
High-Throughput Analysis of Astrocyte Cultures Shows Prevention of Reactive Astrogliosis by the Multi-Nutrient Combination Fortasyn Connect

 ,
, 
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Primary Hippocampal Astrocyte Culture
2.2. Cytokine and FC Treatment
2.3. Immunofluorescence
2.4. High-Throughput Screening
2.5. Analysis of Reactive Astrogliosis
2.6. Viability/Cytotoxicity Assay
2.7. SDS-PAGE Immunoblotting
2.8. Data Collection and Statistical Analysis
3. Results
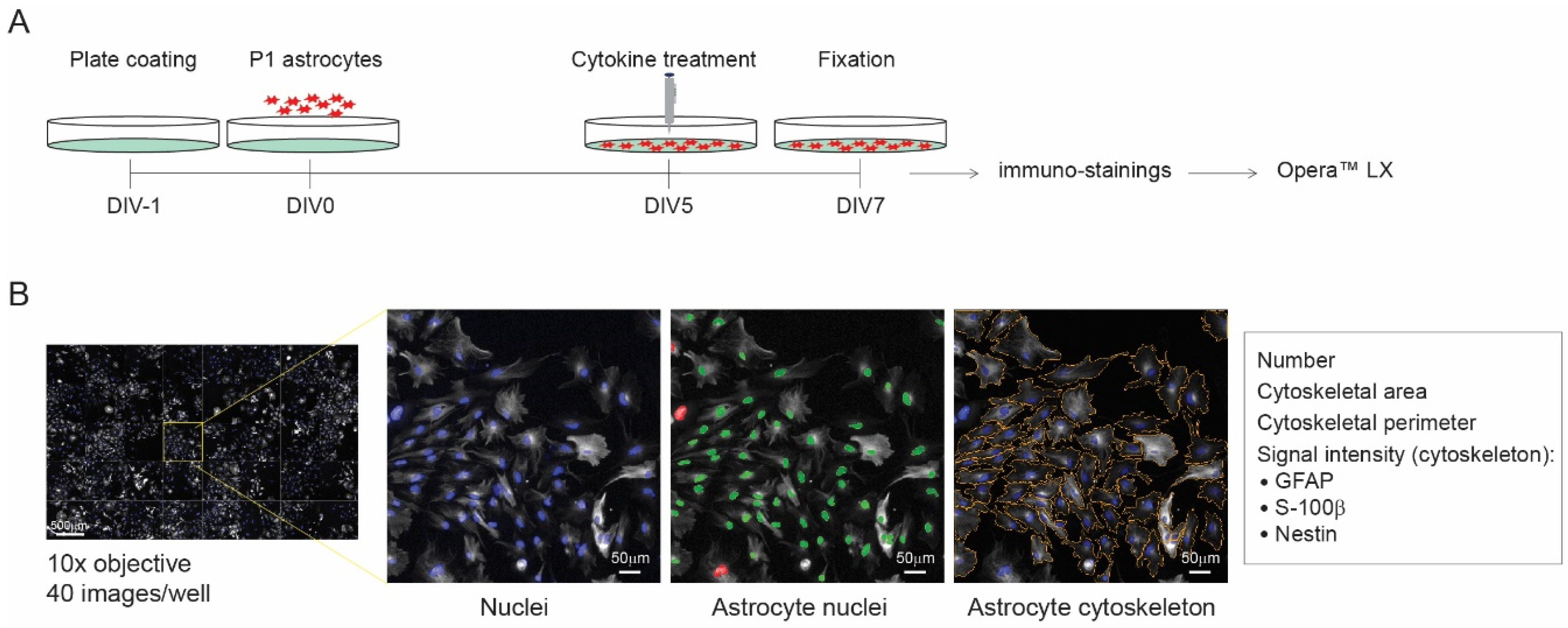
3.1. Development of a Reactive Astrogliosis In Vitro Model; the Effect of the Plate Coating
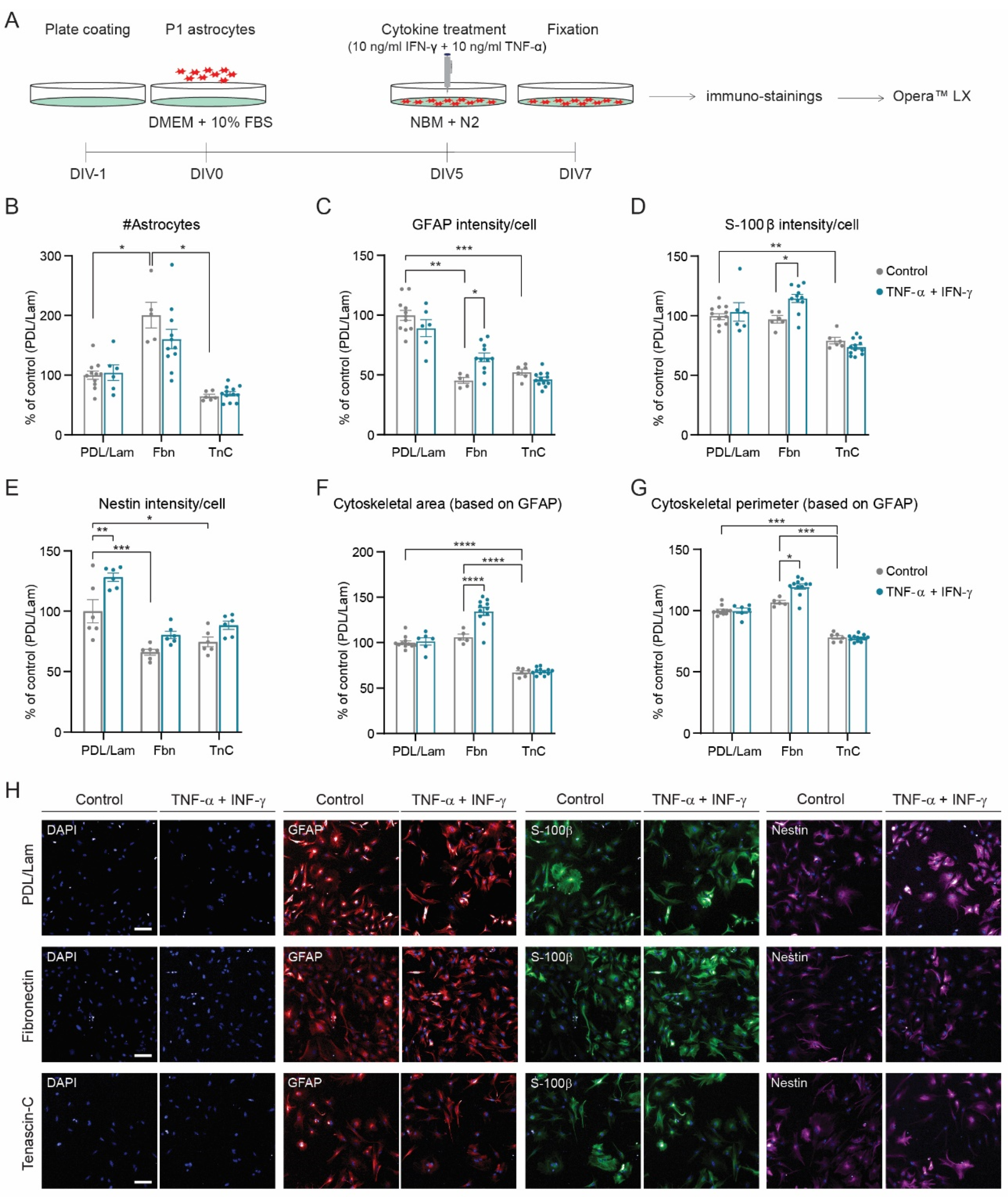
3.2. Development of a Reactive Astrogliosis In Vitro Model; the Effect of Culture Media

3.3. Differential Effect of TNF-α and IFN-γ Concentration on the Development of a Reactive Astrogliosis Model
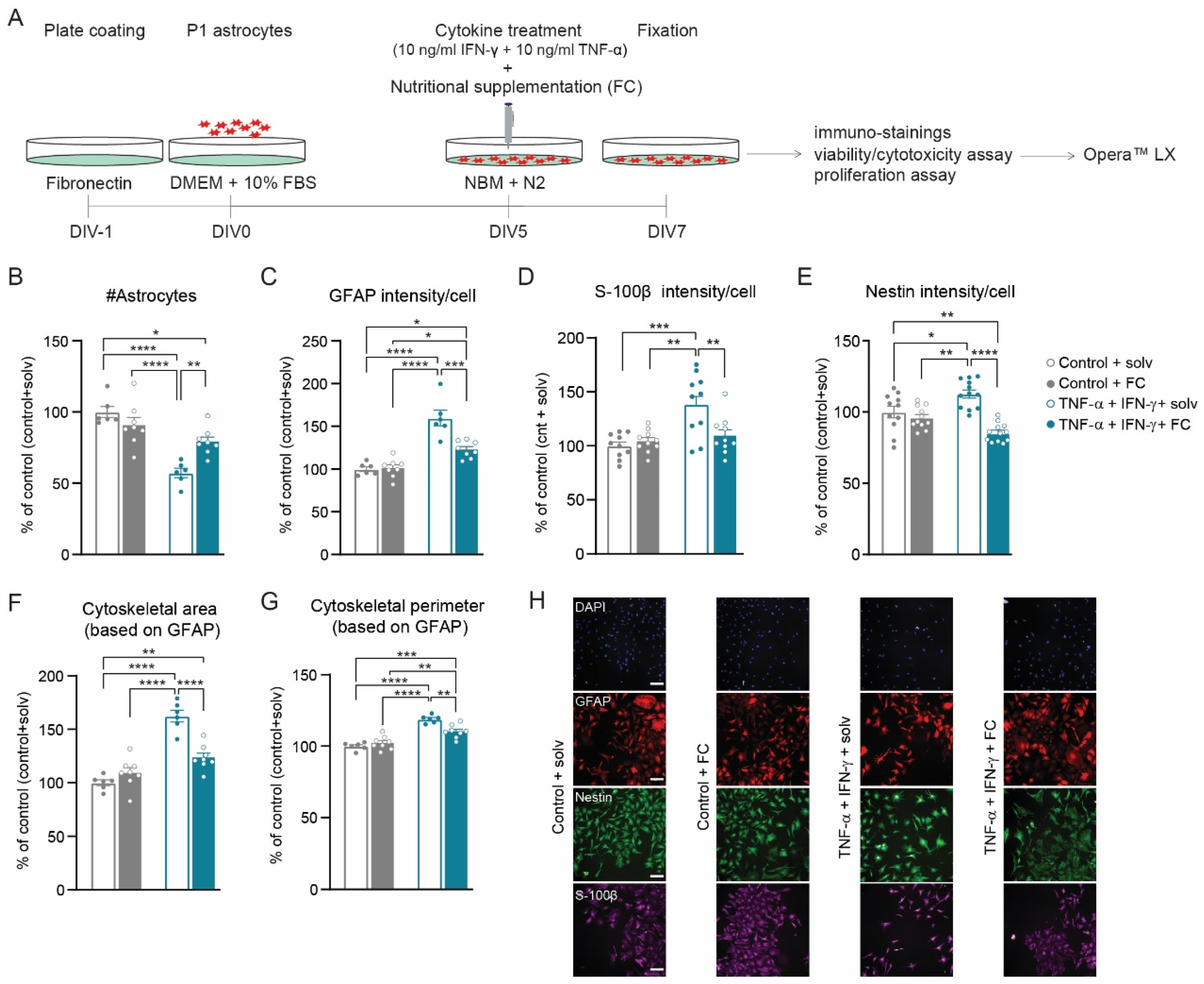
3.4. FC Prevents Cytokine-Induced Reactive Astrogliosis
4. Discussion
4.1. AstroScan: High-Throughput Analysis of Astrocyte Reactivity In Vitro
4.2. FC Supplementation Prevents Cytokine-Induced Reactive Astrogliosis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Parpura, V.; Verkhratsky, A. Homeostatic Function of Astrocytes: Ca2+ and Na+ Signalling. Transl. Neurosci. 2012, 3, 334–344. [Google Scholar] [CrossRef] [PubMed]
- Perea, G.; Navarrete, M.; Araque, A. Tripartite Synapses: Astrocytes Process and Control Synaptic Information. Trends Neurosci. 2009, 32, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Pellerin, L.; Magistretti, P.J. Neuroenergetics: Calling Upon Astrocytes to Satisfy Hungry Neurons. Neuroscientist 2004, 10, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Bazargani, N.; Attwell, D. Astrocyte Calcium Signaling: The Third Wave. Nat. Neurosci. 2016, 19, 182–189. [Google Scholar] [CrossRef]
- Eroglu, C.; Barres, B.A. Regulation of Synaptic Connectivity by Glia. Nature 2010, 468, 223–231. [Google Scholar] [CrossRef]
- Perea, G.; Sur, M.; Araque, A. Neuron-Glia Networks: Integral Gear of Brain Function. Front. Cell. Neurosci. 2014, 8, 378. [Google Scholar] [CrossRef] [Green Version]
- Escartin, C.; Galea, E.; Lakatos, A.; O’Callaghan, J.P.; Petzold, G.C.; Serrano-Pozo, A.; Steinhäuser, C.; Volterra, A.; Carmignoto, G.; Agarwal, A.; et al. Reactive Astrocyte Nomenclature, Definitions, and Future Directions. Nat. Neurosci. 2021, 24, 312–325. [Google Scholar] [CrossRef]
- Sofroniew, M.V. Astrogliosis. Cold Spring Harb. Perspect. Biol. 2015, 7, a020420. [Google Scholar] [CrossRef] [Green Version]
- Sofroniew, M.V. Molecular Dissection of Reactive Astrogliosis and Glial Scar Formation. Trends Neurosci. 2009, 32, 638–647. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Li, Z.; Noori, A.; Hyman, B.T.; Serrano-Pozo, A. Meta-Analysis of Mouse Transcriptomic Studies Supports a Context-Dependent Astrocyte Reaction in Acute CNS Injury versus Neurodegeneration. J. Neuroinflamm. 2020, 17, 227. [Google Scholar] [CrossRef]
