




Age, Education Years, and Biochemical Factors Are Associated with Selective Neuronal Changes in the Elderly Hippocampus


Abstract
1. Introduction
2. Materials and Methods
2.1. Brain Samples
2.2. Tissue Processing
2.3. Case Eligibility
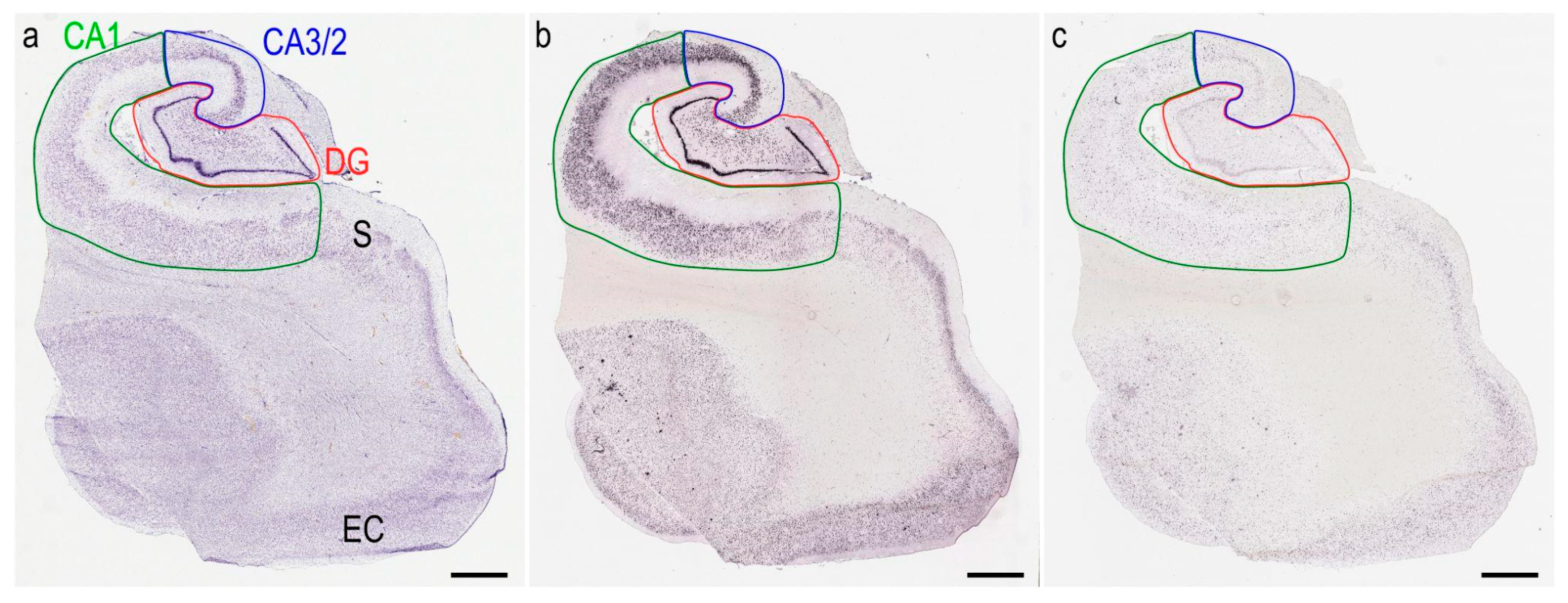
2.4. Image Analysis
2.5. Statistics
3. Results
3.1. Effects of Age and Sex on Hippocampal Neuron-Specific Markers
3.2. Effects of Education Years on Hippocampal Neuron-Specific Markers
3.3. Associations of Biochemical Factors with Hippocampal Neuron-Specific Markers
4. Discussion
4.1. Age and Sex Effects on Glutamatergic and GABAergic Markers
4.2. Associations of Biological Sex and Education Years with VGLUT1 and GAT1 Expression
4.3. Association of Neuropathological Proteins and VGLUT1 Expression
4.4. IL-7 Levels Are Associated with VGLUT1 Expression
4.5. IFNg and BDNF Levels Are Associated with GAT1 Expression
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hc Subfield | Correlation | VGLUT1 | GAT1 | ||||
|---|---|---|---|---|---|---|---|
| Female | Male | Both | Female | Male | Both | ||
| DG | r | −0.13 | −0.56 | −0.33 | 0.66 | 0.08 | 0.29 |
| p | 0.72 | 0.12 | 0.16 | 0.036 * | 0.84 | 0.23 | |
| CA3/2 | r | 0.15 | −0.43 | −0.21 | 0.76 | −0.3 | 0.07 |
| p | 0.68 | 0.24 | 0.39 | 0.01 * | 0.43 | 0.77 | |
| CA1 | r | 0.14 | 0.01 | 0.05 | 0.56 | −0.21 | 0.11 |
| p | 0.69 | 0.99 | 0.82 | 0.09 | 0.59 | 0.66 | |
| Total Hc | r | 0.12 | −0.26 | −0.09 | 0.7 | −0.15 | 0.17 |
| p | 0.74 | 0.51 | 0.71 | 0.02 * | 0.69 | 0.48 | |
References
- Niccoli, T.; Partridge, L. Ageing as a Risk Factor for Disease. Curr. Biol. 2012, 22, R741–R752. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Dan, X.; Babbar, M.; Wei, Y.; Hasselbalch, S.G.; Croteau, D.L.; Bohr, V.A. Ageing as a risk factor for neurodegenerative disease. Nat. Rev. Neurol. 2019, 15, 565–581. [Google Scholar] [CrossRef] [PubMed]
- Carmona, J.J.; Michan, S. Biology of Healthy Aging and Longevity. Rev. Investig. Clin. 2016, 68, 7–16. [Google Scholar]
- Todhunter, M.E.; Sayaman, R.W.; Miyano, M.; LaBarge, M.A. Tissue aging: The integration of collective and variant responses of cells to entropic forces over time. Curr. Opin. Cell Biol. 2018, 54, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Schaum, N.; Lehallier, B.; Hahn, O.; Pálovics, R.; Hosseinzadeh, S.; Lee, S.E.; Sit, R.; Lee, D.P.; Losada, P.M.; Zardeneta, M.E.; et al. Ageing Hallmarks Exhibit Organ-Specific Temporal Signatures. Nature 2020, 583, 596–602. [Google Scholar] [CrossRef] [PubMed]
- West, M.J. Regionally Specific Loss of Neurons in the Aging Human Hippocampus. Neurobiol. Aging 1993, 14, 287–293. [Google Scholar] [CrossRef]
- Pakkenberg, B.; Gundersen, H.J.G. Neocortical Neuron Number in Humans: Effect of Sex and Age. J. Comp. Neurol. 1997, 384, 312–320. [Google Scholar] [CrossRef]
- Andersen, B.B.; Gundersen, H.J.G.; Pakkenberg, B. Aging of the Human Cerebellum: A Stereological Study. J. Comp. Neurol. 2003, 466, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.E.; Rapp, P.R.; McKay, H.M.; Roberts, J.A.; Tuszynski, M.H. Memory Impairment in Aged Primates Is Associated with Focal Death of Cortical Neurons and Atrophy of Subcortical Neurons. J. Neurosci. 2004, 24, 4373–4381. [Google Scholar] [CrossRef] [PubMed]
