health and disease. Mol. Psychiatry 2019, 24, 67–87. [Google Scholar] [CrossRef]
- Malberg, J.E. Implications of adult hippocampal neurogenesis in antidepressant action. J. Psychiatry Neurosci. 2004, 29, 196–205. [Google Scholar] [PubMed]
- Morais, M.; Patrício, P.; Mateus-Pinheiro, A.; Alves, N.D.; Machado-Santos, A.R.; Correia, J.S.; Pereira, J.; Pinto, L.; Sousa, N.; Bessa, J.M. The modulation of adult neuroplasticity is involved in the mood-improving actions of atypical antipsychotics in an animal model of depression. Transl. Psychiatry 2017, 7, e1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tartt, A.N.; Mariani, M.B.; Hen, R.; Mann, J.J.; Boldrini, M. Dysregulation of adult hippocampal neuroplasticity in major depression: Pathogenesis and therapeutic implications. Mol. Psychiatry 2022, 27, 2689–2699. [Google Scholar] [CrossRef] [PubMed]
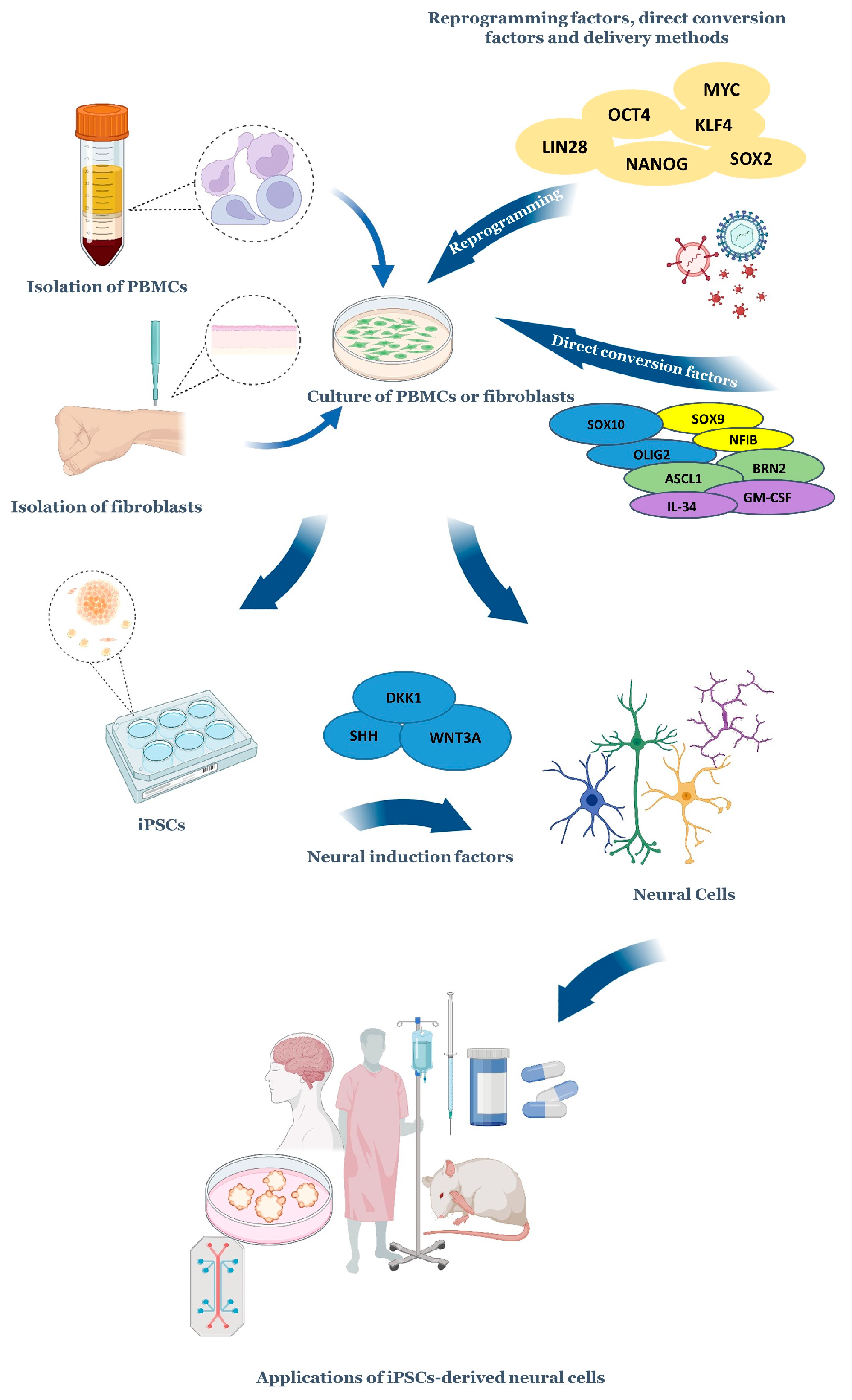
- Brooks, I.R.; Garrone, C.M.; Kerins, C.; Kiar, C.S.; Syntaka, S.; Xu, J.Z.; Spagnoli, F.M.; Watt, F.M. Functional genomics and the future of iPSCs in disease modeling. Stem Cell Rep. 2022, 17, 1033–1047. [Google Scholar] [CrossRef]
- Yamasaki, A.E.; Panopoulos, A.D.; Belmonte, J.C.I. Understanding the genetics behind complex human disease with large-scale iPSC collections. Genome Biol. 2017, 18, 135. [Google Scholar] [CrossRef] [Green Version]
- Fernando, M.B.; Ahfeldt, T.; Brennand, K.J. Modeling the complex genetic architectures of brain disease. Nat. Genet. 2020, 52, 363–369. [Google Scholar] [CrossRef]
- Vasan, L.; Park, E.; David, L.A.; Fleming, T.; Schuurmans, C. Direct Neuronal Reprogramming: Bridging the Gap Between Basic Science and Clinical Application. Front. Cell Dev. Biol. 2021, 9, 681087. [Google Scholar] [CrossRef]
- Mollinari, C.; Merlo, D. Direct Reprogramming of Somatic Cells to Neurons: Pros and Cons of Chemical Approach. Neurochem. Res. 2021, 46, 1330–1336. [Google Scholar] [CrossRef]
- Mandai, M.; Watanabe, A.; Kurimoto, Y.; Hirami, Y.; Morinaga, C.; Daimon, T.; Fujihara, M.; Akimaru, H.; Sakai, N.; Shibata, Y.; et al. Autologous Induced Stem-Cell–Derived Retinal Cells for Macular Degeneration. N. Engl. J. Med. 2017, 376, 1038–1046. [Google Scholar] [CrossRef]
- Takagi, S.; Mandai, M.; Gocho, K.; Hirami, Y.; Yamamoto, M.; Fujihara, M.; Sugita, S.; Kurimoto, Y.; Takahashi, M. Evaluation of Transplanted Autologous Induced Pluripotent Stem Cell-Derived Retinal Pigment Epithelium in Exudative Age-Related Macular Degeneration. Ophthalmol. Retin. 2019, 3, 850–859. [Google Scholar] [CrossRef]