- Colangelo, A.M.; Alberghina, L.; Papa, M. Astrogliosis as a Therapeutic Target for Neurodegenerative Diseases. Neurosci. Lett. 2014, 565, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barcia, C.; Ros, C.M.; Annese, V.; Gómez, A.; Ros-Bernal, F.; Aguado-Llera, D.; Martínez-Pagán, M.E.; de Pablos, V.; Fernandez-Villalba, E.; Herrero, M.T. IFN-γ Signaling, with the Synergistic Contribution of TNF-α, Mediates Cell Specific Microglial and Astroglial Activation in Experimental Models of Parkinson’s Disease. Cell Death Dis. 2011, 2, e142. [Google Scholar] [CrossRef] [Green Version]
- Clarke, L.E.; Liddelow, S.A.; Chakraborty, C.; Münch, A.E.; Heiman, M.; Barres, B.A. Normal Aging Induces A1-like Astrocyte Reactivity. Proc. Natl. Acad. Sci. USA 2018, 115, E1896–E1905. [Google Scholar] [CrossRef] [Green Version]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.-S.; Peterson, T.C.; et al. Neurotoxic Reactive Astrocytes Are Induced by Activated Microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef]
- Zamanian, J.L.; Xu, L.; Foo, L.C.; Nouri, N.; Zhou, L.; Giffard, R.G.; Barres, B.A. Genomic Analysis of Reactive Astrogliosis. J. Neurosci. 2012, 32, 6391–6410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben Haim, L.; Carrillo-de Sauvage, M.-A.; Ceyzeriat, K.; Escartin, C. Elusive Roles for Reactive Astrocytes in Neurodegenerative Diseases. Front. Cell. Neurosci. 2015, 9, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridet, J.L.; Privat, A.; Malhotra, S.K.; Gage, F.H. Reactive Astrocytes: Cellular and Molecular Cues to Biological Function. Trends Neurosci. 1997, 20, 570–577. [Google Scholar] [CrossRef]
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: Biology and Pathology. Acta Neuropathol. 2010, 119, 7–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigetomi, E.; Saito, K.; Sano, F.; Koizumi, S. Aberrant Calcium Signals in Reactive Astrocytes: A Key Process in Neurological Disorders. Int. J. Mol. Sci. 2019, 20, 996. [Google Scholar] [CrossRef] [Green Version]
- Stevens, B.; Allen, N.J.; Vazquez, L.E.; Howell, G.R.; Christopherson, K.S.; Nouri, N.; Micheva, K.D.; Mehalow, A.K.; Huberman, A.D.; Stafford, B.; et al. The Classical Complement Cascade Mediates CNS Synapse Elimination. Cell 2007, 131, 1164–1178. [Google Scholar] [CrossRef] [Green Version]
- Zumkehr, J.; Rodriguez-Ortiz, C.J.; Cheng, D.; Kieu, Z.; Wai, T.; Hawkins, C.; Kilian, J.; Lim, S.L.; Medeiros, R.; Kitazawa, M. Ceftriaxone Ameliorates Tau Pathology and Cognitive Decline via Restoration of Glial Glutamate Transporter in a Mouse Model of Alzheimer’s Disease. Neurobiol. Aging 2015, 36, 2260–2271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Q.; Li, Y.; Chopp, M. Bone Marrow Stromal Cells Increase Astrocyte Survival via Upregulation of Phosphoinositide 3-Kinase/Threonine Protein Kinase and Mitogen-Activated Protein Kinase Kinase/Extracellular Signal-Regulated Kinase Pathways and Stimulate Astrocyte Trophic Factor Gene Expression after Anaerobic Insult. Neuroscience 2005, 136, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Zador, Z.; Stiver, S.; Wang, V.; Manley, G.T. Role of Aquaporin-4 in Cerebral Edema and Stroke. In Aquaporins; Beitz, E., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 159–170. ISBN 978-3-540-79885-9. [Google Scholar]