- Yates, M.A.; Markham, J.A.; Anderson, S.E.; Morris, J.R.; Juraska, J.M. Regional Variability in Age-Related Loss of Neurons from the Primary Visual Cortex and Medial Prefrontal Cortex of Male and Female Rats. Brain Res. 2008, 1218, 1–12. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fiuza, F.P.; Aquino, A.C.Q.; Câmara, D.A.; Cavalcanti, J.R.L.P.; Nascimento Júnior, E.S.; Lima, R.H.; Engelberth, R.C.G.J.; Cavalcante, J.S. Region-Specific Glial Hyperplasia and Neuronal Stability of Rat Lateral Geniculate Nucleus during Aging. Exp. Gerontol. 2017, 100, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Cabello, C.R.; Thune, J.J.; Pakkenberg, H.; Pakkenberg, B. Ageing of Substantia Nigra in Humans: Cell Loss May Be Compensated by Hypertrophy. Neuropathol. Appl. Neurobiol. 2002, 28, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Fiuza, F.P.; Silva, K.D.A.; Pessoa, R.A.; Pontes, A.L.B.; Cavalcanti, R.L.P.; Pires, R.S.; Soares, J.G.; Nascimento Júnior, E.S.; Costa, M.S.M.O.; Engelberth, R.C.G.J.; et al. Age-Related Changes in Neurochemical Components and Retinal Projections of Rat Intergeniculate Leaflet. Age 2016, 38, 4. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lamerand, S.; Shahidehpour, R.; Ayala, I.; Gefen, T.; Mesulam, M.-M.; Bigio, E.; Geula, C. Calbindin-D28K, Parvalbumin, and Calretinin in Young and Aged Human Locus Coeruleus. Neurobiol. Aging 2020, 94, 243–249. [Google Scholar] [CrossRef]
- Fiuza, F.P.; Queiroz, J.P.G.; Aquino, A.C.Q.; Câmara, D.A.; Brandão, L.E.M.; Lima, R.H.; Cavalcanti, J.R.L.P.; Engelberth, R.C.G.J.; Cavalcante, J.S. Aging Alters Daily and Regional Calretinin Neuronal Expression in the Rat Non-Image Forming Visual Thalamus. Front. Aging Neurosci. 2021, 13, 613305. [Google Scholar] [CrossRef]
- Stern, Y. Cognitive Reserve in Ageing and Alzheimer’s Disease. Lancet Neurol. 2012, 11, 1006–1012. [Google Scholar] [CrossRef]
- Armstrong, N.M.; An, Y.; Beason-Held, L.; Doshi, J.; Erus, G.; Ferrucci, L.; Davatzikos, C.; Resnick, S.M. Sex Differences in Brain Aging and Predictors of Neurodegeneration in Cognitively Healthy Older Adults. Neurobiol. Aging 2019, 81, 146–156. [Google Scholar] [CrossRef]
- Barth, E.; Srivastava, A.; Stojiljkovic, M.; Frahm, C.; Axer, H.; Witte, O.W.; Marz, M. Conserved Aging-Related Signatures of Senescence and Inflammation in Different Tissues and Species. Aging 2019, 11, 8556–8572. [Google Scholar] [CrossRef]
- Bettio, L.E.B.; Rajendran, L.; Gil-Mohapel, J. The Effects of Aging in the Hippocampus and Cognitive Decline. Neurosci. Biobehav. Rev. 2017, 79, 66–86. [Google Scholar] [CrossRef]
- Eichenbaum, H. The Hippocampus and Mechanisms of Declarative Memory. Behav. Brain Res. 1999, 103, 123–133. [Google Scholar] [CrossRef]
- Eichenbaum, H.; Cohen, N.J. Can We Reconcile the Declarative Memory and Spatial Navigation Views on Hippocampal Function? Neuron 2014, 83, 764–770. [Google Scholar] [CrossRef] [PubMed]
- Amunts, K.; Kedo, O.; Kindler, M.; Pieperhoff, P.; Mohlberg, H.; Shah, N.J.; Habel, U.; Schneider, F.; Zilles, K. Cytoarchitectonic Mapping of the Human Amygdala, Hippocampal Region and Entorhinal Cortex: Intersubject Variability and Probability Maps. Anat. Embryol. 2005, 210, 343–352. [Google Scholar] [CrossRef]
- Palomero-Gallagher, N.; Kedo, O.; Mohlberg, H.; Zilles, K.; Amunts, K. Multimodal Mapping and Analysis of the Cyto- and Receptorarchitecture of the Human Hippocampus. Brain Struct. Funct. 2020, 225, 881–907. [Google Scholar] [CrossRef] [PubMed]
