
The Ancient and Evolved Mouse Sperm-Associated Antigen 6 Genes Have Different Biologic Functions In Vivo


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Constructs for the Studies
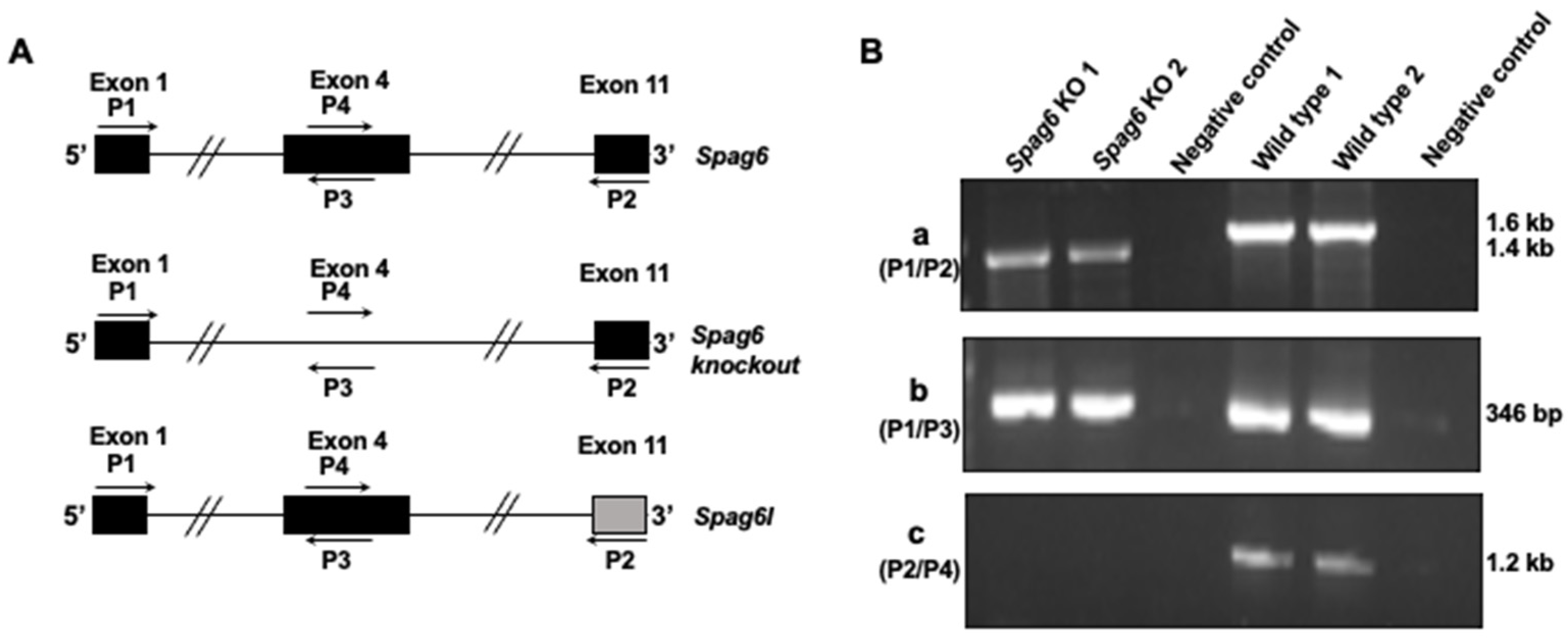
2.3. Animals and Genotyping
2.4. Histological Examination of Testicular and Epididymal Tissues
2.5. RT-PCR
2.6. Western Blotting
2.7. Cell Culture and Transient Transfection
2.8. Immunofluorescence Staining of Cultured Mammalian Cells
2.9. Male Fertility Test
2.10. Sperm Parameters
2.11. Direct Yeast Two-Hybrid Assay
2.12. Structural Analysis and Comparison
3. Results
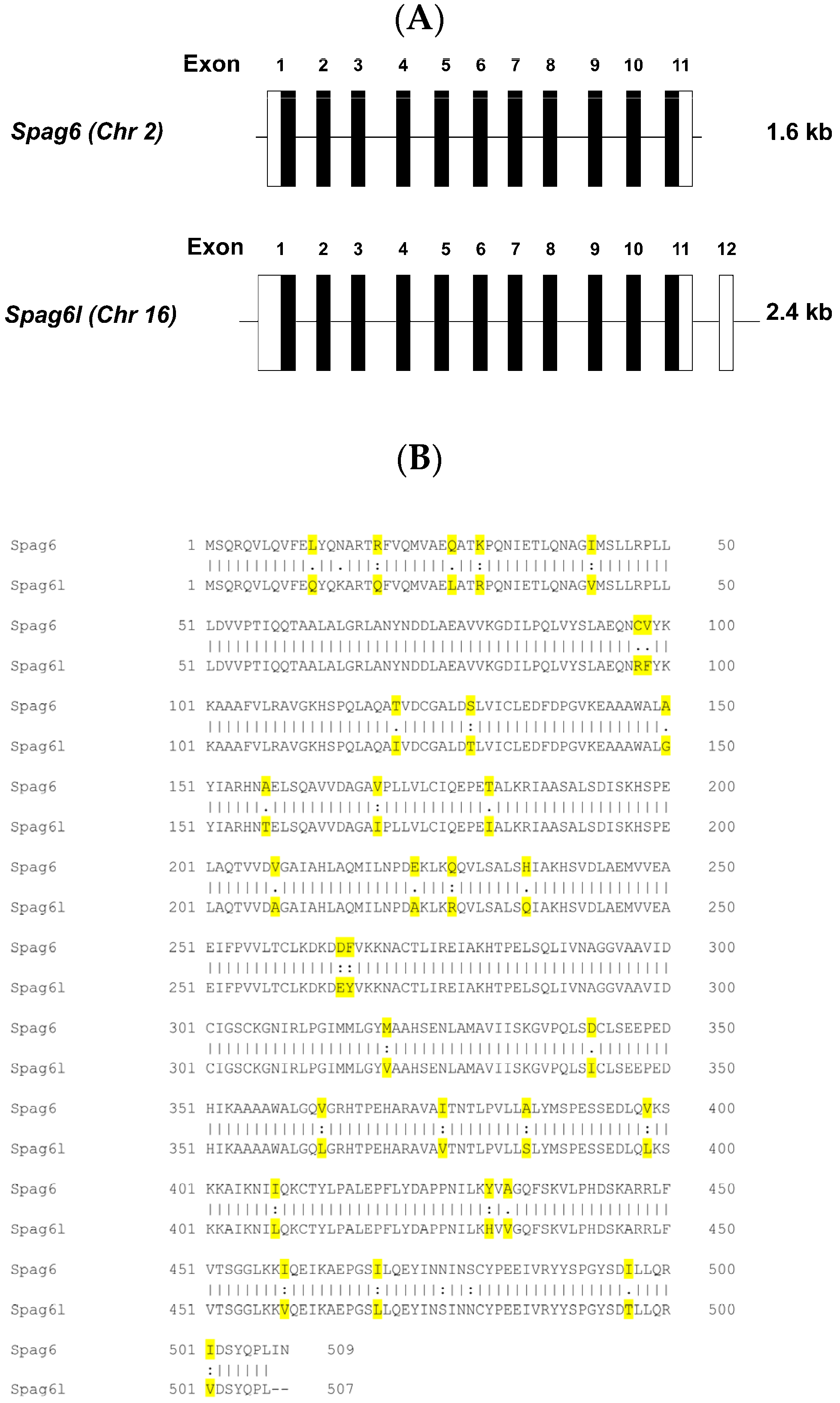
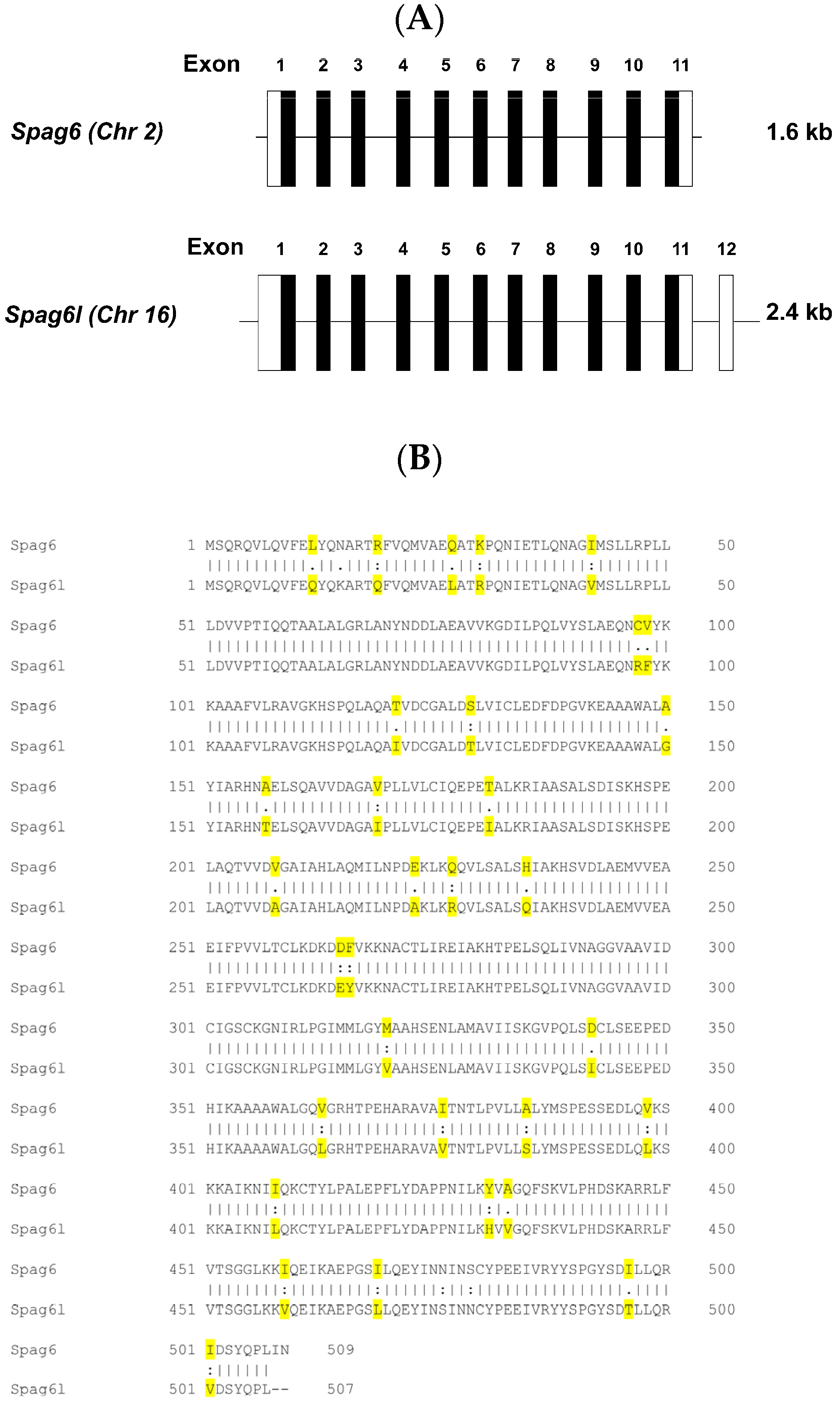
3.1. Spag6 Shares High Sequence Homology with Spag6l
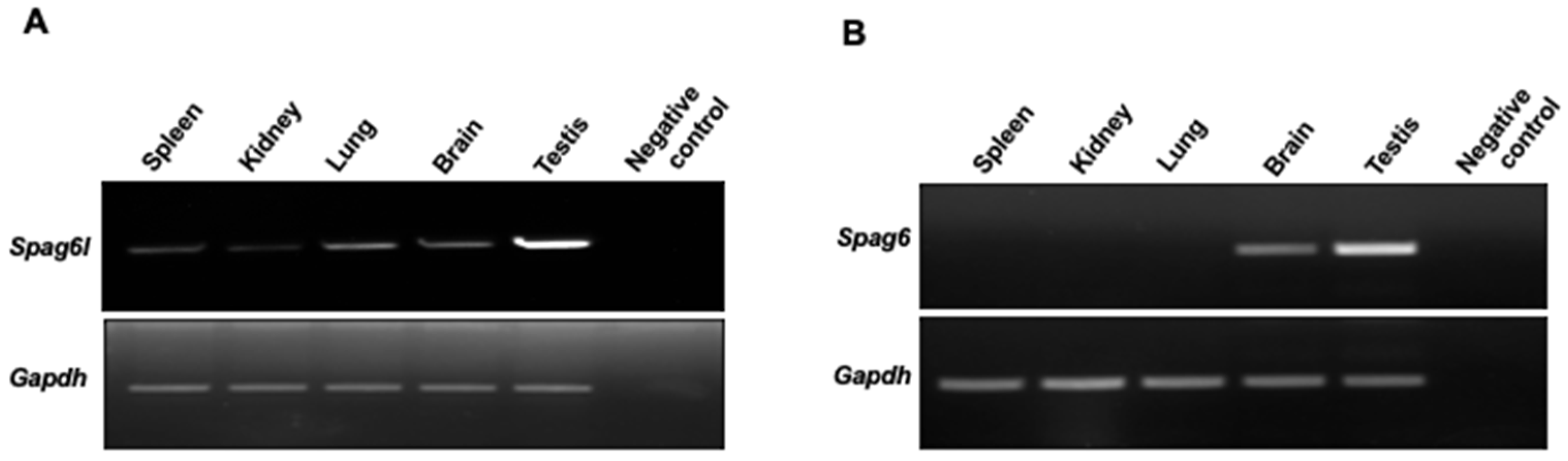
3.2. Spag6 and Spag6l Have Different mRNA Distributions In Vivo
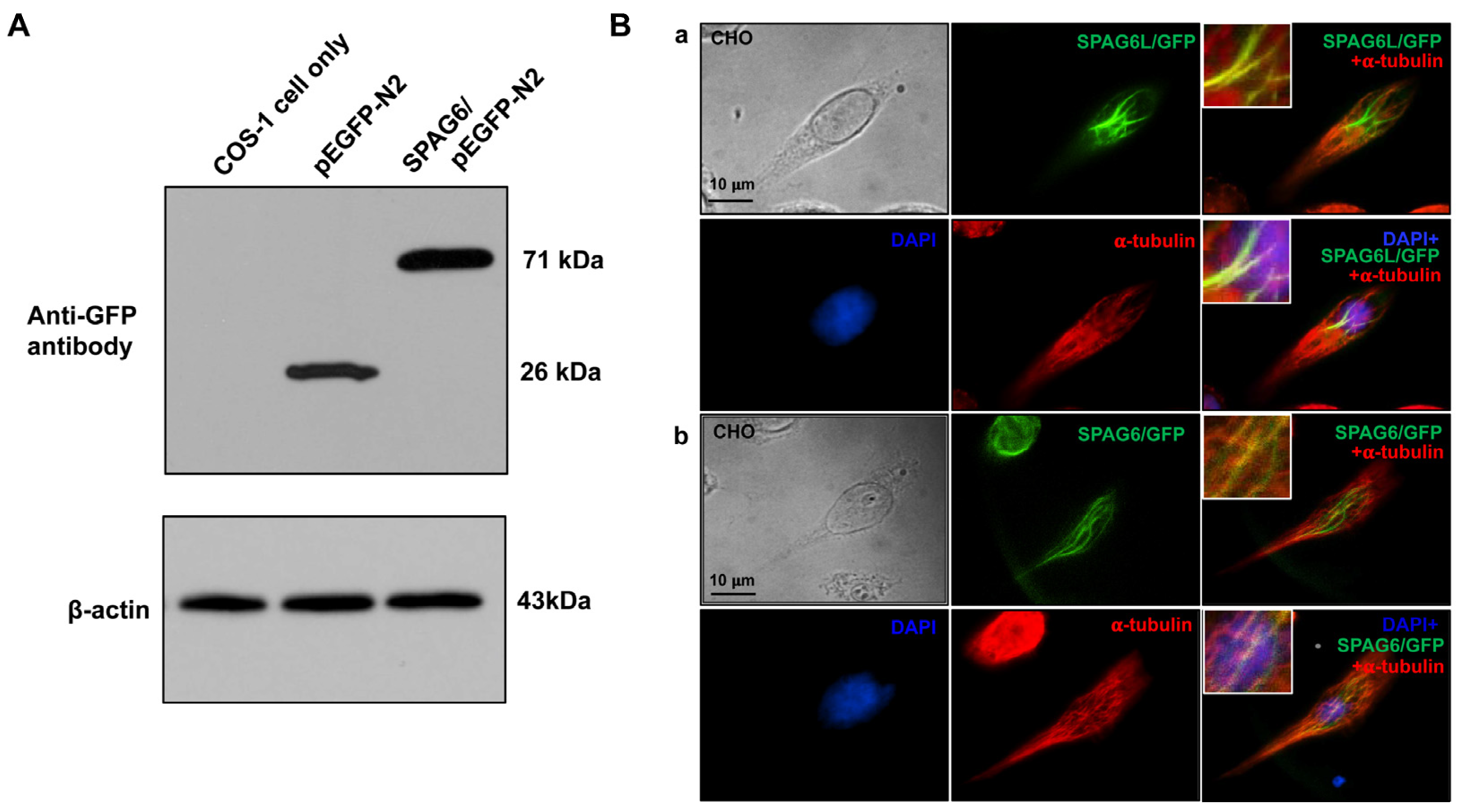
3.3. SPAG6 Protein Has a Similar Localization as SPAG6L in Transfected Mammalian Cells
3.4. Mice Deficient in Spag6 Gene Expression Are Grossly Normal and Fertile
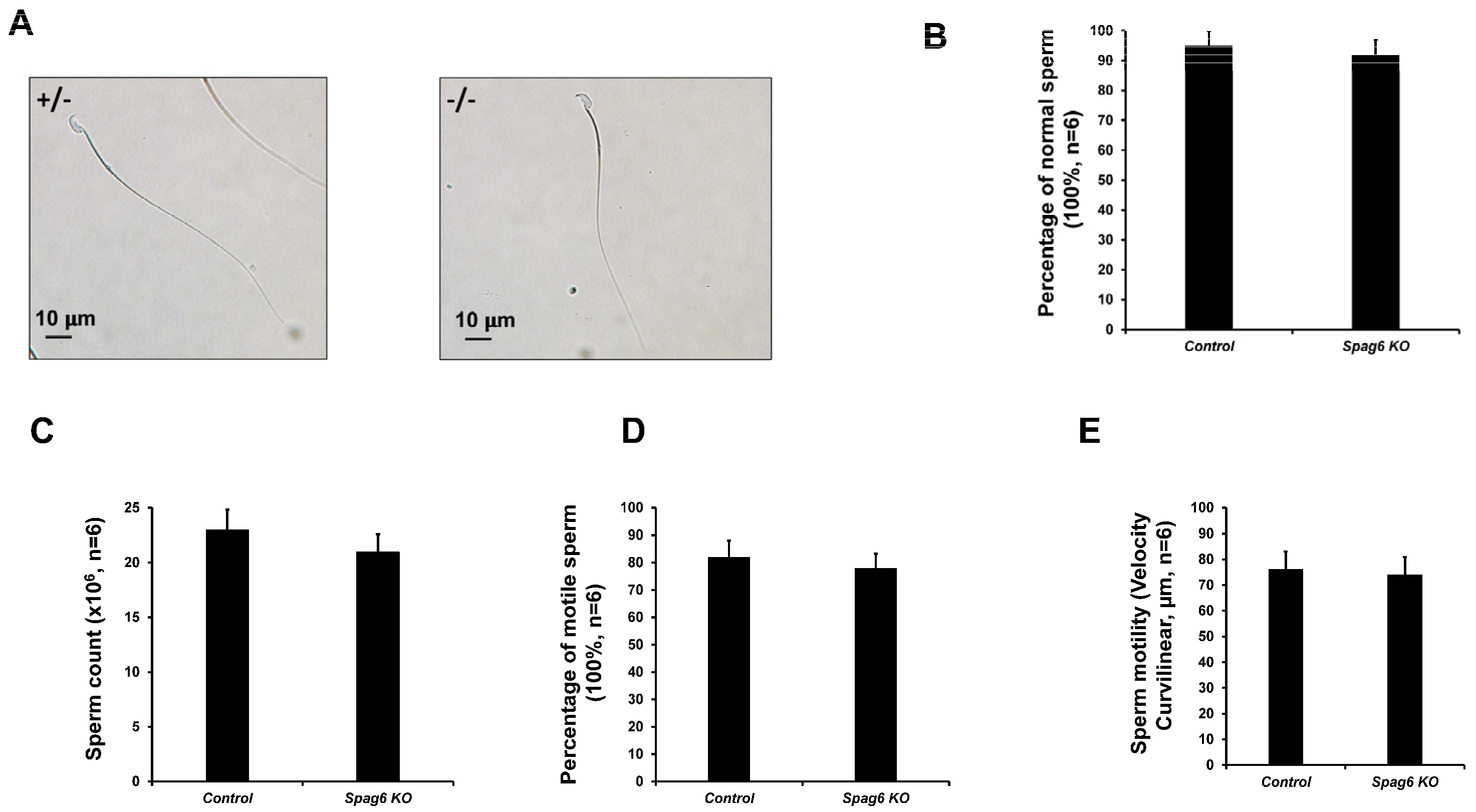
3.5. Spag6 KO Mice Have Normal Sperm Parameters
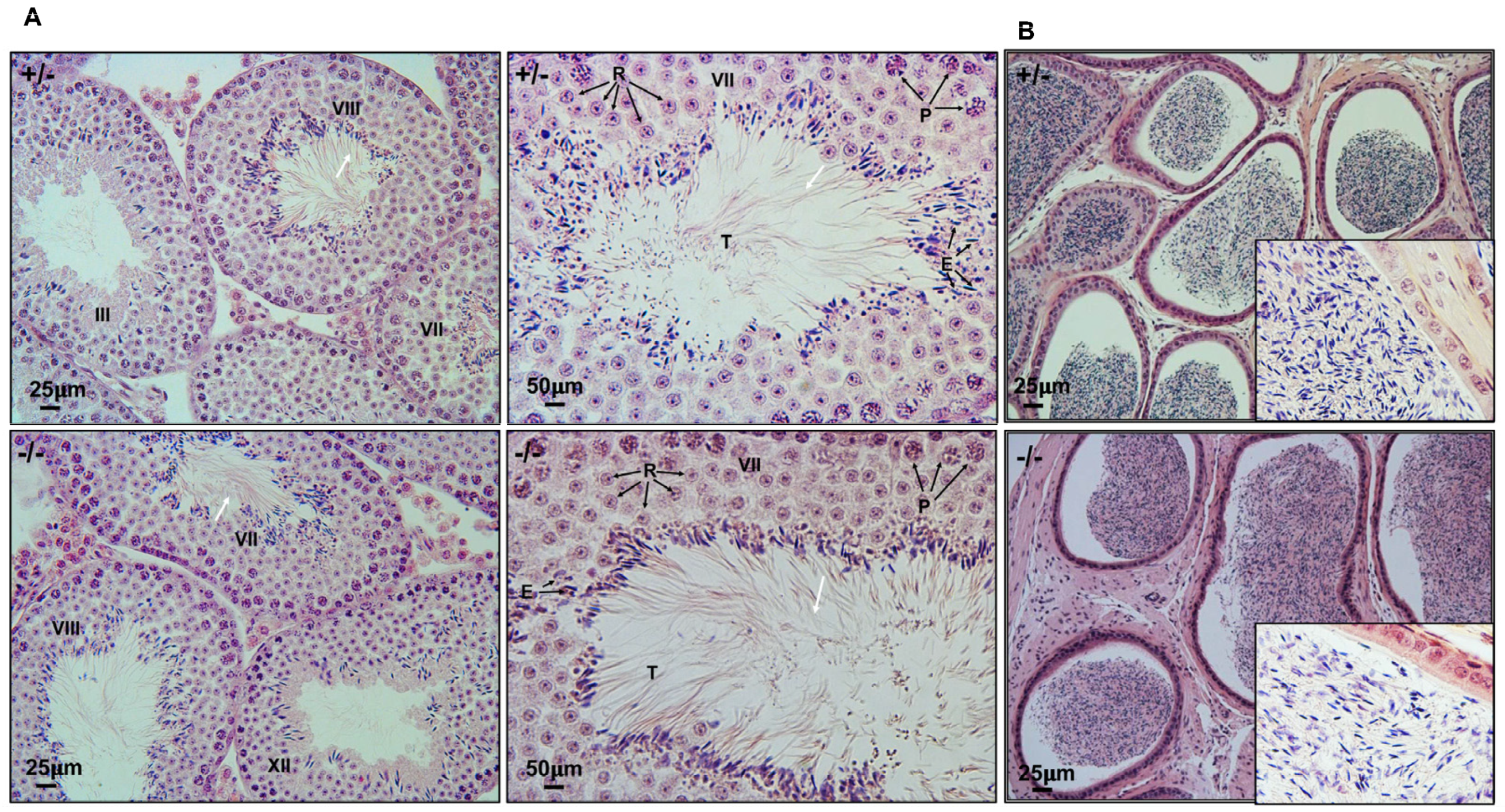
3.6. Spermatogenesis Is Not Affected in Spag6 KO Mice
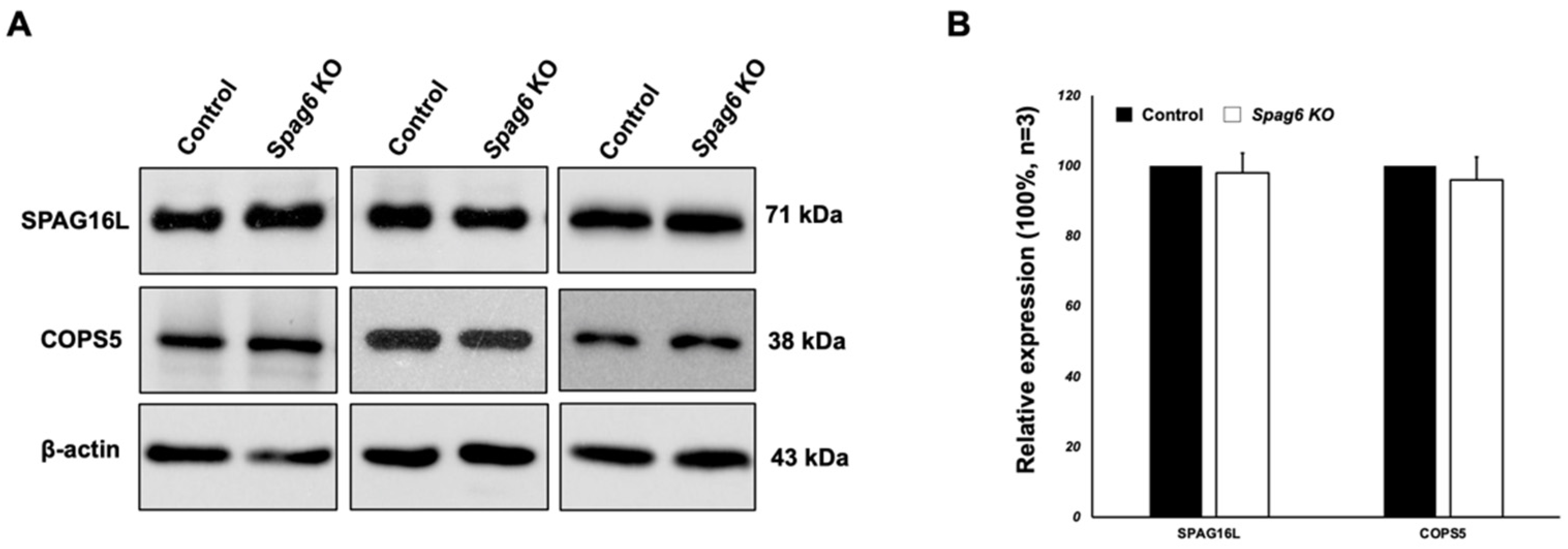
3.7. Testicular Expression of Selected SPAG6L-Binding Partners in the Spag6-Deficient Mice
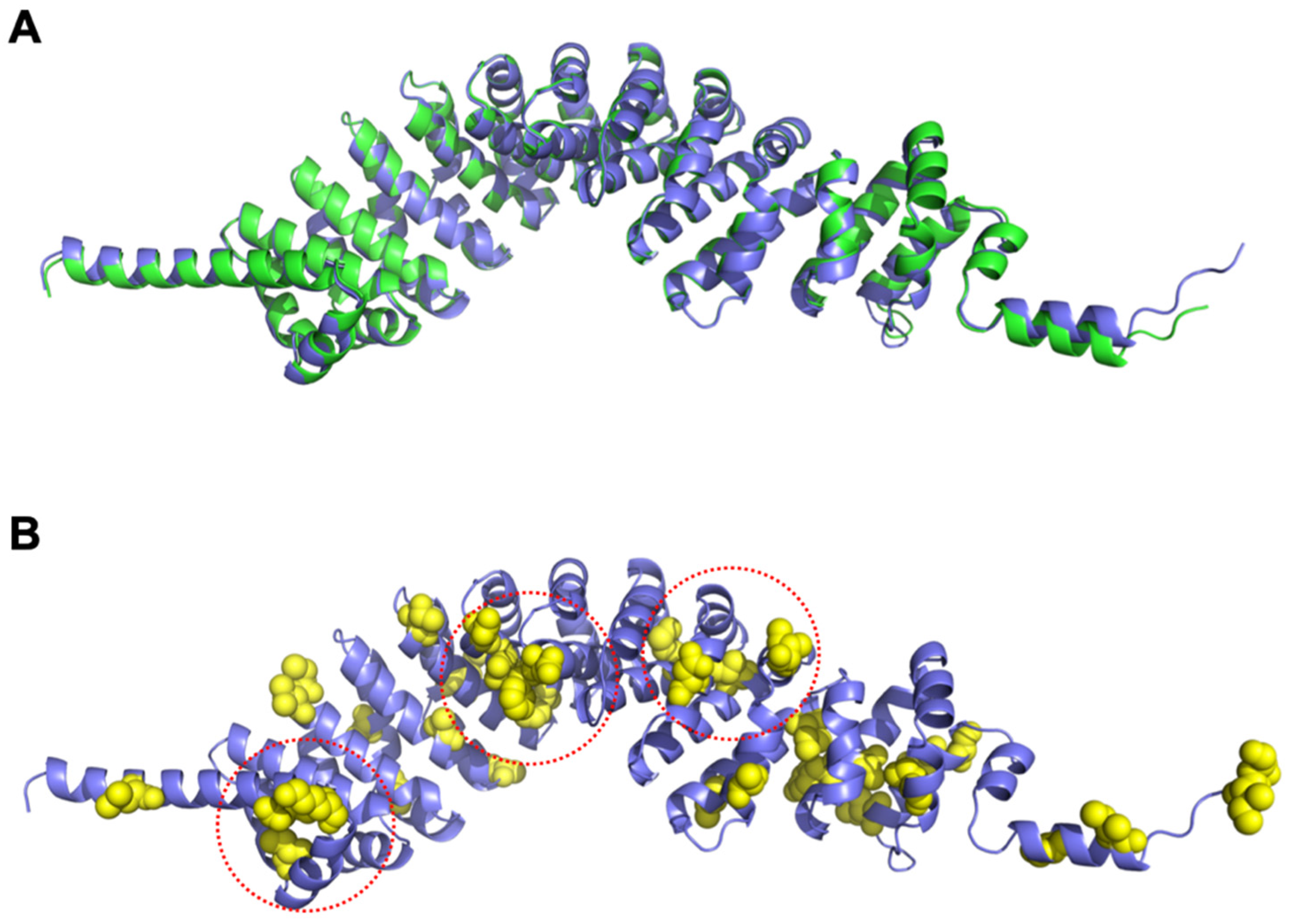
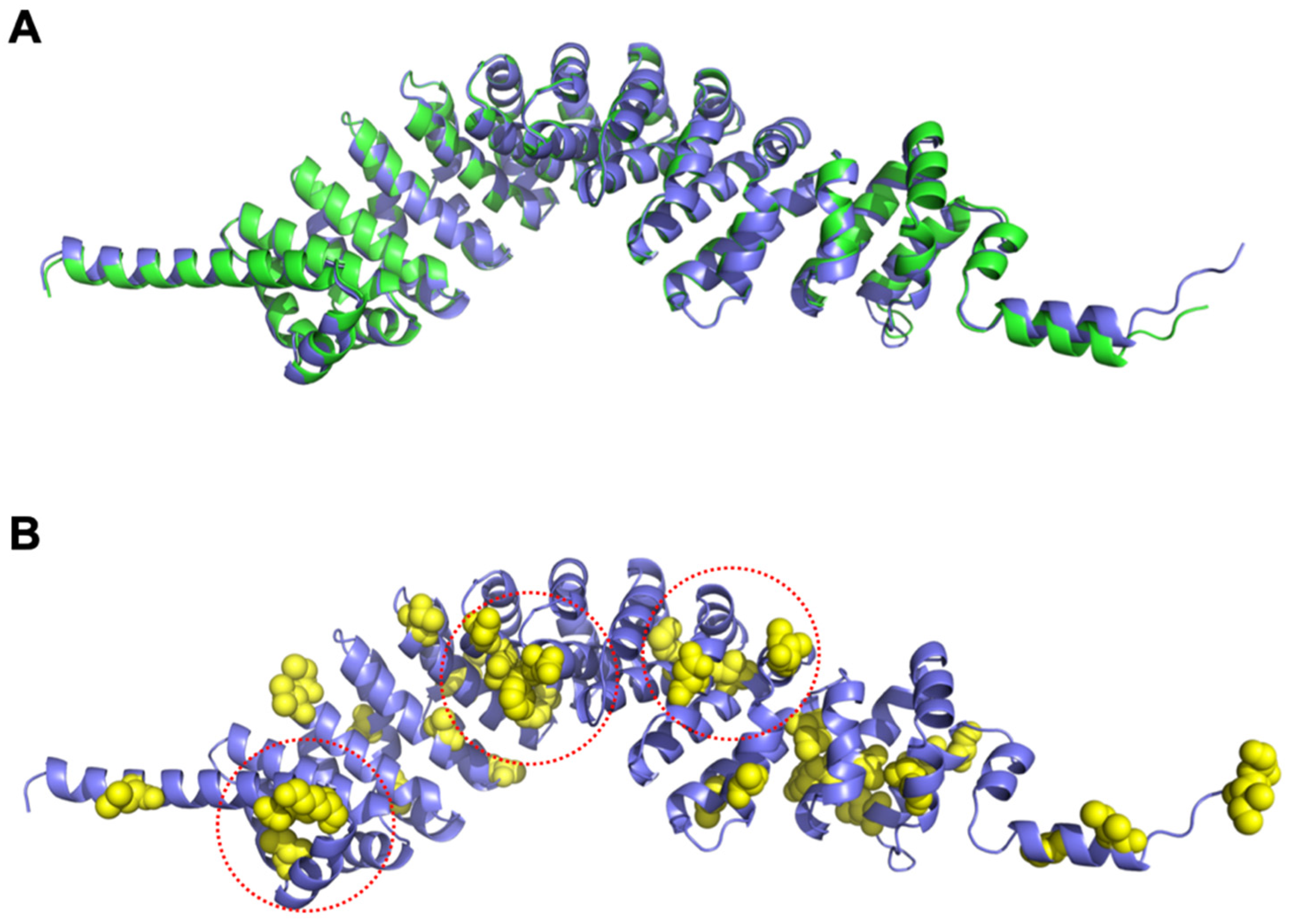
3.8. Structural Differences between SPAG6 and SPAG6L
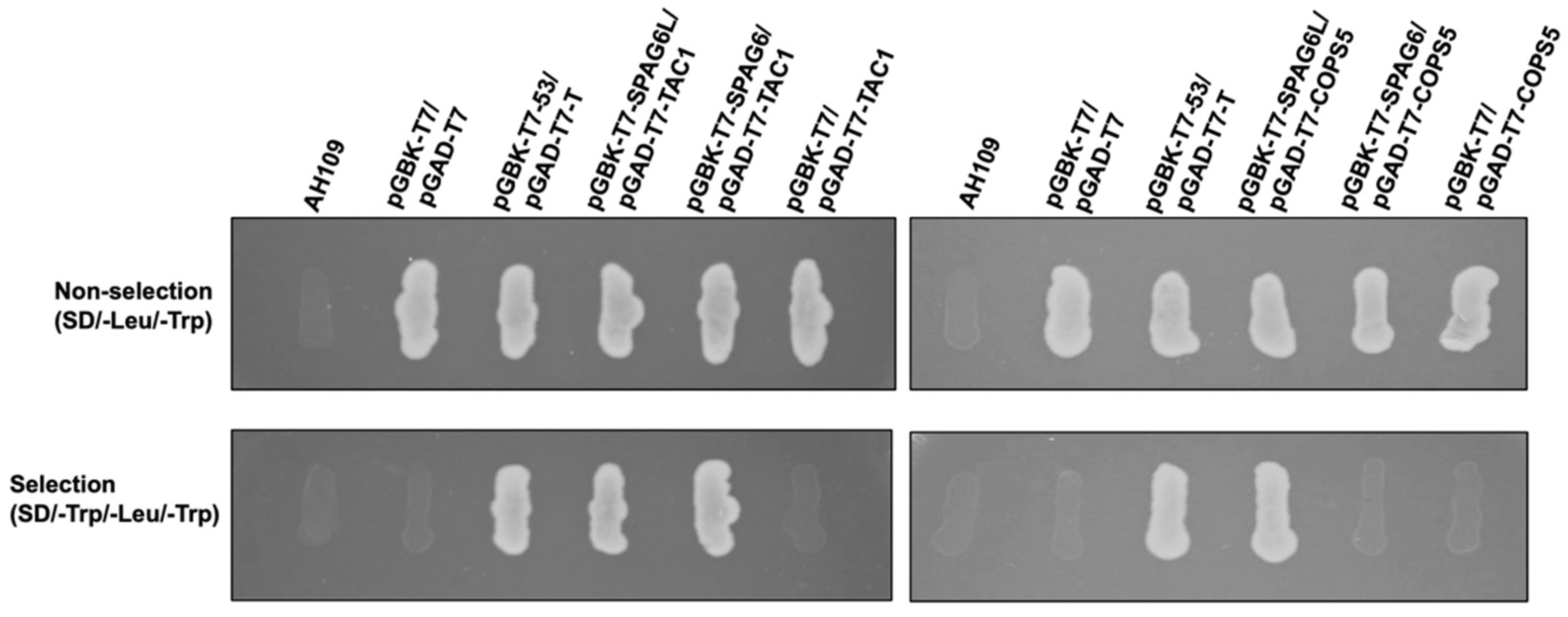
3.9. SPAG6 and SPAG6L Bind to Different Proteins