- Viejo, L.; Noori, A.; Merrill, E.; Das, S.; Hyman, B.T.; Serrano-Pozo, A. Systematic Review of Human Post-mortem Immunohistochemical Studies and Bioinformatics Analyses Unveil the Complexity of Astrocyte Reaction in Alzheimer’s Disease. Neuropathol. Appl. Neurobiol. 2021, 48, e12753. [Google Scholar] [CrossRef] [PubMed]
- Halliday, G.M.; Stevens, C.H. Glia: Initiators and Progressors of Pathology in Parkinson’s Disease: Glia in Parkinson’s Disease. Mov. Disord. 2011, 26, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhao, R.; Gao, K.; Wei, Z.; Yaoyao Yin, M.; Ting Lau, L.; Chui, D.; Cheung Hoi Yu, A. Astrocytes: Implications for Neuroinflammatory Pathogenesis of Alzheimers Disease. Curr. Alzheimer Res. 2011, 8, 67–80. [Google Scholar] [CrossRef]
- Rossi, D.; Volterra, A. Astrocytic Dysfunction: Insights on the Role in Neurodegeneration. Brain Res. Bull. 2009, 80, 224–232. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Parpura, V.; Rodriguez-Arellano, J.J.; Zorec, R. Astroglia in Alzheimer’s Disease. In Neuroglia in Neurodegenerative Diseases; Verkhratsky, A., Ho, M.S., Zorec, R., Parpura, V., Eds.; Advances in Experimental Medicine and Biology; Springer: Singapore, 2019; Volume 1175, pp. 273–324. ISBN 9789811399121. [Google Scholar]
- Sadick, J.S.; Liddelow, S.A. Don’t Forget Astrocytes When Targeting Alzheimer’s Disease. Br. J. Pharmacol. 2019, 176, 3585–3598. [Google Scholar] [CrossRef]
- van Deijk, A.-L.F.; Broersen, L.M.; Verkuyl, J.M.; Smit, A.B.; Verheijen, M.H.G. High Content Analysis of Hippocampal Neuron-Astrocyte Co-Cultures Shows a Positive Effect of Fortasyn Connect on Neuronal Survival and Postsynaptic Maturation. Front. Neurosci. 2017, 11, 440. [Google Scholar] [CrossRef]
- Soininen, H.; Solomon, A.; Visser, P.J.; Hendrix, S.B.; Blennow, K.; Kivipelto, M.; Hartmann, T.; The LipiDiDiet Clinical Study Group. 36-month LipiDiDiet Multinutrient Clinical Trial in Prodromal Alzheimer’s Disease. Alzheimers Dement. 2021, 17, 29–40. [Google Scholar] [CrossRef]
- Soininen, H.; Solomon, A.; Visser, P.J.; Hendrix, S.B.; Blennow, K.; Kivipelto, M.; Hartmann, T.; Hallikainen, I.; Hallikainen, M.; Helisalmi, S.; et al. 24-Month Intervention with a Specific Multinutrient in People with Prodromal Alzheimer’s Disease (LipiDiDiet): A Randomised, Double-Blind, Controlled Trial. Lancet Neurol. 2017, 16, 965–975. [Google Scholar] [CrossRef] [Green Version]
- Scheltens, P.; Twisk, J.W.R.; Blesa, R.; Scarpini, E.; von Arnim, C.A.F.; Bongers, A.; Harrison, J.; Swinkels, S.H.N.; Stam, C.J.; de Waal, H.; et al. Efficacy of Souvenaid in Mild Alzheimer’s Disease: Results from a Randomized, Controlled Trial. J. Alzheimers Dis. 2012, 31, 225–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheltens, P.; Kamphuis, P.J.G.H.; Verhey, F.R.J.; Olde Rikkert, M.G.M.; Wurtman, R.J.; Wilkinson, D.; Twisk, J.W.R.; Kurz, A. Efficacy of a Medical Food in Mild Alzheimer’s Disease: A Randomized, Controlled Trial. Alzheimers Dement. 2010, 6, 1–10.e1. [Google Scholar] [CrossRef] [PubMed]