- Nobis, L.; Manohar, S.G.; Smith, S.M.; Alfaro-Almagro, F.; Jenkinson, M.; Mackay, C.E.; Husain, M. Hippocampal Volume across Age: Nomograms Derived from over 19,700 People in UK Biobank. Neuroimage Clin. 2019, 23, 101904. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, J.; Ren, J.; Sun, S.; Ma, S.; Zhang, W.; Yu, Y.; Cai, Y.; Yan, K.; Li, W.; et al. Single-Nucleus Transcriptomic Landscape of Primate Hippocampal Aging. Protein Cell 2021, 12, 695–716. [Google Scholar] [CrossRef] [PubMed]
- Booker, S.A.; Vida, I. Morphological Diversity and Connectivity of Hippocampal Interneurons. Cell Tissue Res. 2018, 373, 619–641. [Google Scholar] [CrossRef]
- Takamori, S.; Rhee, J.S.; Rosenmund, C.; Jahn, R. Identification of a Vesicular Glutamate Transporter That Defines a Glutamatergic Phenotype in Neurons. Nature 2000, 407, 189–194. [Google Scholar] [CrossRef]
- Vigneault, É.; Poirel, O.; Riad, M.; Prud’homme, J.; Dumas, S.; Turecki, G.; Fasano, C.; Mechawar, N.; El Mestikawy, S. Distribution of Vesicular Glutamate Transporters in the Human Brain. Front. Neuroanat. 2015, 9, 23. [Google Scholar] [CrossRef]
- Jung, H.Y.; Yoo, D.Y.; Park, J.H.; Kim, J.W.; Chung, J.Y.; Kim, D.W.; Won, M.-H.; Yoon, Y.S.; Hwang, I.K. Age-Dependent Changes in Vesicular Glutamate Transporter 1 and 2 Expression in the Gerbil Hippocampus. Mol. Med. Rep. 2018, 17, 6465–6471. [Google Scholar] [CrossRef]
- Rozycka, A.; Charzynska, A.; Misiewicz, Z.; Stepniewski, T.M.; Sobolewska, A.; Kossut, M.; Liguz-Lecznar, M. Glutamate, GABA, and Presynaptic Markers Involved in Neurotransmission Are Differently Affected by Age in Distinct Mouse Brain Regions. ACS Chem. Neurosci. 2019, 10, 4449–4461. [Google Scholar] [CrossRef]
- Roalf, D.R.; Sydnor, V.J.; Woods, M.; Wolk, D.A.; Scott, J.C.; Reddy, R.; Moberg, P.J. A Quantitative Meta-Analysis of Brain Glutamate Metabolites in Aging. Neurobiol. Aging 2020, 95, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Gasiorowska, A.; Wydrych, M.; Drapich, P.; Zadrozny, M.; Steczkowska, M.; Niewiadomski, W.; Niewiadomska, G. The Biology and Pathobiology of Glutamatergic, Cholinergic, and Dopaminergic Signaling in the Aging Brain. Front. Aging Neurosci. 2021, 13, 654931. [Google Scholar] [CrossRef] [PubMed]
- Cox, M.F.; Hascup, E.R.; Bartke, A.; Hascup, K.N. Friend or Foe? Defining the Role of Glutamate in Aging and Alzheimer’s Disease. Front. Aging 2022, 3, 929474. [Google Scholar] [CrossRef] [PubMed]
- Sahara, S.; Yanagawa, Y.; O’Leary, D.D.M.; Stevens, C.F. The Fraction of Cortical GABAergic Neurons Is Constant from Near the Start of Cortical Neurogenesis to Adulthood. J. Neurosci. 2012, 32, 4755–4761. [Google Scholar] [CrossRef]
- Scimemi, A. Structure, Function, and Plasticity of GABA Transporters. Front. Cell. Neurosci. 2014, 8, 161. [Google Scholar] [CrossRef]
- Pandya, M.; Palpagama, T.H.; Turner, C.; Waldvogel, H.J.; Faull, R.L.; Kwakowsky, A. Sex- and Age-Related Changes in GABA Signaling Components in the Human Cortex. Biol. Sex Differ. 2019, 10, 5. [Google Scholar] [CrossRef]
- Ethiraj, J.; Palpagama, T.H.; Turner, C.; van der Werf, B.; Waldvogel, H.J.; Faull, R.L.M.; Kwakowsky, A. The Effect of Age and Sex on the Expression of GABA Signaling Components in the Human Hippocampus and Entorhinal Cortex. Sci. Rep. 2021, 11, 21470. [Google Scholar] [CrossRef]
