
Topical Spray of dsRNA Induces Mortality and Inhibits Chilli Leaf Curl Virus Transmission by Bemisia tabaci Asia II 1


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Whitefly Population
2.2. Virus Culture
2.3. Designing and Synthesis of dsRNA

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5’-3’) | Amplicon Size | Annealing Temperature in PCR/Real-Time PCR | Amplified Region | Purpose | Reference |
|---|---|---|---|---|---|---|
| C1-J-2195 | TTGATTTTTTGGTCATCCAGAAGT | 860 bp | 53 °C | B. tabaci mtCOI | Detection of B. tabaci cryptic species | [46] |
| L2-N-3014 | TCCAATGCACTAATCTGCCATATTA | |||||
| Begomo F | ACGCGTGCCGTGCTGCTGCCCCCATTGTCC | 2.7 kb | 57 °C | Begomovirus DNA-A | Detection of begomovirus | [44] |
| Begomo R | ACGCGTATGGGCTGYCGAAGTTSAGAC | |||||
| AG137F | TCAAAGAACATTTTTGTGCTACT | 128 bp | 56 °C | B. tabaci hsp70 dsRNA | dsRNA synthesis | This study |
| AG138R | GACCATTGTCTAGGTCTTCATTT | |||||
| AG283F | CTGGTGTTTTGACAATCGAC | 150 bp | 56 °C | B. tabaci fas2 dsRNA | dsRNA synthesis and RT-qPCR | This study |
| AG284R | TGATTATGCCTTCTTCCGTC | |||||
| AG177F | ACATGGAAAAGATCTGGCAT | 121 bp | 56 °C | B. tabaci β-actin | RT-qPCR | This study |
| AG178R | TGAGTCATCTTTTCACGGTT | |||||
| AG204F | GTCAATGATTGCAGTAAGCC | 105 bp | 56 °C | B. tabaci hsp70 | RT-qPCR | This study |
| AG205R | TTCCCTCATTTTCGTAAGCA | |||||
| AG149F | TGAACAGGCCCATGAACAG | 290 bp | 53 °C | ChiLCV coat protein | qPCR and ChiLCV detection | [47] |