- de Waal, H.; Stam, C.J.; Lansbergen, M.M.; Wieggers, R.L.; Kamphuis, P.J.G.H.; Scheltens, P.; Maestú, F.; van Straaten, E.C.W. The Effect of Souvenaid on Functional Brain Network Organisation in Patients with Mild Alzheimer’s Disease: A Randomised Controlled Study. PLoS ONE 2014, 9, e86558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zerbi, V.; Jansen, D.; Wiesmann, M.; Fang, X.; Broersen, L.M.; Veltien, A.; Heerschap, A.; Kiliaan, A.J. Multinutrient Diets Improve Cerebral Perfusion and Neuroprotection in a Murine Model of Alzheimer’s Disease. Neurobiol. Aging 2014, 35, 600–613. [Google Scholar] [CrossRef] [PubMed]
- Pallier, P.N.; Poddighe, L.; Zbarsky, V.; Kostusiak, M.; Choudhury, R.; Hart, T.; Burguillos, M.A.; Musbahi, O.; Groenendijk, M.; Sijben, J.W.; et al. A Nutrient Combination Designed to Enhance Synapse Formation and Function Improves Outcome in Experimental Spinal Cord Injury. Neurobiol. Dis. 2015, 82, 504–515. [Google Scholar] [CrossRef]
- Price, P.J.; Brewer, G.J. Serum-Free Media for Neural Cell Cultures: Adult and Embryonic. In Protocols for Neural Cell Culture; Humana Press: Totowa, NJ, USA, 2001; pp. 255–264. ISBN 978-1-59259-207-4. [Google Scholar]
- Trindade, P.; Loiola, E.C.; Gasparotto, J.; Ribeiro, C.T.; Cardozo, P.L.; Devalle, S.; Salerno, J.A.; Ornelas, I.M.; Ledur, P.F.; Ribeiro, F.M.; et al. Short and Long TNF-alpha Exposure Recapitulates Canonical Astrogliosis Events in Human-induced Pluripotent Stem Cells-derived Astrocytes. Glia 2020, 68, 1396–1409. [Google Scholar] [CrossRef]
- Hashioka, S.; McGeer, E.; Miyaoka, T.; Wake, R.; Horiguchi, J.; McGeer, P. Interferon-γ-induced neurotoxicity of human astrocytes. CNS Neurol. Disord.—Drug Targets 2015, 14, 251–256. [Google Scholar] [CrossRef] [PubMed]
- John, G.R.; Lee, S.C.; Brosnan, C.F. Cytokines: Powerful Regulators of Glial Cell Activation. The Neuroscientist 2003, 9, 10–22. [Google Scholar] [CrossRef]
- Hsiao, H.-Y.; Mak, O.-T.; Yang, C.-S.; Liu, Y.-P.; Fang, K.-M.; Tzeng, S.-F. TNF-α/IFN-γ-Induced INOS Expression Increased by Prostaglandin E2 in Rat Primary Astrocytes via EP2-Evoked CAMP/PKA and Intracellular Calcium Signaling. Glia 2007, 55, 214–223. [Google Scholar] [CrossRef]
- Zhao, J.; O’Connor, T.; Vassar, R. The Contribution of Activated Astrocytes to Aβ Production: Implications for Alzheimer’s Disease Pathogenesis. J. Neuroinflamm. 2011, 8, 150. [Google Scholar] [CrossRef] [Green Version]
- Holley, J.E.; Gveric, D.; Whatmore, J.L.; Gutowski, N.J. Tenascin C Induces a Quiescent Phenotype in Cultured Adult Human Astrocytes. Glia 2005, 52, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.M.; Milner, R.; Crocker, S.J. Extracellular Matrix Composition Determines Astrocyte Responses to Mechanical and Inflammatory Stimuli. Neurosci. Lett. 2015, 600, 104–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perriot, S.; Mathias, A.; Perriard, G.; Canales, M.; Jonkmans, N.; Merienne, N.; Meunier, C.; El Kassar, L.; Perrier, A.L.; Laplaud, D.-A.; et al. Human Induced Pluripotent Stem Cell-Derived Astrocytes Are Differentially Activated by Multiple Sclerosis-Associated Cytokines. Stem Cell Rep. 2018, 11, 1199–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selmaj, K.W.; Farooq, M.; Norton, W.T.; Raine, C.S.; Brosnan, C.F. Proliferation of Astrocytes in Vitro in Response to Cytokines. A Primary Role for Tumor Necrosis Factor. J. Immunol. Baltim. Md 1950 1990, 144, 129–135. [Google Scholar]