- Kukull, W.A.; Higdon, R.; Bowen, J.D.; McCormick, W.C.; Teri, L.; Schellenberg, G.D.; van Belle, G.; Jolley, L.; Larson, E.B. Dementia and Alzheimer Disease Incidence: A Prospective Cohort Study. Arch. Neurol. 2002, 59, 1737–1746. [Google Scholar] [CrossRef]
- Miller, J.A.; Guillozet-Bongaarts, A.; Gibbons, L.E.; Postupna, N.; Renz, A.; Beller, A.E.; Sunkin, S.M.; Ng, L.; Rose, S.E.; Smith, K.A.; et al. Neuropathological and Transcriptomic Characteristics of the Aged Brain. eLife 2017, 6, e31126. [Google Scholar] [CrossRef]
- Teng, E.L.; Hasegawa, K.; Homma, A.; Imai, Y.; Larson, E.; Graves, A.; Sugimoto, K.; Yamaguchi, T.; Sasaki, H.; Chiu, D.; et al. The Cognitive Abilities Screening Instrument (CASI): A Practical Test for Cross-Cultural Epidemiological Studies of Dementia. Int. Psychogeriatr. 1994, 6, 45–58. [Google Scholar] [CrossRef]
- Schroeder, A.; Mueller, O.; Stocker, S.; Salowsky, R.; Leiber, M.; Gassmann, M.; Lightfoot, S.; Menzel, W.; Granzow, M.; Ragg, T. The RIN: An RNA Integrity Number for Assigning Integrity Values to RNA Measurements. BMC Mol. Biol. 2006, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Stoner, R.; Chow, M.L.; Boyle, M.P.; Sunkin, S.M.; Mouton, P.R.; Roy, S.; Wynshaw-Boris, A.; Colamarino, S.A.; Lein, E.S.; Courchesne, E. Patches of Disorganization in the Neocortex of Children with Autism. N. Engl. J. Med. 2014, 27, 1209–1219. [Google Scholar] [CrossRef] [PubMed]
- Highet, B.; Parker, R.; Faull, R.L.M.; Curtis, M.A.; Ryan, B. RNA Quality in Post-Mortem Human Brain Tissue Is Affected by Alzheimer’s Disease. Front. Mol. Neurosci. 2021, 14, 780352. [Google Scholar] [CrossRef] [PubMed]
- Lein, E.S.; Hawrylycz, M.J.; Ao, N.; Ayres, M.; Bensinger, A.; Bernard, A.; Boe, A.F.; Boguski, M.S.; Brockway, K.S.; Byrnes, E.J.; et al. Genome-Wide Atlas of Gene Expression in the Adult Mouse Brain. Nature 2007, 445, 168–176. [Google Scholar] [CrossRef]
- Villar-Conde, S.; Astillero-Lopez, V.; Gonzalez-Rodriguez, M.; Villanueva-Anguita, P.; Saiz-Sanchez, D.; Martinez-Marcos, A.; Flores-Cuadrado, A.; Ubeda-Bañon, I. The Human Hippocampus in Parkinson’s Disease: An Integrative Stereological and Proteomic Study. J. Park. Dis. 2021, 11, 1345–1365. [Google Scholar] [CrossRef]
- Gonzalez-Rodriguez, M.; Villar-Conde, S.; Astillero-Lopez, V.; Villanueva-Anguita, P.; Ubeda-Banon, I.; Flores-Cuadrado, A.; Martinez-Marcos, A.; Saiz-Sanchez, D. Neurodegeneration and Astrogliosis in the Human CA1 Hippocampal Subfield Are Related to Hsp90ab1 and Bag3 in Alzheimer’s Disease. Int. J. Mol. Sci. 2022, 23, 165. [Google Scholar] [CrossRef]
- Howard, V.; Reed, M.G. Unbiased Stereology: Three-Dimensional Measurement in Microscopy (Advanced Methods), 2nd ed.; Garland Science: New York, NY, USA, 2005; p. 53. [Google Scholar]
- Bonasera, S.J.; Arikkath, J.; Boska, M.D.; Chaudoin, T.R.; DeKorver, N.W.; Goulding, E.H.; Hoke, T.A.; Mojtahedzedah, V.; Reyelts, C.D.; Sajja, B.; et al. Age-Related Changes in Cerebellar and Hypothalamic Function Accompany Non-Microglial Immune Gene Expression, Altered Synapse Organization, and Excitatory Amino Acid Neurotransmission Deficits. Aging 2016, 8, 2153–2181. [Google Scholar] [CrossRef]
- Kashani, A.; Lepicard, E.; Poirel, O.; Videau, C.; David, J.P.; Fallet-Bianco, C.; Simon, A.; Delacourte, A.; Giros, B.; Epelbaum, J.; et al. Loss of VGLUT1 and VGLUT2 in the Prefrontal Cortex Is Correlated with Cognitive Decline in Alzheimer Disease. Neurobiol. Aging 2008, 29, 1619–1630. [Google Scholar] [CrossRef]