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, E.F.; Lefebvre, P.A. PF16 encodes a protein with armadillo repeats and localizes to a single microtubule of the central apparatus in Chlamydomonas flagella. J. Cell Biol. 1996, 132, 359–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neilson, L.I.; Schneider, P.A.; Van Deerlin, P.G.; Kiriakidou, M.; Driscoll, D.A.; Pellegrini, M.C.; Millinder, S.; Yamamoto, K.K.; French, C.K.; Strauss, J.F., 3rd. cDNA cloning and characterization of a human sperm antigen (SPAG6) with homology to the product of the Chlamydomonas PF16 locus. Genomics 1999, 60, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Sapiro, R.; Tarantino, L.M.; Velazquez, F.; Kiriakidou, M.; Hecht, N.B.; Bucan, M.; Strauss, J.F., 3rd. Sperm antigen 6 is the murine homologus of the Chlamydomonas reinhardtii central apparatus protein encoded by the PF16 locus. Biol. Reprod. 2000, 62, 511–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sapiro, R.; Kostetskii, I.; Olds-Clarke, P.; Gerton, G.L.; Radice, G.L.; Strauss, J.F., 3rd. Male infertility, impaired sperm motility, and hydrocephalus in mice deficient in sperm-associated antigen 6. Mol. Cell. Biol. 2002, 22, 6298–6305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teves, M.E.; Sears, P.R.; Li, W.; Zhang, Z.; Tang, W.; van Reesema, L.; Costanzo, R.M.; Davis, C.W.; Knowles, M.R.; Strauss, J.F., 3rd; et al. Sperm-associated antigen 6 (SPAG6) deficiency and defects in ciliogenesis and cilia function: Polarity, density, and beat. PLoS ONE 2014, 