
Notch2 Blockade Mitigates Methotrexate Chemotherapy-Induced Bone Loss and Marrow Adiposity




Abstract
:1. Introduction
2. Materials and Methods
2.1. Rat MTX Chemotherapy Time-Course Study
2.2. PCR Array Analysis
2.3. Anti-Notch2 Treatment Study
2.4. Micro-Computed Tomography (Micro-CT)
2.5. Histological Cell Density Analyses
2.6. RNA Isolation and Quantitative RT-PCR Analyses of Gene Expression
2.7. Immunohistochemistry
2.8. Ex Vivo CFU-f/ALP Assays
2.9. Ex Vivo Osteogenesis and Mineralisation Assays
2.10. Ex Vivo Adipogenesis Assays
2.11. Ex Vivo Osteoclastogenesis Assays
2.12. Western Blot
2.13. Statistics Analyses
3. Results
3.1. MTX Chemotherapy Altered mRNA Expression of Key Regulatory Factors for Bone Homeostasis
3.2. Time Course Analyses of MTX Treatment Effects on mRNA Expression Levels of Notch Receptors and Target Genes
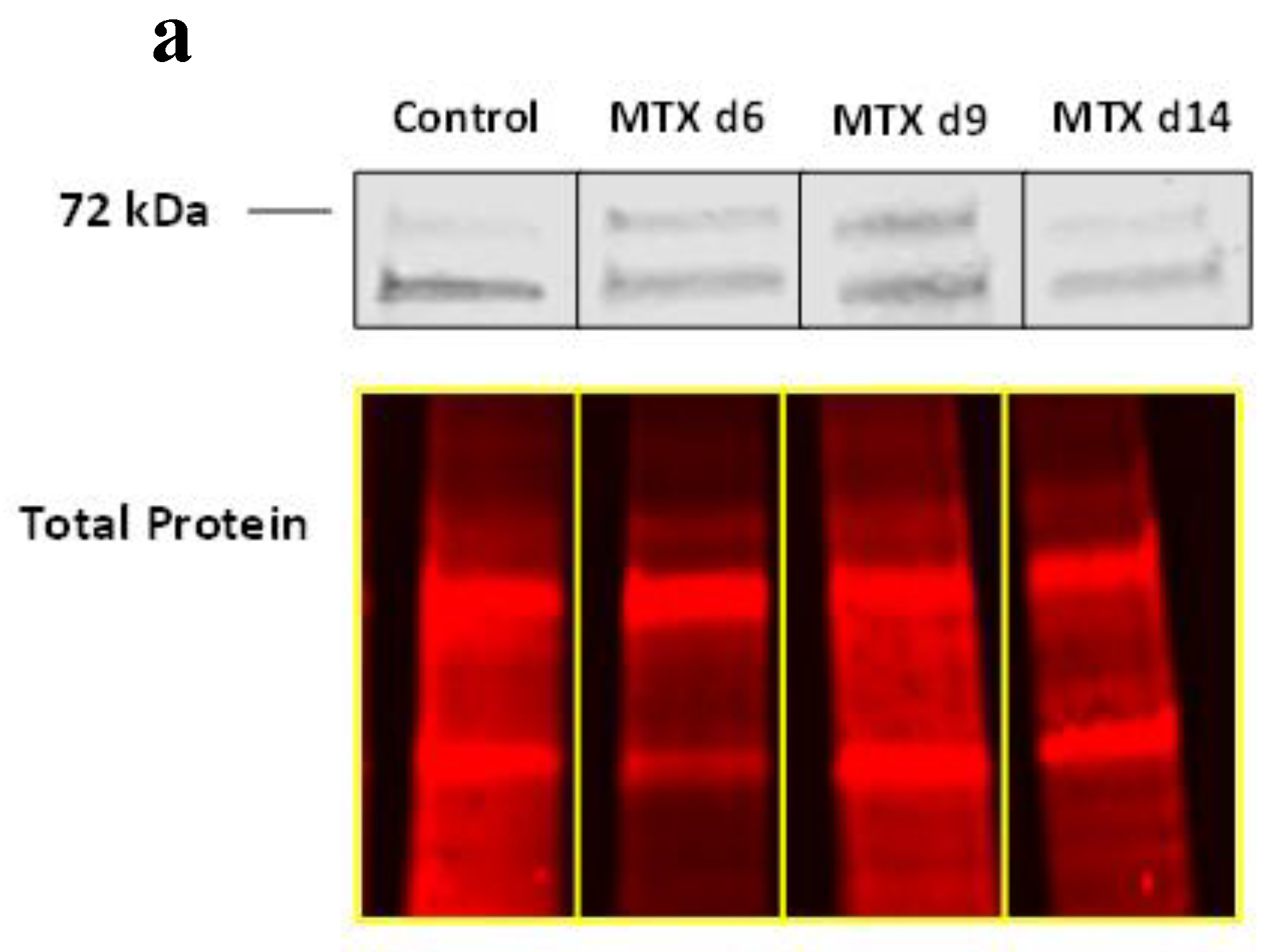
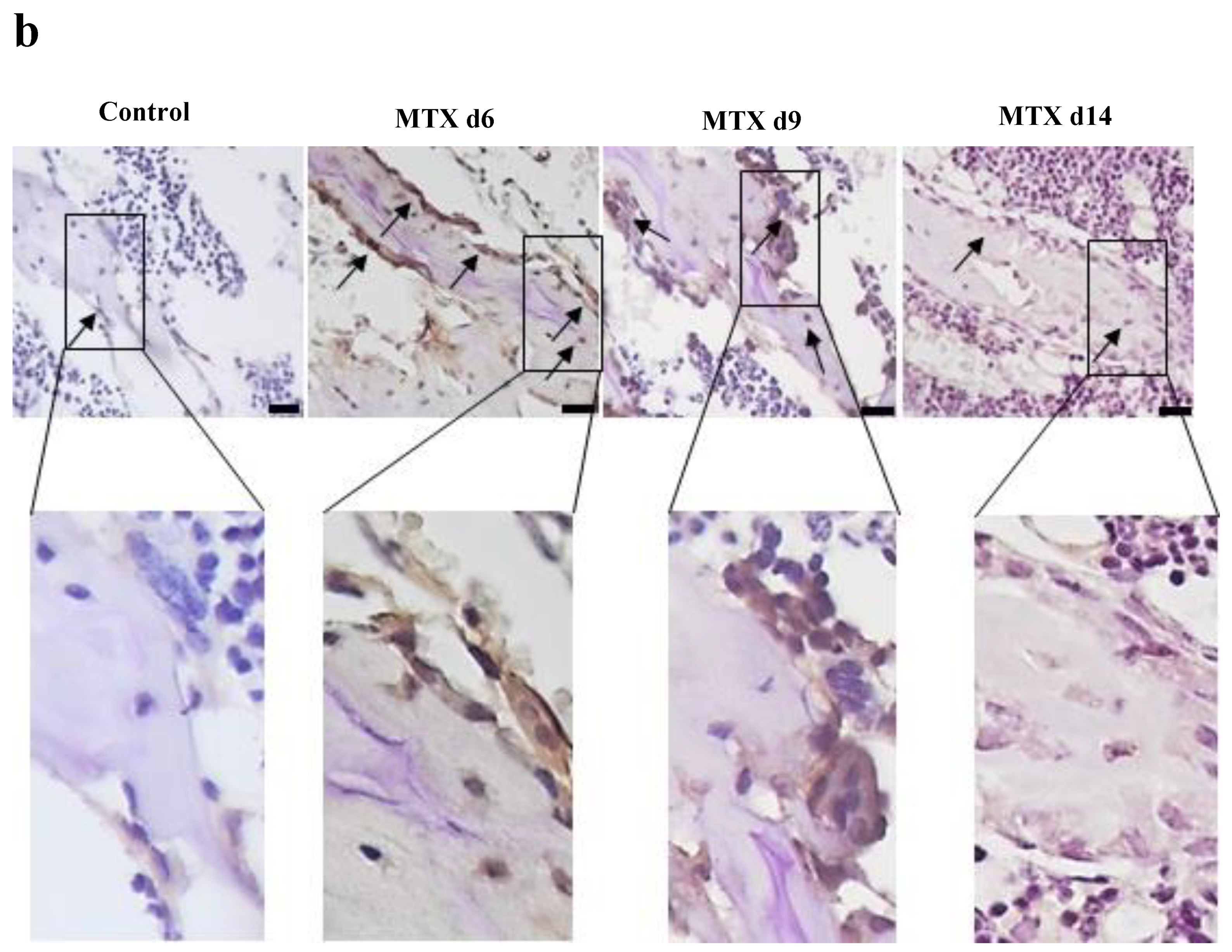