| AG150R | ACGGACAAGGAAAAACATCAC |
2.4. Bioassay of dsRNA through Artificial Feeding under Controlled Conditions
2.5. Estimating Relative Expression of hsp70 and fas2 mRNA
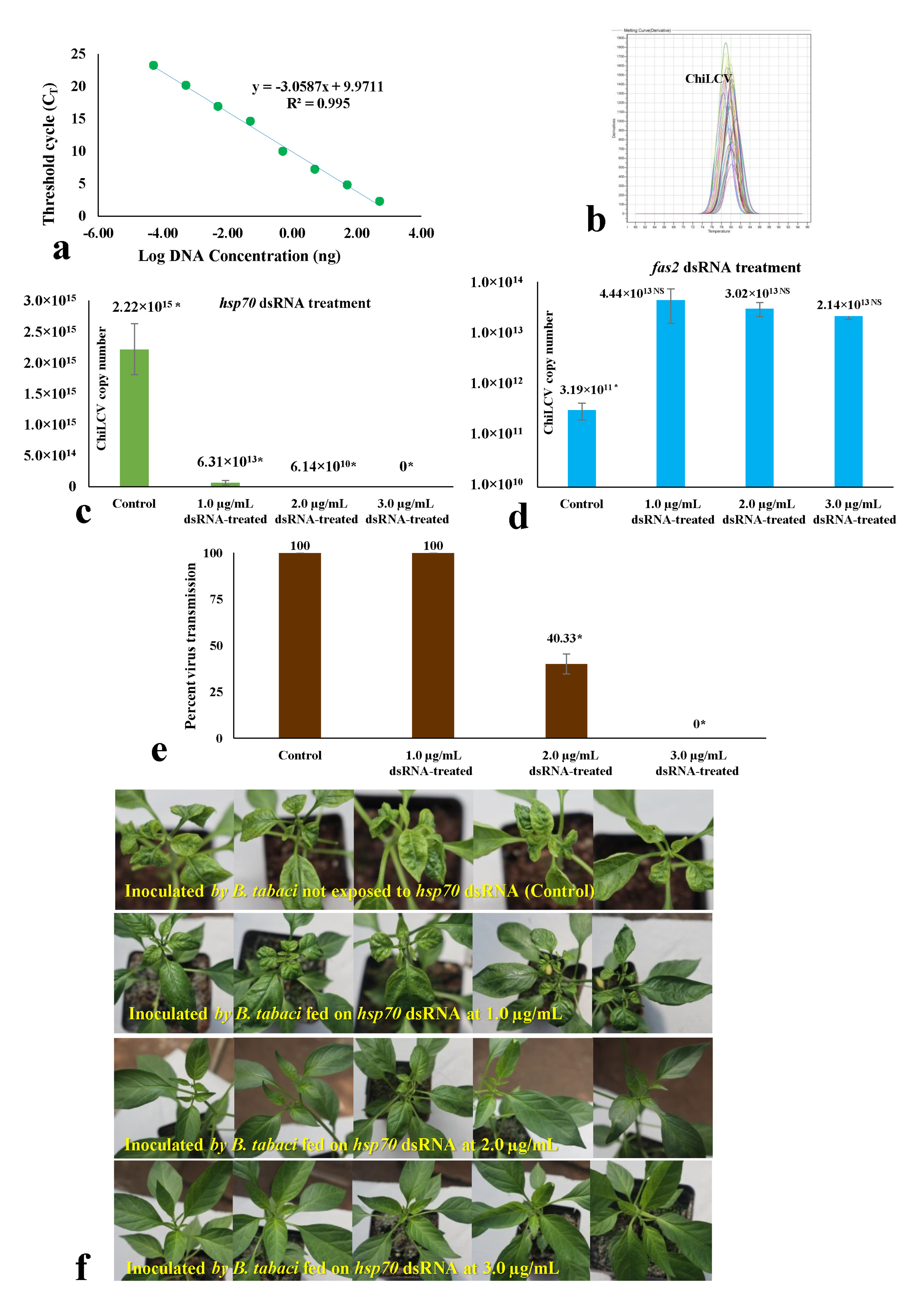
2.6. Quantification of Virus Copies in B. tabaci and Transmission of ChiLCV
2.7. Topical Spray of hsp70 dsRNA under Contained Semi-Field Conditions
2.8. Stability of hsp70 dsRNA in Leaf Tissue
2.9. Persistent Efficacy of hsp70 dsRNA to Eradicate B. tabaci
3. Results
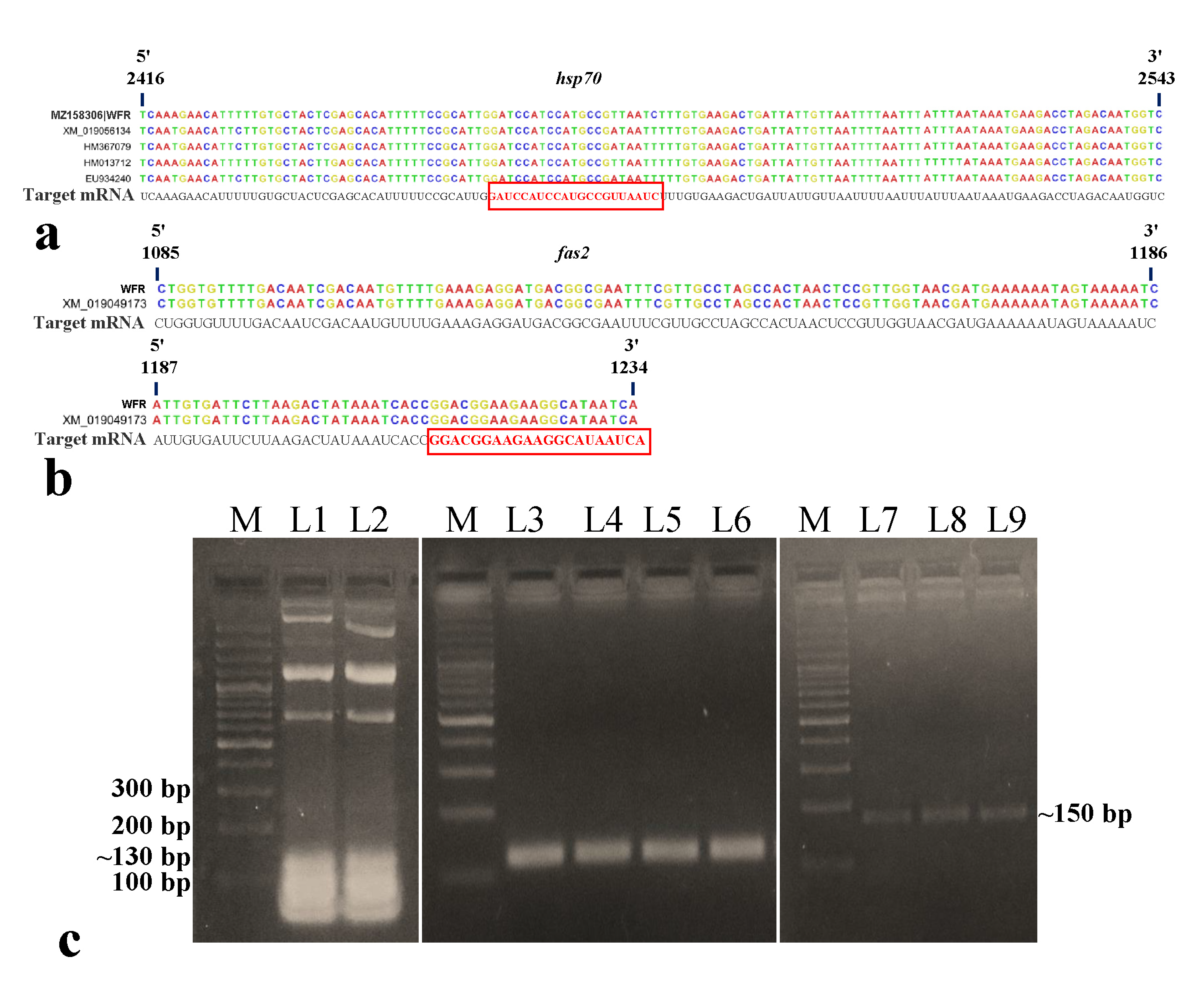
3.1. Characterization of B. tabaci and Begomovirus
3.2. Synthesis of dsRNA Targeting B. tabaci hsp70 and fas2
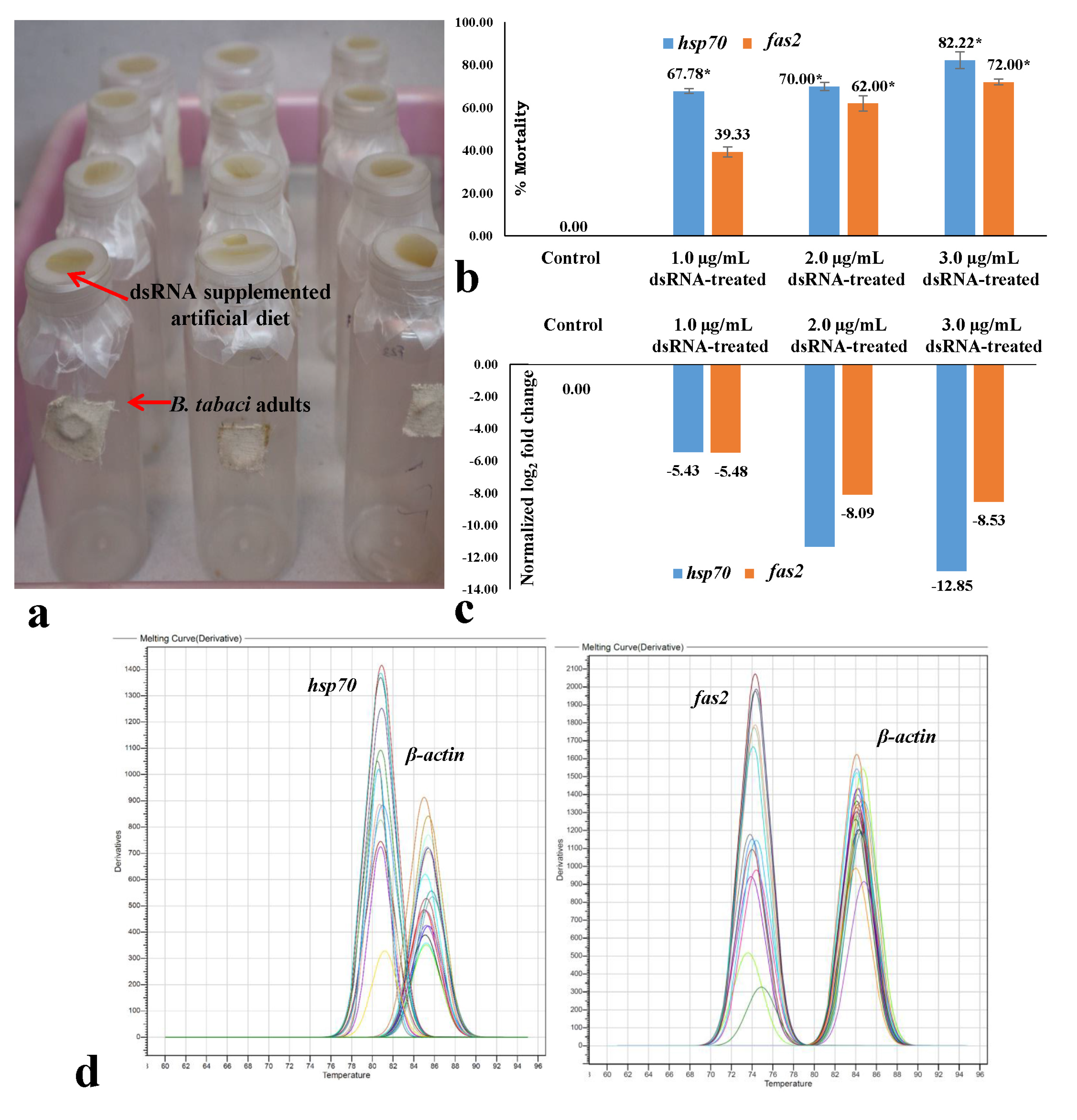
3.3. Effect of dsRNA on Mortality and mRNA Expression of B. tabaci under Controlled Conditions
3.4. Effect of dsRNA on Virus Acquisition and Transmission by B. tabaci under Controlled Conditions
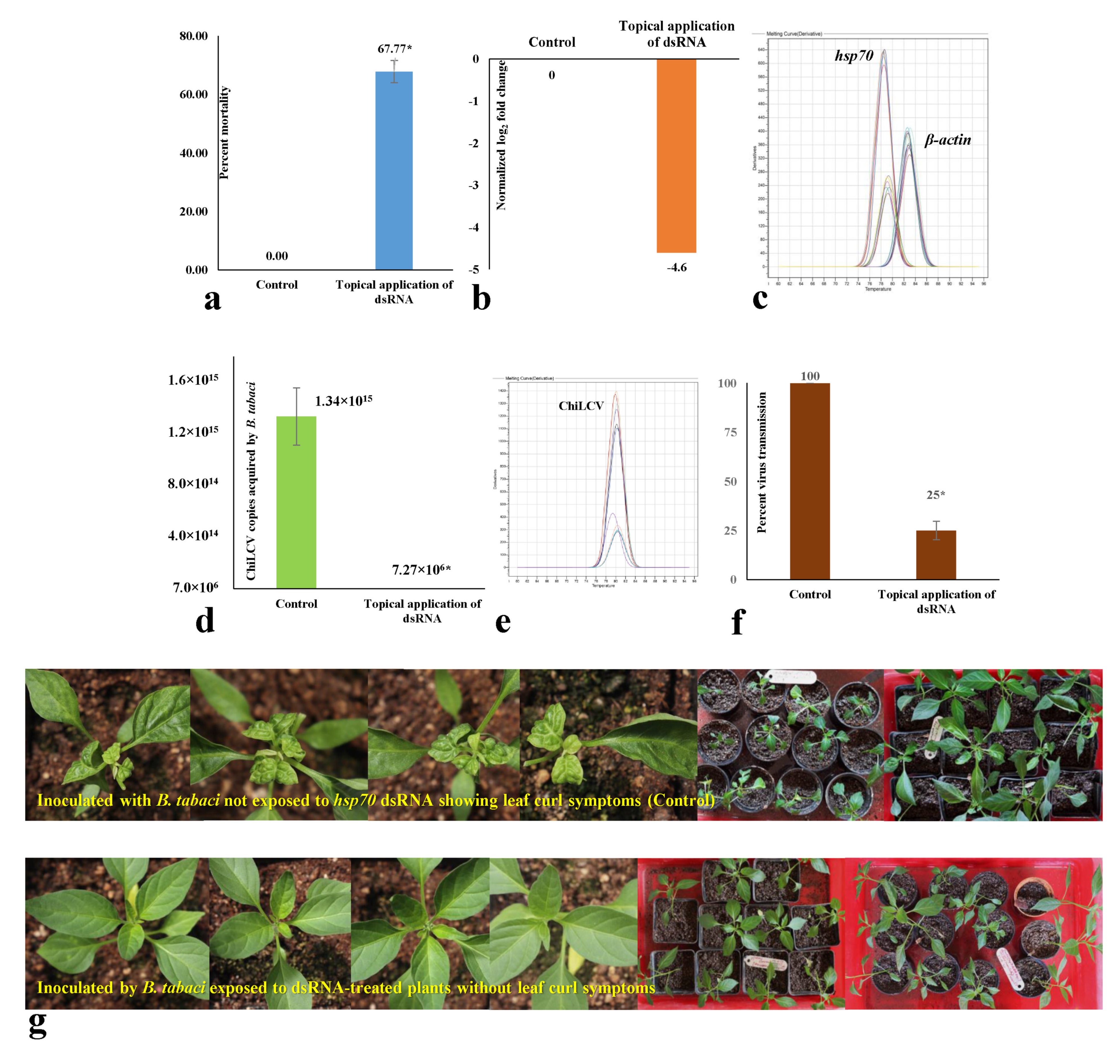
3.5. Effect of Topical Spray of hsp70 dsRNA on Mortality and Virus Transmission by B. tabaci under Semi-Field Conditions
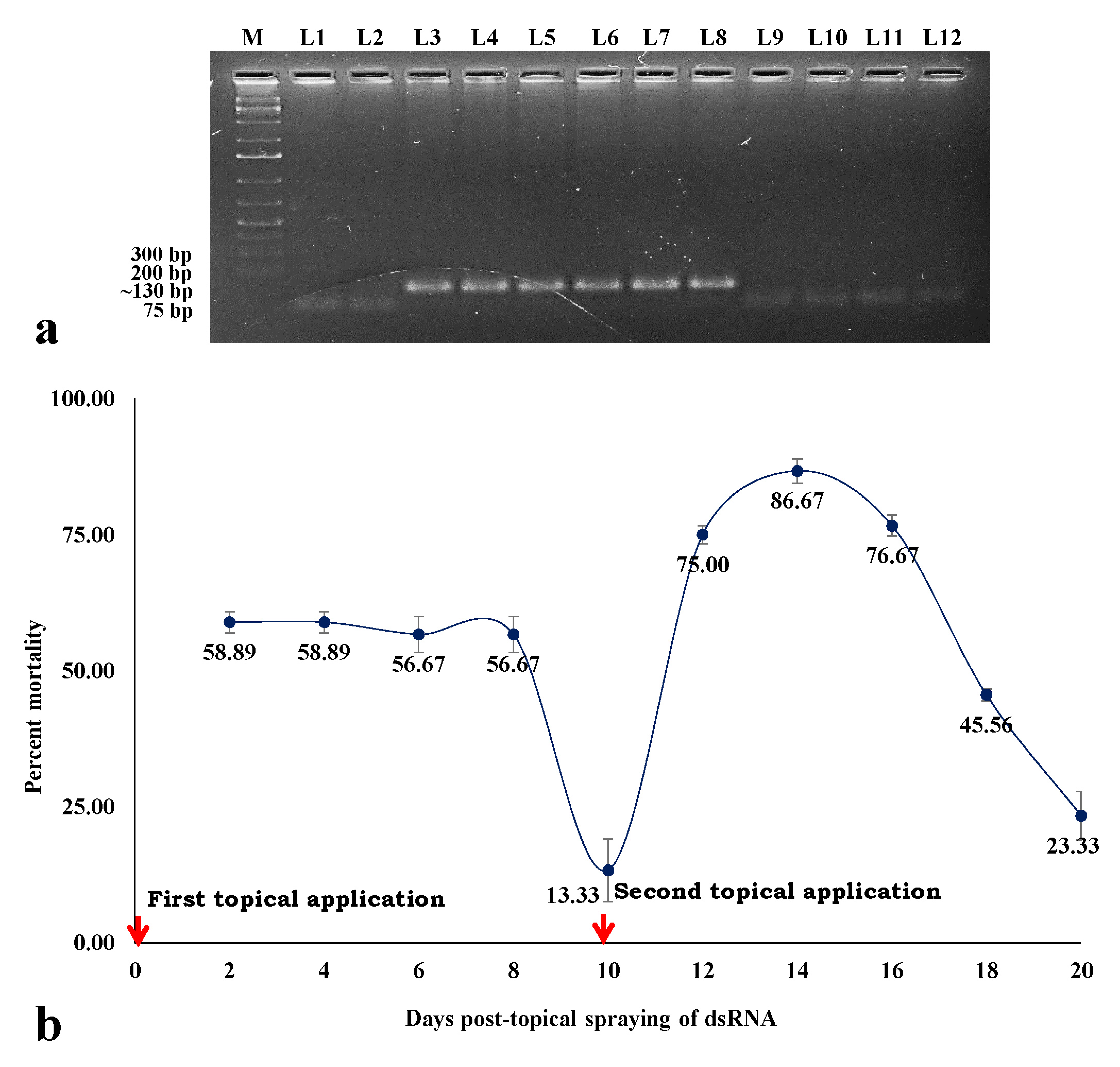
3.6. Stability and Persistent Efficacy of hsp70 dsRNA in Eradicating B. tabaci Population
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethical Approval
References
- Oliveira, M.R.V.; Henneberry, T.J.; Anderson, P. History, Current Status, and Collaborative Research Projects for Bemisia tabaci. Crop Prot. 2001, 20, 709–723. [Google Scholar] [CrossRef] [Green Version]