9, e107271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Xu, L.; Li, J.; Li, B.; Bai, X.; Strauss, J.F., 3rd; Zhang, Z.; Wang, H. Otitis media in sperm-associated antigen 6 (Spag6)-deficient mice. PLoS ONE 2014, 9, e112879. [Google Scholar] [CrossRef]
- Li, W.; Mukherjee, A.; Wu, J.; Zhang, L.; Teves, M.E.; Li, H.; Nabmiar, S.; Henderson, S.C.; Horwitz, A.R.; Strauss, J.F., III; et al. Sperm associated antigen 6 (SPAG6) regulates fibroblast cell growth, morphology, migration and ciliogenesis. Sci. Rep. 2015, 5, 16506. [Google Scholar] [CrossRef]
- Cooley, L.F.; El Shikh, M.E.; Li, W.; Keim, R.C.; Zhang, Z.; Strauss, J.F.; Zhang, Z.; Conrad, D.H. Impaired immunological synapse in sperm associated antigen 6 (SPAG6) deficient mice. Sci. Rep. 2016, 6, 25840. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xu, L.; Sun, G.; Wu, X.; Bai, X.; Li, J.; Strauss, J.F.; Zhang, Z.; Wang, H. Spag6 Mutant Mice Have Defects in Development and Function of Spiral Ganglion Neurons, Apoptosis, and Higher Sensitivity to Paclitaxel. Sci. Rep. 2017, 7, 8638. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, L.; Li, W.; Huang, Q.; Yuan, S.; Li, Y.; Liu, J.; Zhang, S.; Pin, G.; Song, S.; et al. The sperm-associated antigen 6 interactome and its role in spermatogenesis. Reproduction 2019, 158, 181–197. [Google Scholar] [CrossRef]
- Li, X.; Zhang, D.; Xu, L.; Han, Y.; Liu, W.; Li, W.; Fan, Z.; Costanzo, R.M.; Strauss, J.F., 3rd; Zhang, Z.; et al. Planar cell polarity defects and hearing loss in sperm-associated antigen 6 (Spag6)-deficient mice. Am. J. Physiol. Cell Physiol. 2021, 320, C132–C141. [Google Scholar] [CrossRef]