- Foo, L.C.; Allen, N.J.; Bushong, E.A.; Ventura, P.B.; Chung, W.-S.; Zhou, L.; Cahoy, J.D.; Daneman, R.; Zong, H.; Ellisman, M.H.; et al. Development of a Method for the Purification and Culture of Rodent Astrocytes. Neuron 2011, 71, 799–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J. New Insights into GFAP Negative Astrocytes in Calbindin D28k Immunoreactive Astrocytes. Brain Sci. 2018, 8, 143. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.L.; King, V.R.; Curran, O.E.; Dyall, S.C.; Ward, R.E.; Lal, N.; Priestley, J.V.; Michael-Titus, A.T. A Combination of Intravenous and Dietary Docosahexaenoic Acid Significantly Improves Outcome after Spinal Cord Injury. Brain 2007, 130, 3004–3019. [Google Scholar] [CrossRef] [Green Version]
- Bonizzi, G.; Karin, M. The Two NF-ΚB Activation Pathways and Their Role in Innate and Adaptive Immunity. Trends Immunol. 2004, 25, 280–288. [Google Scholar] [CrossRef]
- Saggu, R.; Schumacher, T.; Gerich, F.; Rakers, C.; Tai, K.; Delekate, A.; Petzold, G.C. Astroglial NF-KB Contributes to White Matter Damage and Cognitive Impairment in a Mouse Model of Vascular Dementia. Acta Neuropathol. Commun. 2016, 4, 76. [Google Scholar] [CrossRef] [Green Version]
- Furman, J.L.; Sama, D.M.; Gant, J.C.; Beckett, T.L.; Murphy, M.P.; Bachstetter, A.D.; Van Eldik, L.J.; Norris, C.M. Targeting Astrocytes Ameliorates Neurologic Changes in a Mouse Model of Alzheimer’s Disease. J. Neurosci. 2012, 32, 16129–16140. [Google Scholar] [CrossRef] [Green Version]
- Abdul, H.M.; Sama, M.A.; Furman, J.L.; Mathis, D.M.; Beckett, T.L.; Weidner, A.M.; Patel, E.S.; Baig, I.; Murphy, M.P.; LeVine, H.; et al. Cognitive Decline in Alzheimer’s Disease Is Associated with Selective Changes in Calcineurin/NFAT Signaling. J. Neurosci. 2009, 29, 12957–12969. [Google Scholar] [CrossRef] [PubMed]
- Sama, M.A.; Mathis, D.M.; Furman, J.L.; Abdul, H.M.; Artiushin, I.A.; Kraner, S.D.; Norris, C.M. Interleukin-1β-Dependent Signaling between Astrocytes and Neurons Depends Critically on Astrocytic Calcineurin/NFAT Activity. J. Biol. Chem. 2008, 283, 21953–21964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyvärinen, T.; Hagman, S.; Ristola, M.; Sukki, L.; Veijula, K.; Kreutzer, J.; Kallio, P.; Narkilahti, S. Co-Stimulation with IL-1β and TNF-α Induces an Inflammatory Reactive Astrocyte Phenotype with Neurosupportive Characteristics in a Human Pluripotent Stem Cell Model System. Sci. Rep. 2019, 9, 16944. [Google Scholar] [CrossRef] [PubMed]