- Poirel, O.; Mella, S.; Videau, C.; Ramet, L.; Davoli, M.A.; Herzog, E.; Katsel, P.; Mechawar, N.; Haroutunian, V.; Epelbaum, J.; et al. Moderate Decline in Select Synaptic Markers in the Prefrontal Cortex (BA9) of Patients with Alzheimer’s Disease at Various Cognitive Stages. Sci. Rep. 2018, 8, 938. [Google Scholar] [CrossRef]
- Liao, C.; Han, Q.; Ma, Y.; Su, B. Age-Related Gene Expression Change of GABAergic System in Visual Cortex of Rhesus Macaque. Gene 2016, 590, 227–233. [Google Scholar] [CrossRef]
- Sumner, P.; Edden, R.A.E.; Bompas, A.; Evans, C.J.; Singh, K.D. More GABA, Less Distraction: A Neurochemical Predictor of Motor Decision Speed. Nat. Neurosci. 2010, 13, 825–827. [Google Scholar] [CrossRef] [PubMed]
- Rozycka, A.; Liguz-Lecznar, M. The Space Where Aging Acts: Focus on the GABAergic Synapse. Aging Cell 2017, 16, 634–643. [Google Scholar] [CrossRef]
- Barron, H.C. Neural Inhibition for Continual Learning and Memory. Curr. Opin. Neurobiol. 2021, 67, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Spurny, B.; Seiger, R.; Moser, P.; Vanicek, T.; Reed, M.B.; Heckova, E.; Michenthaler, P.; Basaran, A.; Gryglewski, G.; Klöbl, M.; et al. Hippocampal GABA Levels Correlate with Retrieval Performance in an Associative Learning Paradigm. NeuroImage 2020, 204, 116244. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Balado, J.; Ycaza Herrera, A.; Igwe, K.; Klem, L.; Buyukturkoglu, K.; Irimia, A.; Chen, L.; Guo, J.; Brickman, A.M.; Eich, T.S. Reduced Hippocampal GABA+ Is Associated With Poorer Episodic Memory in Healthy Older Women: A Pilot Study. Front. Behav. Neurosci. 2021, 15, 695416. [Google Scholar] [CrossRef] [PubMed]
- Ko, K.; Yi, D.; Byun, M.S.; Lee, J.H.; Jeon, S.Y.; Kim, W.J.; Byeon, G.; Sung, K.; Han, D.; Lee, Y.; et al. Cognitive Reserve Proxies, Alzheimer Pathologies, and Cognition. Neurobiol. Aging 2022, 110, 88–95. [Google Scholar] [CrossRef]
- Weiland, N.G.; Orchinik, M. Specific subunit mRNAs of the GABAA receptor are regulated by progesterone in subfields of the hippocampus. Mol. Brain Res. 1995, 32, 271–278. [Google Scholar] [CrossRef]
- Murphy, D.D.; Cole, N.B.; Greenberger, V.; Segal, M. Estradiol Increases Dendritic Spine Density by Reducing GABA Neurotransmission in Hippocampal Neurons. J. Neurosci. 1998, 18, 2550–2559. [Google Scholar] [CrossRef]
- Huijbers, W.; Schultz, A.P.; Papp, K.V.; LaPoint, M.R.; Hanseeuw, B.; Chhatwal, J.P.; Hedden, T.; Johnson, K.A.; Sperling, R.A. Tau Accumulation in Clinically Normal Older Adults Is Associated with Hippocampal Hyperactivity. J. Neurosci. 2019, 39, 548–556. [Google Scholar] [CrossRef]
- Hunsberger, H.C.; Rudy, C.C.; Batten, S.R.; Gerhardt, G.A.; Reed, M.N. P301L Tau Expression Affects Glutamate Release and Clearance in the Hippocampal Trisynaptic Pathway. J. Neurochem. 2014, 132, 169–182. [Google Scholar] [CrossRef]
- Hebron, M.L.; Javidnia, M.; Moussa, C.E.-H. Tau Clearance Improves Astrocytic Function and Brain Glutamate-Glutamine Cycle. J. Neurol. Sci. 2018, 391, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Decker, J.M.; Krüger, L.; Sydow, A.; Dennissen, F.J.; Siskova, Z.; Mandelkow, E.; Mandelkow, E.-M. The Tau/A152T Mutation, a Risk Factor for Frontotemporal-Spectrum Disorders, Leads to NR2B Receptor-Mediated Excitotoxicity. EMBO Rep. 2016, 17, 552–569. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, T.; Stein, L.; Thomas, R.; Djukic, B.; Taneja, P.; Knox, J.; Vossel, K.; Mucke, L. Phosphorylation of Tau at Y18, but Not Tau-Fyn Binding, Is Required for Tau to Modulate NMDA Receptor-Dependent Excitotoxicity in Primary Neuronal Culture. Mol. Neurodegener. 2017, 12, 41. [Google Scholar] [CrossRef]