- Li, H.; Lv, J.; Zhou, Q.; Jin, L.; Kang, Z.; Huang, Y. Establishment of sperm associated antigen 6 gene knockout mouse model and its mechanism of deafness. Saudi J. Biol. Sci. 2020, 27, 1289–1295. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.; Lee, D.J.; Kim, M.H.; Bok, J. Identification of genes concordantly expressed with Atoh1 during inner ear development. Anat. Cell Biol. 2011, 44, 69–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Z.; Zhu, C.; Chang, G.; Wu, N.; Ding, H.; Wang, H. Differential expression analysis and identification of sex-related genes by gonad transcriptome sequencing in estradiol-treated and non-treated Ussuri catfish Pseudobagrus ussuriensis. Fish. Physiol. Biochem. 2021, 47, 565–581. [Google Scholar] [CrossRef] [PubMed]
- Jarrell, Z.R.; Ahammad, M.U.; Sweeney, K.; Wilson, J.L.; Benson, A.P. Characterization of sperm-associated antigen 6 expression in the reproductive tract of the domestic rooster (Gallus domesticus) and its impact on sperm mobility. Poult. Sci. 2020, 99, 6188–6195. [Google Scholar] [CrossRef] [PubMed]
- Gust, K.A.; Lotufo, G.R.; Thiyagarajah, A.; Barker, N.D.; Ji, Q.; Marshall, K.; Wilbanks, M.S.; Chappell, P. Molecular Evaluation of Impacted Reproductive Physiology in Fathead Minnow Testes Provides Mechanistic Insights into Insensitive Munitions Toxicology. Aquat. Toxicol. 2019, 213, 105204. [Google Scholar] [CrossRef] [PubMed]