- Olabarria, M.; Noristani, H.N.; Verkhratsky, A.; Rodríguez, J.J. Concomitant Astroglial Atrophy and Astrogliosis in a Triple Transgenic Animal Model of Alzheimer’s Disease. Glia 2010, 58, 831–838. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhang, Q.; Yu, Z.; Zhang, L.; Tian, D.; Zhu, S.; Bu, B.; Xie, M.; Wang, W. Inhibiting Cell Cycle Progression Reduces Reactive Astrogliosis Initiated by Scratch Injury in Vitro and by Cerebral Ischemia in Vivo. Glia 2007, 55, 546–558. [Google Scholar] [CrossRef]
- Hol, E.M.; Pekny, M. Glial Fibrillary Acidic Protein (GFAP) and the Astrocyte Intermediate Filament System in Diseases of the Central Nervous System. Curr. Opin. Cell Biol. 2015, 32, 121–130. [Google Scholar] [CrossRef]
- Michetti, F.; D’Ambrosi, N.; Toesca, A.; Puglisi, M.A.; Serrano, A.; Marchese, E.; Corvino, V.; Geloso, M.C. The S100B Story: From Biomarker to Active Factor in Neural Injury. J. Neurochem. 2019, 148, 168–187. [Google Scholar] [CrossRef] [Green Version]
- Moreels, M.; Vandenabeele, F.; Dumont, D.; Robben, J.; Lambrichts, I. Alpha-Smooth Muscle Actin (α-SMA) and Nestin Expression in Reactive Astrocytes in Multiple Sclerosis Lesions: Potential Regulatory Role of Transforming Growth Factor-Beta 1 (TGF-Β1). Neuropathol. Appl. Neurobiol. 2008, 34, 532–546. [Google Scholar] [CrossRef]
- Escartin, C.; Guillemaud, O.; Carrillo-de Sauvage, M. Questions and (Some) Answers on Reactive Astrocytes. Glia 2019, 67, 2221–2247. [Google Scholar] [CrossRef]
- Faber-Elman, A.; Lavie, V.; Schvartz, I.; Shaltiel, S.; Schwartz, M. Vitronectin Overrides a Negative Effect of TNF-Alpha on Astrocyte Migration. FASEB J. 1995, 9, 1605–1613. [Google Scholar] [CrossRef]
- Summers, L.; Kangwantas, K.; Nguyen, L.; Kielty, C.; Pinteaux, E. Adhesion to the Extracellular Matrix Is Required for Interleukin-1 Beta Actions Leading to Reactive Phenotype in Rat Astrocytes. Mol. Cell. Neurosci. 2010, 44, 272–281. [Google Scholar] [CrossRef] [PubMed]
- McKeon, R.; Schreiber, R.; Rudge, J.; Silver, J. Reduction of Neurite Outgrowth in a Model of Glial Scarring Following CNS Injury Is Correlated with the Expression of Inhibitory Molecules on Reactive Astrocytes. J. Neurosci. 1991, 11, 3398–3411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoffels, J.M.J.; de Jonge, J.C.; Stancic, M.; Nomden, A.; van Strien, M.E.; Ma, D.; Šišková, Z.; Maier, O.; ffrench-Constant, C.; Franklin, R.J.M.; et al. Fibronectin Aggregation in Multiple Sclerosis Lesions Impairs Remyelination. Brain 2013, 136, 116–131. [Google Scholar] [CrossRef] [Green Version]
- Sobel, R.A.; Mitchell, M.E. Fibronectin in Multiple Sclerosis Lesions. Am. J. Pathol. 1989, 135, 161–168. [Google Scholar] [PubMed]
- van Horssen, J.; Bö, L.; Vos, C.M.P.; Virtanen, I.; de Vries, H.E. Basement Membrane Proteins in Multiple Sclerosis-Associated Inflammatory Cuffs: Potential Role in Influx and Transport of Leukocytes. J. Neuropathol. Exp. Neurol. 2005, 64, 722–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tom, V.J. Astrocyte-Associated Fibronectin Is Critical for Axonal Regeneration in Adult White Matter. J. Neurosci. 