- Chowda-Reddy, R.; Kirankumar, M.; Seal, S.E.; Muniyappa, V.; Valand, G.B.; Govindappa, M.; Colvin, J. Bemisia tabaci Phylogenetic Groups in India and the Relative Transmission Efficacy of Tomato Leaf Curl Bangalore Virus by an Indigenous and an Exotic Population. J. Integr. Agric. 2012, 11, 235–248. [Google Scholar] [CrossRef]
- Legarrea, S.; Barman, A.; Marchant, W.; Diffie, S.; Srinivasan, R. Temporal Effects of a Begomovirus Infection and Host Plant Resistance on the Preference and Development of an Insect Vector, Bemisia tabaci, and Implications for Epidemics. PLoS ONE 2015, 10, e0142114. [Google Scholar] [CrossRef] [Green Version]
- Mugerwa, H.; Seal, S.; Wang, H.L.; Patel, M.V.; Kabaalu, R.; Omongo, C.A.; Alicai, T.; Tairo, F.; Ndunguru, J.; Sseruwagi, P.; et al. African Ancestry of New World, Bemisia tabaci-Whitefly Species. Sci. Rep. 2018, 8, 2734. [Google Scholar] [CrossRef] [Green Version]
- Jiu, M.; Hu, J.; Wang, L.J.; Dong, J.F.; Song, Y.Q.; Sun, H.Z. Cryptic Species Identification and Composition of Bemisia tabaci (Hemiptera: Aleyrodidae) Complex in Henan Province, China. J. Insect Sci. 2017, 17, 78–84. [Google Scholar] [CrossRef] [Green Version]
- Rehman, M.; Chakraborty, P.; Tanti, B.; Mandal, B.; Ghosh, A. Occurrence of a New Cryptic Species of Bemisia tabaci (Hemiptera: Aleyrodidae): An Updated Record of Cryptic Diversity in India. Phytoparasitica 2021, 49, 869–882. [Google Scholar] [CrossRef]
- Byrne, D.N.; Bellows, T.S. Whitefly Biology. Annu. Rev. Entomol. 1991, 36, 431–457. [Google Scholar] [CrossRef]