- Chi, W.; Gao, Y.; Hu, Q.; Guo, W.; Li, D. Genome-wide analysis of brain and gonad transcripts reveals changes of key sex reversal-related genes expression and signaling pathways in three stages of Monopterus albus. PLoS ONE 2017, 12, e0173974. [Google Scholar] [CrossRef]
- Fischer, D.; Laiho, A.; Gyenesei, A.; Sironen, A. Identification of Reproduction-Related Gene Polymorphisms Using Whole Transcriptome Sequencing in the Large White Pig Population. G3 Genes Genomes Genet. 2015, 5, 1351–1360. [Google Scholar] [CrossRef] [Green Version]
- Hamada, T.; Teraoka, M.; Imaki, J.; Ui-Tei, K.; Ladher, R.K.; Asahara, T. Gene expression of Spag6 in chick central nervous system. Anat. Histol. Embryol. 2010, 39, 227–232. [Google Scholar] [CrossRef]
- Wu, H.; Wang, J.; Cheng, H.; Gao, Y.; Liu, W.; Zhang, Z.; Jiang, H.; Li, W.; Zhu, F.; Lv, M.; et al. Patients with severe asthenoteratospermia carrying SPAG6 or RSPH3 mutations have a positive pregnancy outcome following intracytoplasmic sperm injection. Assist. Reprod. Genet. 2020, 37, 829–840. [Google Scholar] [CrossRef]
- Steinbach, D.; Schramm, A.; Eggert, A.; Onda, M.; Dawczynski, K.; Rump, A.; Pastan, I.; Wittig, S.; Pfaffendorf, N.; Voigt, A.; et al. Identification of a set of seven genes for the monitoring of minimal residual disease in pediatric acute myeloid leukemia. Clin. Cancer Res. 2006, 12, 2434–2441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silina, K.; Zayakin, P.; Kalniņa, Z.; Ivanova, L.; Meistere, I.; Endzeliņš, E.; Abols, A.; Stengrēvics, A.; Leja, M.; Ducena, K.; et al. Sperm-associated antigens as targets for cancer immunotherapy: Expression pattern and humoral immune response in cancer patients. J. Immunother. 2011, 34, 28–44. [Google Scholar] [CrossRef]