- Schweitzer, J.S.; Song, B.; Herrington, T.M.; Park, T.-Y.; Lee, N.; Ko, S.; Jeon, J.; Cha, Y.; Kim, K.; Li, Q.; et al. Personalized iPSC-Derived Dopamine Progenitor Cells for Parkinson’s Disease. N. Engl. J. Med. 2020, 382, 1926–1932. [Google Scholar] [CrossRef] [PubMed]
- Marchetto, M.C.; Gage, F.H. Cell-Reprogramming Technology and Neuroscience; Whalley, K., Ed.; Nature Reviews; Nature Publishing Group: Berlin, Germany, 2015. [Google Scholar]
- Kristofova, M.; Ori, A.; Wang, Z.-Q. Multifaceted Microcephaly-Related Gene MCPH1. Cells 2022, 11, 275. [Google Scholar] [CrossRef] [PubMed]
- Roll, L.; Lessmann, K.; Brüstle, O.; Faissner, A. Cerebral Organoids Maintain the Expression of Neural Stem Cell-Associated Glycoepitopes and Extracellular Matrix. Cells 2022, 11, 760. [Google Scholar] [CrossRef]
- Kumar, V.; Goutam, R.S.; Umair, Z.; Park, S.; Lee, U.; Kim, J. Foxd4l1.1 Negatively Regulates Chordin Transcription in Neuroectoderm of Xenopus Gastrula. Cells 2021, 10, 2779. [Google Scholar] [CrossRef] [PubMed]
- Rastoldo, G.; Marouane, E.; El-Mahmoudi, N.; Péricat, D.; Watabe, I.; Lapotre, A.; Tonetto, A.; López-Juárez, A.; El-Ahmadi, A.; Caron, P.; et al. L-Thyroxine Improves Vestibular Compensation in a Rat Model of Acute Peripheral Vestibulopathy: Cellular and Behavioral Aspects. Cells 2022, 11, 684. [Google Scholar] [CrossRef]
- Marouane, E.; El Mahmoudi, N.; Rastoldo, G.; Péricat, D.; Watabe, I.; Lapôtre, A.; Tonetto, A.; Xavier, F.; Dumas, O.; Chabbert, C.; et al. Sensorimotor Rehabilitation Promotes Vestibular Compensation in a Rodent Model of Acute Peripheral Vestibulopathy by Promoting Microgliogenesis in the Deafferented Vestibular Nuclei. Cells 2021, 10, 3377. [Google Scholar] [CrossRef]
- Machado-Santos, A.R.; Loureiro-Campos, E.; Patrício, P.; Araújo, B.; Alves, N.D.; Mateus-Pinheiro, A.; Correia, J.S.; Morais, M.; Bessa, J.M.; Sousa, N.; et al. Beyond New Neurons in the Adult Hippocampus: Imipramine Acts as a Pro-Astrogliogenic Factor and Rescues Cognitive Impairments Induced by Stress Exposure. Cells 2022, 11, 390. [Google Scholar] [CrossRef]
- Butt, A.M.; Rivera, A.D.; Fulton, D.; Azim, K. Targeting the Subventricular Zone to Promote Myelin Repair in the Aging Brain. Cells 2022, 11, 1809. [Google Scholar] [CrossRef] [PubMed]
- Essa, H.; Peyton, L.; Hasan, W.; León, B.E.; Choi, D.S. Implication of Adult Hippocampal Neurogenesis in Alzheimer’s Disease and Potential Therapeutic Approaches. Cells 2022, 11, 286. [Google Scholar] [CrossRef]
- Willinger, Y.; Turgeman, G. Interleukine-17 Modulates Neurogenesis and Behavior Following Exposure to Trauma in Mice. Cells 2022, 11, 343. [Google Scholar] [CrossRef]