- Jones, D.R. Plant Viruses Transmitted by Whiteflies. Eur. J. Plant Pathol. 2003, 109, 195–219. [Google Scholar] [CrossRef]
- Navas-Castillo, J.; Fiallo-Olivé, E.; Sánchez-Campos, S. Emerging Virus Diseases Transmitted by Whiteflies. Annu. Rev. Phytopathol. 2011, 49, 219–248. [Google Scholar] [CrossRef]
- Brown, J.K.; Zerbini, F.M.; Navas-Castillo, J.; Moriones, E.; Ramos-Sobrinho, R.; Silva, J.C.; Fiallo-Olivé, E.; Briddon, R.W.; Hernández-Zepeda, C.; Idris, A.; et al. Revision of Begomovirus Taxonomy Based on Pairwise Sequence Comparisons. Arch. Virol. 2015, 160, 1593–1619. [Google Scholar] [CrossRef]
- Orfanidou, C.G.; Pappi, P.G.; Efthimiou, K.E.; Katis, N.I.; Maliogka, V.I. Transmission of Tomato Chlorosis Virus (ToCV) by Bemisia tabaci Biotype Q and Evaluation of Four Weed Species as Viral Sources. Plant Dis. 2016, 100, 2043–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanardo, L.G.; Carvalho, C.M. Cowpea mild mottle virus (Carlavirus, Betaflexiviridae): A review. Trop. Plant Pathol. 2017, 42, 417–430. [Google Scholar] [CrossRef]
- Ghosh, S.; Kanakala, S.; Lebedev, G.; Kontsedalov, S.; Silverman, D.; Alon, T.; Mor, N.; Sela, N.; Luria, N.; Dombrovsky, A.; et al. Transmission of a New Polerovirus Infecting Pepper by the Whitefly Bemisia tabaci. J. Virol. 2019, 93, e00488-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, T.M.; Inoue-Nagata, A.K.; Vida, A.H.; Ribeiro, S.G.; Nagata, T. The recombinant isolate of cucurbit aphid-borne yellows virus from Brazil is a polerovirus transmitted by whiteflies. Plant Pathol. 2020, 69, 1042–1050. [Google Scholar] [CrossRef]