2004, 24, 9282–9290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Codeluppi, S.; Gregory, E.N.; Kjell, J.; Wigerblad, G.; Olson, L.; Svensson, C.I. Influence of Rat Substrain and Growth Conditions on the Characteristics of Primary Cultures of Adult Rat Spinal Cord Astrocytes. J. Neurosci. Methods 2011, 197, 118–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Valk, J.; Brunner, D.; De Smet, K.; Fex Svenningsen, Å.; Honegger, P.; Knudsen, L.E.; Lindl, T.; Noraberg, J.; Price, A.; Scarino, M.L.; et al. Optimization of Chemically Defined Cell Culture Media—Replacing Fetal Bovine Serum in Mammalian in Vitro Methods. Toxicol. In Vitro 2010, 24, 1053–1063. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.G.; Soltysiak, K.A.; Gao, Z.-G.; Chang, T.-S.; Chung, E.; Jacobson, K.A. Tumor Necrosis Factor α-Induced Apoptosis in Astrocytes Is Prevented by the Activation of P2Y6, but Not P2Y4 Nucleotide Receptors. Biochem. Pharmacol. 2003, 65, 923–931. [Google Scholar] [CrossRef]
- Hostenbach, S.; Cambron, M.; D’haeseleer, M.; Kooijman, R.; De Keyser, J. Astrocyte Loss and Astrogliosis in Neuroinflammatory Disorders. Neurosci. Lett. 2014, 565, 39–41. [Google Scholar] [CrossRef]
- Broersen, L.M.; Kuipers, A.A.M.; Balvers, M.; van Wijk, N.; Savelkoul, P.J.M.; de Wilde, M.C.; van der Beek, E.M.; Sijben, J.W.C.; Hageman, R.J.J.; Kamphuis, P.J.G.H.; et al. A Specific Multi-Nutrient Diet Reduces Alzheimer-Like Pathology in Young Adult AβPPswe/PS1dE9 Mice. J. Alzheimers Dis. 2012, 33, 177–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Pardo, P.; Broersen, L.M.; Kliest, T.; van Wijk, N.; Attali, A.; Garssen, J.; Kraneveld, A.D. Additive Effects of Levodopa and a Neurorestorative Diet in a Mouse Model of Parkinson’s Disease. Front. Aging Neurosci. 2018, 10, 237. [Google Scholar] [CrossRef] [PubMed]
- Thau-Zuchman, O.; Gomes, R.N.; Dyall, S.C.; Davies, M.; Priestley, J.V.; Groenendijk, M.; De Wilde, M.C.; Tremoleda, J.L.; Michael-Titus, A.T. Brain Phospholipid Precursors Administered Post-Injury Reduce Tissue Damage and Improve Neurological Outcome in Experimental Traumatic Brain Injury. J. Neurotrauma 2019, 36, 25–42. [Google Scholar] [CrossRef] [PubMed]
- Wiesmann, M.; Zinnhardt, B.; Reinhardt, D.; Eligehausen, S.; Wachsmuth, L.; Hermann, S.; Dederen, P.J.; Hellwich, M.; Kuhlmann, M.T.; Broersen, L.M.; et al. A Specific Dietary Intervention to Restore Brain Structure and Function after Ischemic Stroke. Theranostics 2017, 7, 493–512. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-ΚB Signaling in Inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, S.L.; Kearns, J.D.; Zadorozhnaya, V.; Lynch, C.; O’Dea, E.; Boldin, M.P.; Ma, A.; Baltimore, D.; Hoffmann, A. Encoding NF-ΚB Temporal Control in Response to TNF: Distinct Roles for the Negative Regulators IκBα and A20. Genes Dev. 2008, 22, 2093–2101. [Google Scholar] [CrossRef] [Green Version]
- Adamson, A.; Boddington, C.; Downton, P.; Rowe, W.; Bagnall, J.; Lam, C.; Maya-Mendoza, A.; Schmidt, L.; Harper, C.V.; Spiller, D.G.; et al. Signal Transduction Controls Heterogeneous NF-ΚB Dynamics and Target Gene Expression through Cytokine-Specific Refractory States. Nat. Commun. 2016, 7, 12057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, I.-M.; Yang, C.-H.; Yang, C.-M. Docosahexaenoic Acid Reduces Linoleic Acid Induced Monocyte Chemoattractant Protein-1 Expression via PPARγ and Nuclear Factor-ΚB Pathway in Retinal Pigment Epithelial Cells. Mol. Nutr. Food Res. 2014, 58, 2053–2065. [Google Scholar] [CrossRef] [PubMed]