- Timmer, N.M.; Metaxas, A.; van der Stelt, I.; Kluijtmans, L.A.J.; van Berckel, B.N.; Verbeek, M.M. Cerebral Level of VGlut1 Is Increased and Level of Glycine Is Decreased in TgSwDI Mice. J. Alzheimer’s Dis. 2014, 39, 89–101. [Google Scholar] [CrossRef]
- Hascup, E.R.; Broderick, S.O.; Russell, M.K.; Fang, Y.; Bartke, A.; Boger, H.A.; Hascup, K.N. Diet-Induced Insulin Resistance Elevates Hippocampal Glutamate as Well as VGLUT1 and GFAP Expression in AβPP/PS1 Mice. J. Neurochem. 2018, 148, 219–237. [Google Scholar] [CrossRef]
- Hascup, K.N.; Findley, C.A.; Sime, L.N.; Hascup, E.R. Hippocampal Alterations in Glutamatergic Signaling during Amyloid Progression in AβPP/PS1 Mice. Sci. Rep. 2020, 10, 14503. [Google Scholar] [CrossRef] [PubMed]
- Fogel, H.; Frere, S.; Segev, O.; Bharill, S.; Shapira, I.; Gazit, N.; O’Malley, T.; Slomowitz, E.; Berdichevsky, Y.; Walsh, D.M.; et al. APP Homodimers Transduce an Amyloid-β-Mediated Increase in Release Probability at Excitatory Synapses. Cell Rep. 2014, 7, 1560–1576. [Google Scholar] [CrossRef] [PubMed]
- Hascup, K.N.; Hascup, E.R. Soluble Amyloid-Β42 Stimulates Glutamate Release through Activation of the A7 Nicotinic Acetylcholine Receptor. J. Alzheimers Dis. 2016, 53, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Sokolow, S.; Luu, S.H.; Nandy, K.; Miller, C.A.; Vinters, H.V.; Poon, W.W.; Gylys, K.H. Preferential Accumulation of Amyloid-Beta in Presynaptic Glutamatergic Terminals (VGluT1 and VGluT2) in Alzheimer’s Disease Cortex. Neurobiol. Dis. 2012, 45, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Busche, M.A.; Eichhoff, G.; Adelsberger, H.; Abramowski, D.; Wiederhold, K.-H.; Haass, C.; Staufenbiel, M.; Konnerth, A.; Garaschuk, O. Clusters of Hyperactive Neurons Near Amyloid Plaques in a Mouse Model of Alzheimer’s Disease. Science 2008, 321, 1686–1689. [Google Scholar] [CrossRef]
- Kirvell, S.L.; Esiri, M.; Francis, P.T. Down-Regulation of Vesicular Glutamate Transporters Precedes Cell Loss and Pathology in Alzheimer’s Disease. J. Neurochem. 2006, 98, 939–950. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Perdigon, M.; Tordera, R.M.; Gil-Bea, F.J.; Gerenu, G.; Ramirez, M.J.; Solas, M. Down-Regulation of Glutamatergic Terminals (VGLUT1) Driven by Aβ in Alzheimer’s Disease. Hippocampus 2016, 26, 1303–1312. [Google Scholar] [CrossRef] [PubMed]
- Araujo, D.M.; Cotman, C.W. Trophic Effects of Interleukin-4, -7 and -8 on Hippocampal Neuronal Cultures: Potential Involvement of Glial-Derived Factors. Brain Res. 1993, 600, 49–55. [Google Scholar] [CrossRef]
- Andrew, D.; Aspinall, R. Age-Associated Thymic Atrophy Is Linked to a Decline in IL-7 Production. Exp. Gerontol. 2002, 37, 455–463. [Google Scholar] [CrossRef] [PubMed]
- De Simone, R.; Levi, G.; Aloisi, F. Interferon γ gene expression in rat central nervous system glial cells. Cytokine 1998, 10, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Flood, L.; Korol, S.V.; Ekselius, L.; Birnir, B.; Jin, Z. Interferon-γ Potentiates GABAA Receptor-Mediated Inhibitory Currents in Rat Hippocampal CA1 Pyramidal Neurons. J. Neuroimmunol. 2019, 337, 577050. [Google Scholar] [CrossRef]