- Pinheiro-Lima, B.; Pereira-Carvalho, R.C.; Alves-Freitas, D.M.T.; Kitajima, E.W.; Vidal, A.H.; Lacorte, C.; Godinho, M.T.; Fontenele, R.S.; Faria, J.C.; Abreu, E.F.M.; et al. Transmission of the Bean-Associated Cytorhabdovirus by the Whitefly Bemisia tabaci MEAM1. Viruses 2020, 12, 1028. [Google Scholar] [CrossRef]
- Cornejo-Franco, J.F.; Reyes-Proaño, E.G.; Mollov, D.; Mowery, J.; Quito-Avila, D.F. Transmission and Pathogenicity of Papaya Virus E: Insights from an Experimental Papaya Orchard. Plant Dis. 2022, 2017, 2467940. [Google Scholar] [CrossRef]
- Mansoor, S.; Briddon, R.W.; Zafar, Y.; Stanley, J. Geminivirus Disease Complexes: An Emerging Threat. Trends Plant Sci. 2003, 8, 128–134. [Google Scholar] [CrossRef]
- Briddon, R.W.; Patil, B.L.; Bagewadi, B.; Nawaz-ul-Rehman, M.S.; Fauquet, C.M. Distinct Evolutionary Histories of the DNA-A and DNA-B Components of Bipartite Begomoviruses. BMC. Evol. Biol. 2010, 10, 97. [Google Scholar] [CrossRef] [Green Version]
- Saeed, S.T.; Samad, A. Emerging Threats of Begomoviruses to the Cultivation of Medicinal and Aromatic Crops and Their Management Strategies. Virus Dis. 2017, 28, 1–17. [Google Scholar] [CrossRef] [Green Version]
- CABI. Capsicum Annuum (Bell Pepper). Invasive Species Compendium. Available online: https://www.cabi.org/isc/datasheet/15784 (accessed on 13 April 2021).