- Marion-Letellier, R.; Butler, M.; Déchelotte, P.; Playford, R.J.; Ghosh, S. Comparison of Cytokine Modulation by Natural Peroxisome Proliferator–Activated Receptor γ Ligands with Synthetic Ligands in Intestinal-like Caco-2 Cells and Human Dendritic Cells—Potential for Dietary Modulation of Peroxisome Proliferator–Activated Receptor γ in Intestinal Inflammation. Am. J. Clin. Nutr. 2008, 87, 939–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, S.-N.; Huang, W.; Hall, J.C.E.; Ward, R.E.; Priestley, J.V.; Michael-Titus, A.T. The Acute Administration of Eicosapentaenoic Acid Is Neuroprotective after Spinal Cord Compression Injury in Rats. Prostaglandins Leukot. Essent. Fat. Acids PLEFA 2010, 83, 193–201. [Google Scholar] [CrossRef]
- Kamphuis, P.J.G.H.; Scheltens, P. Can Nutrients Prevent or Delay Onset of Alzheimer’s Disease? J. Alzheimers Dis. 2010, 20, 765–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Wijk, N.; Broersen, L.M.; de Wilde, M.C.; Hageman, R.J.J.; Groenendijk, M.; Sijben, J.W.C.; Kamphuis, P.J.G.H. Targeting Synaptic Dysfunction in Alzheimer’s Disease by Administering a Specific Nutrient Combination. J. Alzheimers Dis. 2013, 38, 459–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 0.2× FC (µM) | 0.05× FC (µM) | |
|---|---|---|
| DHA | 2.88 | 0.72 |
| EPA | 2.02 | 0.51 |
| Uridine | 10 | 2.5 |
| Choline | 4 | 1 |
| Vitamin B6 | 2 | 0.5 |
| Vitamin B12 | 0.02 | 0.005 |
| Vitamin B9 | 3 | 0.75 |
| PC | 5 | 1.25 |
| Vitamin C | 15 | 3.75 |
| Vitamin E | 4 | 1 |
| Selenium | 0.02 | 0.005 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badia-Soteras, A.; de Vries, J.; Dykstra, W.; Broersen, L.M.; Verkuyl, J.M.; Smit, A.B.; Verheijen, M.H.G. High-Throughput Analysis of Astrocyte Cultures Shows Prevention of Reactive Astrogliosis by the Multi-Nutrient Combination Fortasyn Connect. Cells 2022, 11, 1428. https://doi.org/10.3390/cells11091428
Badia-Soteras A, de Vries J, Dykstra W, Broersen LM, Verkuyl JM, Smit AB, Verheijen MHG. High-Throughput Analysis of Astrocyte Cultures Shows Prevention of Reactive Astrogliosis by the Multi-Nutrient Combination Fortasyn Connect. Cells. 2022; 11(9):1428. https://doi.org/10.3390/cells11091428
Chicago/Turabian StyleBadia-Soteras, Aina, Janneke de Vries, Werner Dykstra, Laus M. Broersen, Jan Martin Verkuyl, August B. Smit, and Mark H. G. Verheijen. 2022. "High-Throughput Analysis of Astrocyte Cultures Shows Prevention of Reactive Astrogliosis by the Multi-Nutrient Combination Fortasyn Connect" Cells 11, no. 9: 1428. https://doi.org/10.3390/cells11091428
APA StyleBadia-Soteras, A., de Vries, J., Dykstra, W., Broersen, L. M., Verkuyl, J. M., Smit, A. B., & Verheijen, M. H. G. (2022). High-Throughput Analysis of Astrocyte Cultures Shows Prevention of Reactive Astrogliosis by the Multi-Nutrient Combination Fortasyn Connect. Cells, 11(9), 1428. https://doi.org/10.3390/cells11091428