- Janach, G.M.S.; Reetz, O.; Döhne, N.; Stadler, K.; Grosser, S.; Byvaltcev, E.; Bräuer, A.U.; Strauss, U. Interferon-γ Acutely Augments Inhibition of Neocortical Layer 5 Pyramidal Neurons. J. Neuroinflamm. 2020, 17, 69. [Google Scholar] [CrossRef]
- Filiano, A.J.; Xu, Y.; Tustison, N.J.; Marsh, R.L.; Baker, W.; Smirnov, I.; Overall, C.C.; Gadani, S.P.; Turner, S.D.; Weng, Z.; et al. Unexpected Role of Interferon-γ in Regulating Neuronal Connectivity and Social Behaviour. Nature 2016, 535, 425–429. [Google Scholar] [CrossRef]
- Colucci-D’Amato, L.; Speranza, L.; Volpicelli, F. Neurotrophic Factor BDNF, Physiological Functions and Therapeutic Potential in Depression, Neurodegeneration and Brain Cancer. Int. J. Mol. Sci. 2020, 21, 7777. [Google Scholar] [CrossRef]
- Buchman, A.S.; Yu, L.; Boyle, P.A.; Schneider, J.A.; De Jager, P.L.; Bennett, D.A. Higher Brain BDNF Gene Expression Is Associated with Slower Cognitive Decline in Older Adults. Neurology 2016, 86, 735–741. [Google Scholar] [CrossRef]
- Kim, J.; Lee, S.; Kang, S.; Kim, S.-H.; Kim, J.-C.; Yang, M.; Moon, C. Brain-Derived Neurotropic Factor and GABAergic Transmission in Neurodegeneration and Neuroregeneration. Neural Regen. Res. 2017, 12, 1733–1741. [Google Scholar] [CrossRef] [PubMed]
- Ohba, S.; Ikeda, T.; Ikegaya, Y.; Nishiyama, N.; Matsuki, N.; Yamada, M.K. BDNF Locally Potentiates GABAergic Presynaptic Machineries: Target-Selective Circuit Inhibition. Cereb. Cortex 2005, 15, 291–298. [Google Scholar] [CrossRef]
- Swanwick, C.C.; Murthy, N.R.; Kapur, J. Activity-Dependent Scaling of GABAergic Synapse Strength Is Regulated by Brain-Derived Neurotrophic Factor. Mol. Cell. Neurosci. 2006, 31, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.; Lewis, D.; Sibille, E. The role of BDNF in age-dependent changes of excitatory and inhibitory synaptic markers in the human prefrontal cortex. Neuropsycopharmacology 2016, 41, 3080–3091. [Google Scholar] [CrossRef] [PubMed]
- Miranda, M.; Morici, J.F.; Zanoni, M.B.; Bekinschtein, P. Brain-Derived Neurotrophic Factor: A Key Molecule for Memory in the Healthy and the Pathological Brain. Front. Cell. Neurosci. 2019, 13, 363. [Google Scholar] [CrossRef] [PubMed]







| Name | Full Name | Description | Measured in |
|---|---|---|---|
| α-SNCA | Alpha Synuclein | Component of Lewy bodies neuropathological marker of Parkinson’s Disease | pg/mg |
| Tau | Total Tau protein | Microtubule binding protein expressed in neurons. | ng/mg |
| pTau-181 | Tau protein phosphorylated at threonine 181 | Tau abnormally phosphorylated at threonine 181: a neuropathological marker of Alzheimer’s Disease | ng/mg |
| pTau:Tau | Ratio of pTau-181 to Tau | ||
| Aβ40 | Beta-amyloid 1–40 | 40-aminoacid beta-amyloid protein, a neuropathological marker of Alzheimer’s Disease. | pg/mg |
| Aβ42 | Beta-amyloid 1–42 | 42-aminoacid beta-amyloid protein, a neuropathological marker of Alzheimer’s Disease | pg/mg |
| Aβ42:Aβ40 | Ratio of Aβ42 to Aβ40 | ||
| IFNγ | interferon-gamma | Pro-inflammatory cytokine | pg/mg |
| TNFα | tumor necrosis factor alpha | Pro-inflammatory cytokine | pg/mg |
| RANTES | Regulated on activation, normal T cells expressed and secreted | Chemokine which recruits leukocytes to inflammatory sites | pg/mg |