- Oraon, U.B.; Tarafdar, J. Occurrence and distribution of chilli leaf curl complex disease in West Bengal. Biomed. J. Sci. Tech. Res. 2018, 3, 3515–3519. [Google Scholar] [CrossRef] [Green Version]
- Kumar, Y.; Hallan, V.; Zaidi, A.A. Chilli Leaf Curl Palampur Virus Is a Distinct Begomovirus species Associated with a Betasatellite. Plant Pathol. 2011, 60, 1040–1047. [Google Scholar] [CrossRef]
- Khan, A.J.; Akhtar, S.; Al-Zaidi, A.M.; Singh, A.K.; Briddon, R.W. Genetic Diversity and Distribution of a Distinct Strain of Chili Leaf Curl Virus and Associated Betasatellite Infecting Tomato and Pepper in Oman. Virus Res. 2013, 177, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Senanayake, D.M.J.B.; Mandal, B.; Lodha, S.; Varma, A. First Report of Chilli Leaf Curl Virus Affecting Chilli in India. Plant Pathol. 2007, 56, 343. [Google Scholar] [CrossRef]
- Senanayake, D.M.J.B.; Jayasinghe, J.E.; Shilpi, S.; Wasala, S.K.; Mandal, B. A New Begomovirus–Betasatellite Complex Is Associated with Chilli Leaf Curl Disease in Sri Lanka. Virus Genes 2013, 46, 128–139. [Google Scholar] [CrossRef]
- Thakur, H.; Jindal, S.K.; Sharma, A.; Dhaliwal, M.S. Chilli Leaf Curl Virus Disease: A Serious Threat for Chilli Cultivation. J. Plant Dis. Prot. 2018, 125, 239–249. [Google Scholar] [CrossRef]
- Menike, G.D.N.; De Costa, D.M. Variation of Field Symptoms and Molecular Diversity of the Virus Isolates Associated with Chilli Leaf Curl Complex in Different Agroecological Regions of Sri Lanka. Trop. Agric. Res. 2017, 28, 144. [Google Scholar] [CrossRef] [Green Version]
- Geley, S.; Müller, C. RNAi: Ancient Mechanism with a Promising Future. Exp. Gerontol. 2004, 39, 985–998. [Google Scholar] [CrossRef]
- Rosen, R.; Kanakala, S.; Kliot, A.; Cathrin Pakkianathan, B.; Farich, B.A.; Santana-Magal, N.; Elimelech, M.; Kontsedalov, S.; Lebedev, G.; Cilia, M.; et al. Persistent, Circulative Transmission of Begomoviruses by Whitefly Vectors. Curr. Opin. Virol. 2015, 15, 1–8. [Google Scholar] [CrossRef]
- Czosnek, H.; Hariton-Shalev, A.; Sobol, I.; Gorovits, R.; Ghanim, M. The Incredible Journey of Begomoviruses in Their Whitefly Vector. Viruses 2017, 9, 273. [Google Scholar] [CrossRef]
- Götz, M.; Popovski, S.; Kollenberg, M.; Gorovits, R.; Brown, J.K.; Cicero, J.M.; Czosnek, H.; Winter, S.; Ghanim, M. Implication of Bemisia tabaci Heat Shock protein 70 in Begomovirus-Whitefly Interactions. J. Virol. 2012, 86, 13241–13252. [Google Scholar] [CrossRef] [Green Version]
- Kanakala, S.; Ghanim, M. Implication of the Whitefly Bemisia tabaci Cyclophilin B Protein in the Transmission of Tomato Yellow Leaf Curl Virus. Front. Plant Sci. 2016, 7, 1702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rana, V.S.; Popli, S.; Saurav, G.K.; Raina, H.S.; Chaubey, R.; Ramamurthy, V.V.; Rajagopal, R. A Bemisia tabaci Midgut Protein Interacts with Begomoviruses and Plays a Role in Virus Transmission. Cell. Microbiol. 2016, 18, 663–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.Z.; Shi, M.; Huang, Y.C.; Wang, X.W.; Stanley, D.; Chen, X.X. A Peptidoglycan Recognition Protein Acts in Whitefly (Bemisia tabaci) Immunity and Involves in Begomovirus Acquisition. Sci. Rep. 2016, 6, 37806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nekkanti, A. Changes in Transcriptome of Whitefly (Bemisia Tabaci Gennadius) Associated with Begomovirus Infection. Master’s Thesis, Post Graduate School, ICAR-Indian Agricultural Research Insitute, New Delhi, India, 2021; p. 66. [Google Scholar]
- Hartl, F.U.; Hayer-Hartl, M. Molecular Chaperones in the Cytosol: From Nascent Chain to Folded Protein. Science 2002, 295, 1852–1858. [Google Scholar] [CrossRef] [Green Version]
- Packard, M.; Mathew, D.; Budnik, V. FASt Remodeling of Synapses in Drosophila. Curr. Opin. Neurobiol. 2003, 13, 527–534. [Google Scholar] [CrossRef]
- Kristiansen, L.V.; Hortsch, M. Fasciclin II: The NCAM Ortholog in Drosophila melanogaster. Adv. Exp. Med. Biol. 2010, 663, 387–401. [Google Scholar] [CrossRef]
- Hotta, K.; Motoi, Y.; Okutani, A.; Kaku, Y.; Noguchi, A.; Inoue, S.; Yamada, A. Role of GPI-Anchored NCAM-120 in Rabies Virus Infection. Microbes Infect. 2007, 9, 167–174. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Chandrashekar, K.; Thakur, N.; Verma, P.C.; Borgio, J.F.; Singh, P.K.; Tuli, R. RNA interference for the Control of Whiteflies (Bemisia tabaci) by Oral Route. J. Biosci. 2011, 36, 153–161. [Google Scholar] [CrossRef]
- Li, H.; Guan, R.; Guo, H.; Miao, X. New Insights into an RNAi Approach for Plant Defense Against Piercing-Sucking and Stem-Borer Insect Pests. Plant Cell Environ. 2015, 38, 2277–2285. [Google Scholar] [CrossRef]
- Vyas, M.; Raza, A.; Ali, M.Y.; Ashraf, M.A.; Mansoor, S.; Shahid, A.A.; Brown, J.K. Knock Down of Whitefly Gut Gene Expression and Mortality by Orally Delivered Gut Gene-Specific dsRNAs. PLoS ONE 2017, 12, e0168921. [Google Scholar] [CrossRef]
- Bar, L.; Czosnek, H.; Sobol, I.; Ghanim, M.; Hariton Shalev, A. Down Regulation of Dystrophin Expression in Pupae of the Whitefly Bemisia tabaci Inhibits the Emergence of Adults. Insect Mol. Biol. 2019, 28, 662–675. [Google Scholar] [CrossRef]