- Yoo, S.; Stremlau, M.; Pinto, A.; Woo, H.; Curtis, O.; van Praag, H. Effects of Combined Anti-Hypertensive and Statin Treatment on Memory, Fear Extinction, Adult Neurogenesis, and Angiogenesis in Adult and Middle-Aged Mice. Cells 2021, 10, 1778. [Google Scholar] [CrossRef] [PubMed]
- Mayerl, S.; Alcaide Martin, A.; Bauer, R.; Schwaninger, M.; Heuer, H.; Ffrench-Constant, C. Distinct Actions of the Thyroid Hormone Transporters Mct8 and Oatp1c1 in Murine Adult Hippocampal Neurogenesis. Cells 2022, 11, 524. [Google Scholar] [CrossRef] [PubMed]
- Soares, L.C.; Al-Dalahmah, O.; Hillis, J.; Young, C.C.; Asbed, I.; Sakaguchi, M.; O’Neill, E.; Szele, F.G. Novel Galectin-3 Roles in Neurogenesis, Inflammation and Neurological Diseases. Cells 2021, 10, 3047. [Google Scholar] [CrossRef]
- Ali, A.A.H.; von Gall, C. Adult Neurogenesis under Control of the Circadian System. Cells 2022, 11, 764. [Google Scholar] [CrossRef] [PubMed]
- Bartkowska, K.; Tepper, B.; Turlejski, K.; Djavadian, R. Postnatal and Adult Neurogenesis in Mammals, Including Marsupials. Cells 2022, 11, 2735. [Google Scholar] [CrossRef] [PubMed]
- Monfared, R.V.; Alhassen, W.; Truong, T.M.; Gonzales, M.A.M.; Vachirakorntong, V.; Chen, S.; Baldi, P.; Civelli, O.; Alachkar, A. Transcriptome Profiling of Dysregulated GPCRs Reveals Overlapping Patterns across Psychiatric Disorders and Age-Disease Interactions. Cells 2021, 10, 2967. [Google Scholar] [CrossRef]
- Bravo, J.; Ribeiro, I.; Terceiro, A.F.; Andrade, E.B.; Portugal, C.C.; Lopes, I.M.; Azevedo, M.M.; Sousa, M.; Lopes, C.D.F.; Lobo, A.C.; et al. Neuron-Microglia Contact-Dependent Mechanisms Attenuate Methamphetamine-Induced Microglia Reactivity and Enhance Neuronal Plasticity. Cells 2022, 11, 355. [Google Scholar] [CrossRef]
- Loeffler-Wirth, H.; Hopp, L.; Schmidt, M.; Zakharyan, R.; Arakelyan, A.; Binder, H. The Transcriptome and Methylome of the Developing and Aging Brain and Their Relations to Gliomas and Psychological Disorders. Cells 2022, 11, 362. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaz, A.; Ribeiro, I.; Pinto, L. Frontiers in Neurogenesis. Cells 2022, 11, 3567. https://doi.org/10.3390/cells11223567
Vaz A, Ribeiro I, Pinto L. Frontiers in Neurogenesis. Cells. 2022; 11(22):3567. https://doi.org/10.3390/cells11223567
Chicago/Turabian StyleVaz, Andreia, Inês Ribeiro, and Luísa Pinto. 2022. "Frontiers in Neurogenesis" Cells 11, no. 22: 3567. https://doi.org/10.3390/cells11223567
APA StyleVaz, A., Ribeiro, I., & Pinto, L. (2022). Frontiers in Neurogenesis. Cells, 11(22), 3567. https://doi.org/10.3390/cells11223567