| MIP-1α | Macrophage inflammatory protein-1 alpha | Chemokine with chemotactic and pro-inflammatory effects | pg/mg |
| MCP-1 | Monocyte chemotactic protein 1 | Chemokine which recruits monocytes to inflammatory sites | pg/mg |
| IL-7 | Interleukin-7 | Growth factor which stimulates lymphocyte differentiation | pg/mg |
| IL-6 | Interleukin-6 | Pro-inflammatory cytokine | pg/mg |
| IL-1β | Interleukin-1β | Pro-inflammatory cytokine | pg/mg |
| IL10 | Interleukin-10 | Pro-inflammatory cytokine | pg/mg |
| VEGF | Vascular endothelial growth factor | Growth factor which stimulates angiogenesis | pg/mg |
| BDNF | Brain-derived neurotrophic factor | Neurotrophic factor which promotes neuronal growth and differentiation | pg/mg ¹ |
| Donor ID * | DSM IV | Age (Years) | Sex | Education Years | RIN | Braak | CERAD | NIA Reagan |
|---|---|---|---|---|---|---|---|---|
| H14.09.094 | No Dementia | 78 | M | 18 | 8.4 | 2 | 1 | 1 |
| H14.09.074 | No Dementia | 82 | F | 16 | 6.7 | 3 | 1 | 1 |
| H14.09.096 | No Dementia | 84 | M | 12 | 5.9 | 3 | 2 | 2 |
| H14.09.060 | No Dementia | 86 | F | 12 | 5.6 | 4 | 2 | 2 |
| H15.09.106 | No Dementia | 86 | M | 14 | 7.1 | 2 | 0 | 1 |
| H14.09.078 | No Dementia | 87 | M | 16 | 6.5 | 1 | 0 | 1 |
| H14.09.058 | No Dementia | 88 | M | 14 | 6.8 | 1 | 1 | 1 |
| H14.09.062 | No Dementia | 89 | F | 16 | 7.9 | 3 | 1 | 1 |
| H14.09.072 | No Dementia | 89 | F | 15 | 5.1 | 4 | 1 | 1 |
| H14.09.090 | No Dementia | 89 | F | 12 | 7 | 5 | 1 | 2 |
| H14.09.006 | No Dementia | 90–94 | F | 14 | 6.7 | 3 | 0 | 1 |
| H14.09.030 | No Dementia | 90–94 | F | 15 | 6 | 3 | 3 | 2 |
| H14.09.052 | No Dementia | 90–94 | F | 14 | 7.4 | 2 | 1 | 1 |
| H15.09.108 | No Dementia | 90–94 | F | 12 | 5.5 | 4 | 3 | 2 |
| H14.09.004 | No Dementia | 90–94 | M | 14 | 7.7 | 3 | 1 | 1 |
| H14.09.102 | No Dementia | 95–99 | F | 16 | 7.2 | 1 | 0 | 1 |
| H14.09.046 | No Dementia | 95–99 | M | 13 | 6.2 | 1 | 2 | 2 |
| H14.09.050 | No Dementia | 95–99 | M | 8 | 6.7 | 3 | 3 | 2 |
| H14.09.070 | No Dementia | 95–99 | M | 16 | 7 | 2 | 0 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro, C.C.M.; Silva, S.P.; Rabelo, L.N.; Queiroz, J.P.G.; Campos, L.D.; Silva, L.C.; Fiuza, F.P. Age, Education Years, and Biochemical Factors Are Associated with Selective Neuronal Changes in the Elderly Hippocampus. Cells 2022, 11, 4033. https://doi.org/10.3390/cells11244033
Castro CCM, Silva SP, Rabelo LN, Queiroz JPG, Campos LD, Silva LC, Fiuza FP. Age, Education Years, and Biochemical Factors Are Associated with Selective Neuronal Changes in the Elderly Hippocampus. Cells. 2022; 11(24):4033. https://doi.org/10.3390/cells11244033
Chicago/Turabian StyleCastro, Carla Cristina Miranda, Sayonara Pereira Silva, Lívia Nascimento Rabelo, José Pablo Gonçalves Queiroz, Laura Damasceno Campos, Larissa Camila Silva, and Felipe Porto Fiuza. 2022. "Age, Education Years, and Biochemical Factors Are Associated with Selective Neuronal Changes in the Elderly Hippocampus" Cells 11, no. 24: 4033. https://doi.org/10.3390/cells11244033
APA StyleCastro, C. C. M., Silva, S. P., Rabelo, L. N., Queiroz, J. P. G., Campos, L. D., Silva, L. C., & Fiuza, F. P. (2022). Age, Education Years, and Biochemical Factors Are Associated with Selective Neuronal Changes in the Elderly Hippocampus. Cells, 11(24), 4033. https://doi.org/10.3390/cells11244033