- Akhter, A.; Qazi, J.; Saeed, M.; Mansoor, S. A Severe Leaf Curl Disease on Chilies in Pakistan Is Associated with Multiple Begomovirus Components. Plant Dis. 2009, 93, 962. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.J.; Donahue, K.; Koh, Y.; Martin, R.R.; Choi, M.Y. Microbial-Based Double-Stranded RNA Production to Develop Cost-Effective RNA interference Application for Insect Pest Management. Int. J. Insect Sci. 2019, 11, 1179543319840323. [Google Scholar] [CrossRef] [PubMed]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, Weighting, and Phylogenetic Utility of Mitochondrial Gene Sequences and a Compilation of Conserved Polymerase Chain Reaction Primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Roy, B.; Chakraborty, P.; Ghosh, A. How many begomovirus copies are acquired and inoculated by its vector, whitefly (Bemisia tabaci) during feeding? PLoS ONE 2021, 16, e0258933. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of Plant DNA from Fresh Tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Abdullah, N.M.M.; Singh, J.; Sohal, B.S. Behavioral Hormoligosis in Oviposition Preference of Bemisia tabaci on Cotton. Pestic. Biochem. Physiol. 2006, 84, 10–16. [Google Scholar] [CrossRef]
- Bacci, L.; Crespo, A.L.; Galvan, T.L.; Pereira, E.J.; Picanço, M.C.; Silva, G.A.; Chediak, M. Toxicity of Insecticides to the Sweetpotato Whitefly (Hemiptera: Aleyrodidae) and Its Natural Enemies. Pest Manag. Sci. 2007, 63, 699–706. [Google Scholar] [CrossRef]
- Houndété, T.A.; Kétoh, G.K.; Hema, O.S.; Brévault, T.; Glitho, I.A.; Martin, T. Insecticide Resistance in Field Populations of Bemisia tabaci (Hemiptera: Aleyrodidae) in West Africa. Pest Manag. Sci. 2010, 66, 1181–1185. [Google Scholar] [CrossRef]
- Sen, G.L.; Blau, H.M. A Brief History of RNAi: The Silence of the Genes. FASEB J. 2006, 20, 1293–1299. [Google Scholar] [CrossRef] [PubMed]
- Baum, J.A.; Bogaert, T.; Clinton, W.; Heck, G.R.; Feldmann, P.; Ilagan, O.; Johnson, S.; Plaetinck, G.; Munyikwa, T.; Pleau, M.; et al. Control of Coleopteran Insect Pests Through RNA interference. Nat. Biotechnol. 2007, 25, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Huvenne, H.; Smagghe, G. Mechanisms of dsRNA Uptake in Insects and Potential of RNAi for Pest Control: A Review. J. Insect Physiol. 2010, 56, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Hasegawa, D.K.; Kaur, N.; Kliot, A.; Pinheiro, P.V.; Luan, J.; Stensmyr, M.C.; Zheng, Y.; Liu, W.; Sun, H.; et al. The Draft Genome of Whitefly Bemisia tabaci MEAM1, a Global Crop Pest, Provides Novel Insights into Virus Transmission, Host Adaptation, and Insecticide Resistance. BMC Biol. 2016, 14, 110. [Google Scholar] [CrossRef]
- Ding, T.B.; Li, J.; Chen, E.H.; Niu, J.Z.; Chu, D. Transcriptome Profiling of the Whitefly Bemisia tabaci MED in Response to Single Infection of Tomato Yellow Leaf Curl Virus, Tomato Chlorosis Virus, and Their Co-Infection. Front. Physiol. 2019, 10, 302. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.; Farooq, M.; Malik, H.J.; Amin, I.; Scheffler, B.E.; Scheffler, J.A.; Liu, S.S.; Mansoor, S. Whole Genome Sequencing of Asia II 1 Species of Whitefly Reveals That Genes Involved in Virus Transmission and Insecticide Resistance Have Genetic Variances Between Asia II 1 and MEAM1 Species. BMC Genom. 2019, 20, 507. [Google Scholar] [CrossRef] [Green Version]
- Grover, S.; Jindal, V.; Banta, G.; Taning, C.N.T.; Smagghe, G.; Christiaens, O. Potential of RNA interference in the Study and Management of the Whitefly, Bemisia tabaci. Arch. Insect Biochem. Physiol. 2019, 100, e21522. [Google Scholar] [CrossRef]
- Kanakala, S.; Kontsedalov, S.; Lebedev, G.; Ghanim, M. Plant-Mediated Silencing of the Whitefly Bemisia tabaci Cyclophilin B and Heat Shock protein 70 Impairs Insect Development and Virus Transmission. Front. Physiol. 2019, 10, 557. [Google Scholar] [CrossRef]
- Kaur, R.; Gupta, M.; Singh, S.; Joshi, N.; Sharma, A. Enhancing RNAi Efficiency to Decipher the Functional Response of Potential Genes in Bemisia tabaci Asia II-1 (Gennadius) Through dsRNA Feeding Assays. Front. Physiol. 2020, 11, 123. [Google Scholar] [CrossRef]
- Tenllado, F.; Martínez-García, B.; Vargas, M.; Díaz-Ruíz, J.R. Crude Extracts of Bacterially Expressed dsRNA Can Be Used to Protect Plants Against Virus Infections. BMC Biotechnol. 2003, 3, 3. [Google Scholar] [CrossRef] [Green Version]
- Leelesh, R.S.; Rieske, L.K. Oral Ingestion of Bacterially Expressed dsRNA Can Silence Genes and Cause Mortality in a Highly Invasive, Tree-Killing Pest, the Emerald Ash Borer. Insects 2020, 11, 440. [Google Scholar] [CrossRef] [PubMed]
- Cherenko, V.A.; Filipenko, E.A.; Golubeva, T.S. Silencing of Nicotiana Benthamiana Phytoendesaturase Using dsRNA Synthesized In Vivo. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2021; p. 723. [Google Scholar] [CrossRef]
- Gregersen, N.; Bross, P.; Andrese, B.S.; Pedersen, C.B.; Corydon, T.J.; Bolund, L. The Role of Chaperone-Assisted Folding and Quality Control in Inborn Errors of Metabolism: Protein Folding Disorders. J. Inherit. Metab. Dis. 2001, 24, 189–212. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.P. Recruitment of Hsp70 Chaperones: A Crucial Part of Viral Survival Strategies. Rev. Physiol. Biochem. Pharmacol. 2005, 153, 1–46. [Google Scholar] [CrossRef]