- Mulaw, M.A.; Krause, A.; Deshpande, A.J.; Krause, L.F.; Rouhi, A.; La Starza, R.; Borkhardt, A.; Buske, C.; Mecucci, C.; Ludwig, W.D.; et al. CALM/AF10-positive leukemias show upregulation of genes involved in chromatin assembly and DNA repair processes and of genes adjacent to the breakpoint at 10p12. Leukemia 2012, 26, 1012–1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinbach, D.; Bader, P.; Willasch, A.; Bartholomae, S.; Debatin, K.M.; Zimmermann, M.; Creutzig, U.; Reinhardt, D.; Gruhn, B. Prospective validation of a new method of monitoring minimal residual disease in childhood acute myelogenous leukemia. Clin. Cancer Res. 2015, 21, 1353–1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitchen, M.O.; Bryan, R.T.; Haworth, K.E.; Emes, R.D.; Luscombe, C.; Gommersall, L.; Cheng, K.K.; Zeegers, M.P.; James, N.D.; Devall, A.J.; et al. Methylation of HOXA9 and ISL1 Predicts Patient Outcome in High-Grade Non-Invasive Bladder Cancer. PLoS ONE 2015, 10, e0137003. [Google Scholar]
- Altenberger, C.; Heller, G.; Ziegler, B.; Tomasich, E.; Marhold, M.; Topakian, T.; Müllauer, L.; Heffeter, P.; Lang, G.; End-Pfützenreuter, A.; et al. SPAG6 and L1TD1 are transcriptionally regulated by DNA methylation in non-small cell lung cancers. Mol. Cancer 2017, 16, 1. [Google Scholar] [CrossRef] [Green Version]
- Coan, M.; Rampioni Vinciguerra, G.L.; Cesaratto, L.; Gardenal, E.; Bianchet, R.; Dassi, E.; Vecchione, A.; Baldassarre, G.; Spizzo, R.; Nicoloso, M.S. Exploring the Role of Fallopian Ciliated Cells in the Pathogenesis of High-Grade Serous Ovarian Cancer. Int. J. Mol. Sci. 2018, 19, 2512. [Google Scholar] [CrossRef] [Green Version]