3.3. MTX Treatment Induced NICD2 Protein Expression Predominantly in Osteoblasts and Osteocytes in Metaphysis
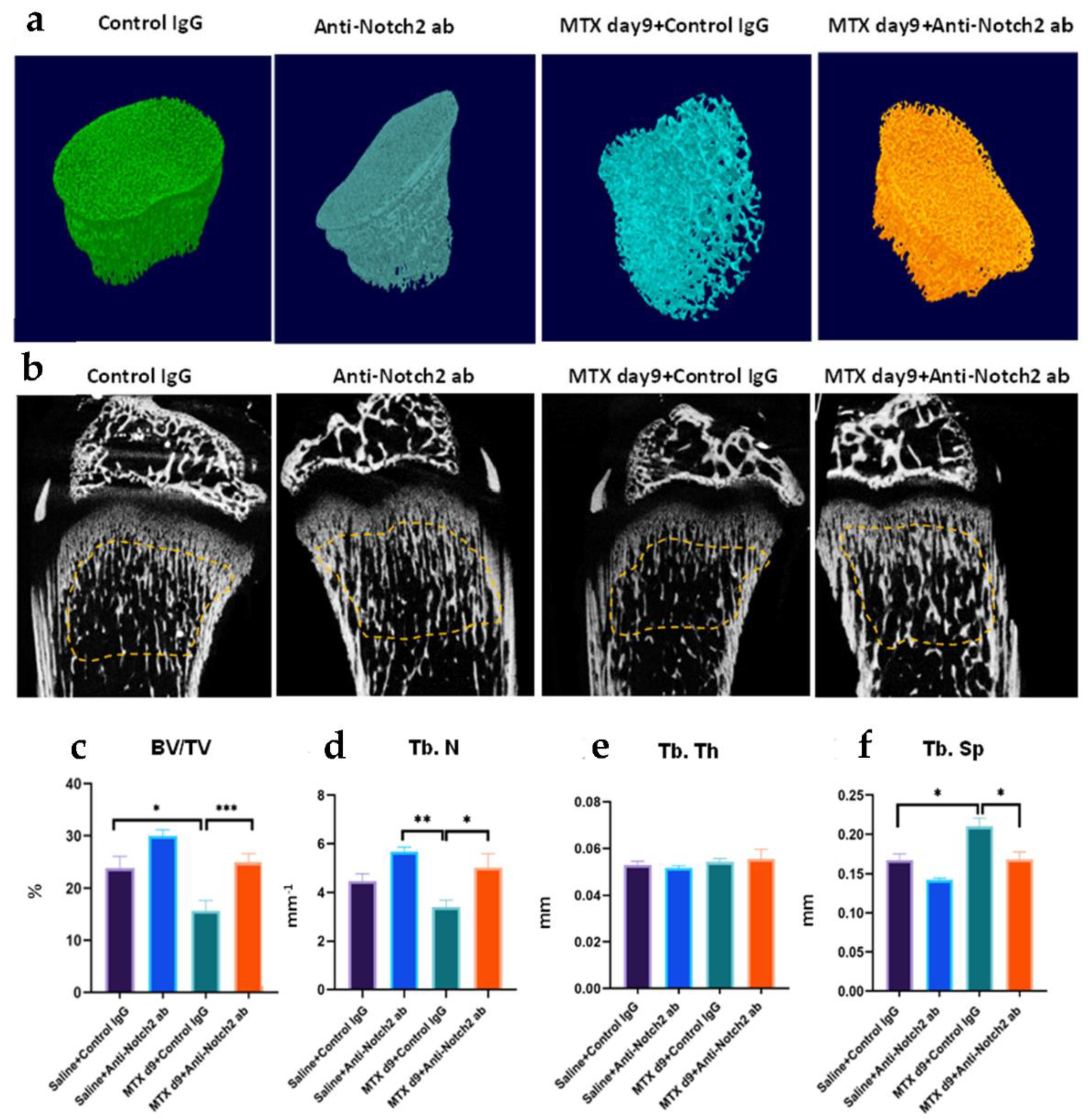
3.4. Notch2 Blockade Alleviated MTX Adverse Effects in Metaphysis Bone Volume and Structure
3.5. Notch2 Blockade Attenuated MTX Treatment-Induced Increases in