- Serva, S.; Nagy, P.D. Proteomics Analysis of the Tombusvirus Replicase: Hsp70 Molecular Chaperone Is Associated with the Replicase and Enhances Viral RNA Replication. J. Virol. 2006, 80, 2162–2169. [Google Scholar] [CrossRef] [Green Version]
- Ivanovic, T.; Agosto, M.A.; Chandran, K.; Nibert, M.L. A Role for Molecular Chaperone Hsc70 in Reovirus Outer Capsid Disassembly. J. Biol. Chem. 2007, 282, 12210–12219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czosnek, H. Tomato Yellow Leaf Curl Virus Disease. In Management, Molecular Biology, Breeding for Resistance; Czosnek, H., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; Volume 440. [Google Scholar] [CrossRef]
- Gorovits, R.; Moshe, A.; Ghanim, M.; Czosnek, H. Recruitment of the Host Plant Heat Shock protein 70 by Tomato Yellow Leaf Curl Virus Coat Protein Is Required for Virus Infection. PLoS ONE 2013, 8, e70280. [Google Scholar] [CrossRef]
- Jiu, M.; Zhou, X.P.; Liu, S.S. Acquisition and Transmission of Tobacco Curly Shoot Virus by the B Biotype of the Whitefly, Bemisia tabaci. Acta Phytophylacica Sin. 2006, 33, 168–172. [Google Scholar] [CrossRef]
- Guo, X.J.; Feng, J.N. Comparisons of Expression Levels of Heat Shock Proteins (hsp70 and hsp90) from Anaphothrips obscurus (Thysanoptera: Thripidae) in Polymorphic Adults Exposed to Different Heat Shock Treatments. J. Insect Sci. 2018, 18, 15. [Google Scholar] [CrossRef]
- Palli, S.R. RNA interference in Colorado Potato Beetle: Steps Toward Development of dsRNA as a Commercial Insecticide. Curr. Opin. Insect Sci. 2014, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Zotti, M.J.; Smagghe, G. RNAi Technology for Insect Management and Protection of Beneficial Insects from Diseases: Lessons, Challenges and Risk Assessments. Neotrop. Entomol. 2015, 44, 197–213. [Google Scholar] [CrossRef]
- Garbutt, J.S.; Reynolds, S.E. Induction of RNA interference Genes by Double-Stranded RNA. Implications for Susceptibility to RNA interference. Insect Biochem. Mol. Biol. 2012, 42, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Shukla, J.N.; Kalsi, M.; Sethi, A.; Narva, K.E.; Fishilevich, E.; Singh, S.; Mogilicherla, K.; Palli, S.R. Reduced Stability and Intracellular Transport of dsRNA Contribute to Poor RNAi Response in Lepidopteran Insects. RNA Biol. 2016, 13, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Christiaens, O.; Whyard, S.; Vélez, A.M.; Smagghe, G. Double-Stranded RNA Technology to Control Insect Pests: Current Status and Challenges. Front. Plant Sci. 2020, 11, 451. [Google Scholar] [CrossRef] [PubMed]
- Namgial, T.; Kaldis, A.; Chakraborty, S.; Voloudakis, A. Topical Application of Double-Stranded RNA Molecules Containing Sequences of Tomato Leaf Curl Virus and Cucumber Mosaic Virus Confers Protection Against the Cognate Viruses. Physiol. Mol. Plant Pathol. 2019, 108, 101432. [Google Scholar] [CrossRef]
- Worrall, E.A.; Bravo-Cazar, A.; Nilon, A.T.; Fletcher, S.J.; Robinson, K.E.; Carr, J.P.; Mitter, N. Exogenous Application of RNAi-Inducing Double-Stranded RNA Inhibits Aphid-Mediated Transmission of a Plant Virus. Front. Plant Sci. 2019, 10, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konakalla, N.C.; Bag, S.; Deraniyagala, A.S.; Culbreath, A.K.; Pappu, H.R. Induction of Plant Resistance in Tobacco (Nicotiana tabacum) Against Tomato Spotted Wilt Orthotospovirus Through Foliar Application of dsRNA. Viruses 2021, 13, 662. [Google Scholar] [CrossRef]
- Konakalla, N.C.; Kaldis, A.; Berbati, M.; Masarapu, H.; Voloudakis, A.E. Exogenous Application of Double-Stranded RNA Molecules from TMV p126 and CP Genes Confers Resistance Against TMV in Tobacco. Planta 2016, 244, 961–969. [Google Scholar] [CrossRef]
- Kaldis, A.; Berbati, M.; Melita, O.; Reppa, C.; Holeva, M.; Otten, P.; Voloudakis, A. Exogenously Applied dsRNA Molecules Deriving from the Zucchini Yellow Mosaic Virus (ZYMV) Genome Move Systemically and Protect Cucurbits Against ZYMV. Mol. Plant Pathol. 2018, 19, 883–895. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Kumar, M. Leaf Curl Disease: A Significant Constraint in the Production of Tomato in India. In Advances in Plant Pathology; Kumatu, J.N., Ed.; IntechOpen: London, UK, 2018; pp. 93–101. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chakraborty, P.; Ghosh, A. Topical Spray of dsRNA Induces Mortality and Inhibits Chilli Leaf Curl Virus Transmission by Bemisia tabaci Asia II 1. Cells 2022, 11, 833. https://doi.org/10.3390/cells11050833
Chakraborty P, Ghosh A. Topical Spray of dsRNA Induces Mortality and Inhibits Chilli Leaf Curl Virus Transmission by Bemisia tabaci Asia II 1. Cells. 2022; 11(5):833. https://doi.org/10.3390/cells11050833
Chicago/Turabian StyleChakraborty, Prosenjit, and Amalendu Ghosh. 2022. "Topical Spray of dsRNA Induces Mortality and Inhibits Chilli Leaf Curl Virus Transmission by Bemisia tabaci Asia II 1" Cells 11, no. 5: 833. https://doi.org/10.3390/cells11050833
APA StyleChakraborty, P., & Ghosh, A. (2022). Topical Spray of dsRNA Induces Mortality and Inhibits Chilli Leaf Curl Virus Transmission by Bemisia tabaci Asia II 1. Cells, 11(5), 833. https://doi.org/10.3390/cells11050833