- Jiang, M.; Chen, Y.; Deng, L.; Luo, X.; Wang, L.; Liu, L. Upregulation of SPAG6 in Myelodysplastic Syndrome: Knockdown Inhibits Cell Proliferation via AKT/FOXO Signaling Pathway. DNA Cell Biol. 2019, 38, 476–484. [Google Scholar] [CrossRef]
- Qiu, H.; Gołas, A.; Grzmil, P.; Wojnowski, L. Lineage-specific duplications of Muroidea Faim and Spag6 genes and atypical accelerated evolution of the parental Spag6 gene. J. Mol. Evol. 2013, 77, 119–129. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Mirdita, M.; Ovchinnikov, S.; Steinegger, M. ColabFold—Making Protein Folding Accessible to All. bioRxiv 2021, preprint. [Google Scholar]
- McLachlan, A.D. Rapid Comparison of Protein Structures. Acta Cryst. 1982, A38, 871–873. [Google Scholar] [CrossRef]
- Leppek, K.; Das, R.; Barna, M. Functional 5′ UTR mRNA structures in eukaryotic translation regulation and how to find them. Nat. Rev. Mol. Cell Biol. 2018, 19, 158–174. [Google Scholar] [CrossRef] [PubMed]
- Tewari, R.; Bailes, E.; Bunting, K.A.; Coates, J.C. Armadillo-repeat protein functions: Questions for little creatures. Trends Cell Biol. 2010, 20, 470–481. [Google Scholar] [CrossRef] [PubMed]
- Falkenberg, L.G.; Beckman, S.A.; Ravisankar, P.; Dohn, T.E.; Waxman, J.S. Ccdc103 promotes myeloid cell proliferation and migration independent of motile cilia. DMM Dis. Model. Mech. 2021, 14, dmm048439. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spag6 Genotype | Male Fertility a | Litter Size | Female Fertility a | Litter Size |
|---|---|---|---|---|
| +/− | 6/6 | 7.9 ± 0.5 | 6/6 | 8.1 ± 0.6 |
| −/− | 6/6 | 7.6 ± 0.7 | 6/6 | 7.4 ± 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yap, Y.T.; Li, W.; Zhou, Q.; Haj-Diab, S.; Chowdhury, D.D.; Vaishnav, A.; Harding, P.; Williams, D.C., Jr.; Edwards, B.F.; Strauss, J.F., III; et al. The Ancient and Evolved Mouse Sperm-Associated Antigen 6 Genes Have Different Biologic Functions In Vivo. Cells 2022, 11, 336. https://doi.org/10.3390/cells11030336
Yap YT, Li W, Zhou Q, Haj-Diab S, Chowdhury DD, Vaishnav A, Harding P, Williams DC Jr., Edwards BF, Strauss JF III, et al. The Ancient and Evolved Mouse Sperm-Associated Antigen 6 Genes Have Different Biologic Functions In Vivo. Cells. 2022; 11(3):336. https://doi.org/10.3390/cells11030336
Chicago/Turabian StyleYap, Yi Tian, Wei Li, Qi Zhou, Sarah Haj-Diab, Dipanwita Dutta Chowdhury, Asmita Vaishnav, Pamela Harding, David C. Williams, Jr., Brian F. Edwards, Jerome F. Strauss, III, and et al. 2022. "The Ancient and Evolved Mouse Sperm-Associated Antigen 6 Genes Have Different Biologic Functions In Vivo" Cells 11, no. 3: 336. https://doi.org/10.3390/cells11030336
APA StyleYap, Y. T., Li, W., Zhou, Q., Haj-Diab, S., Chowdhury, D. D., Vaishnav, A., Harding, P., Williams, D. C., Jr., Edwards, B. F., Strauss, J. F., III, & Zhang, Z. (2022). The Ancient and Evolved Mouse Sperm-Associated Antigen 6 Genes Have Different Biologic Functions In Vivo. Cells, 11(3), 336. https://doi.org/10.3390/cells11030336