Osteoclast Formation and Expression of Osteoclastogenesis-Related Genes
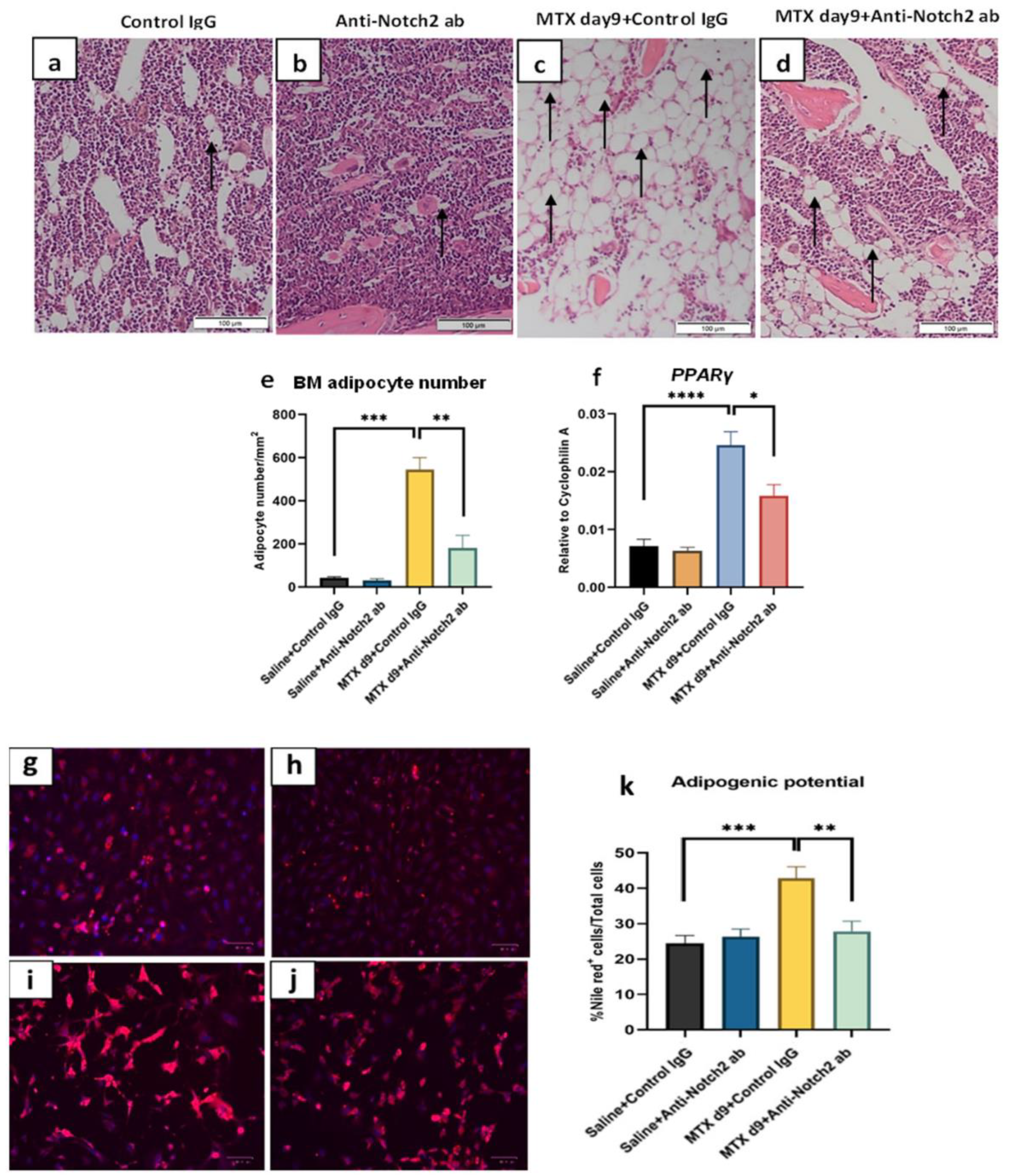
3.6. Notch2 Blockade Attenuated MTX Treatment-Induced Reciprocal Changes in Marrow Fat Volume
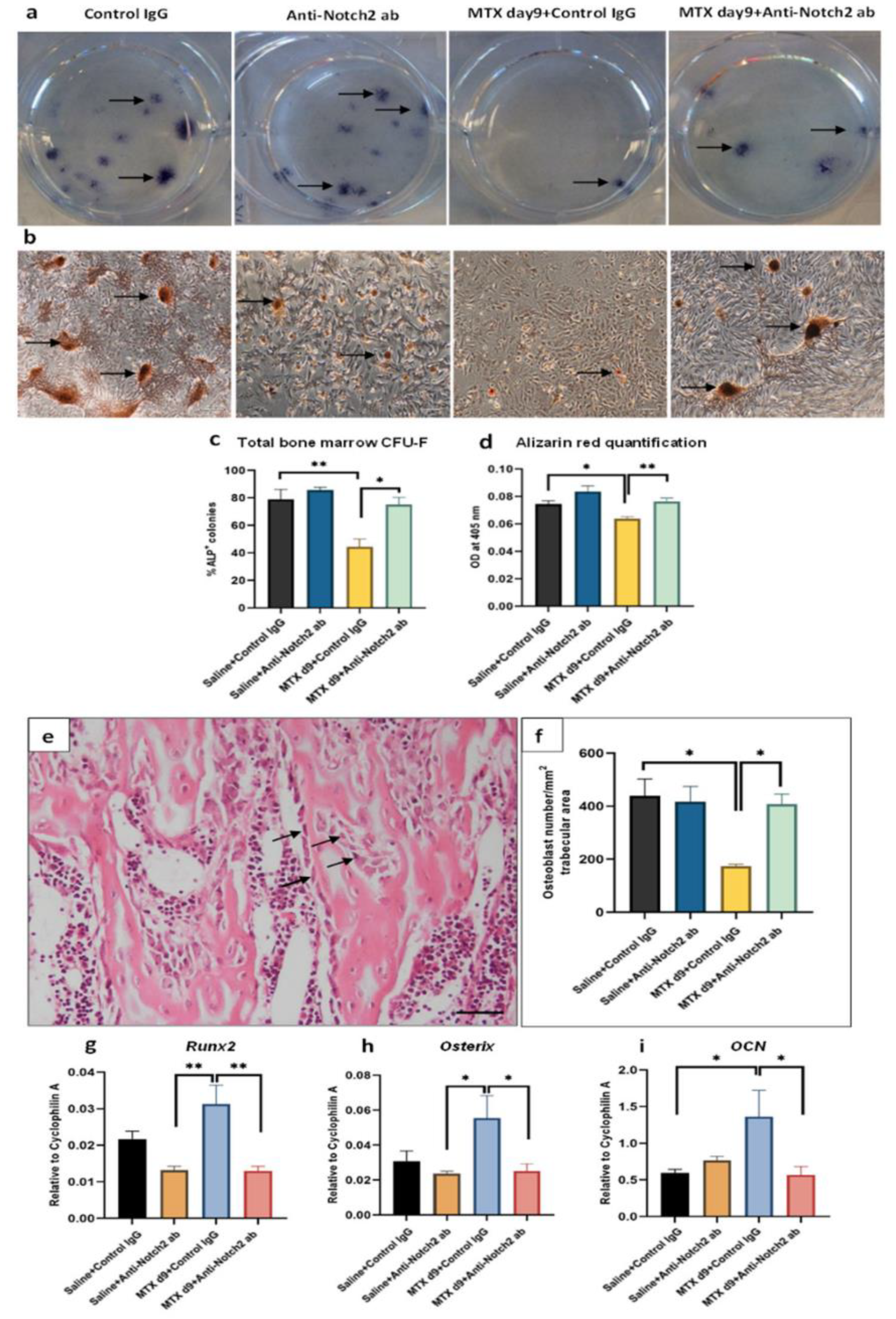
3.7. Notch2 Blockade Ameliorated MTX Treatment-Induced Increase in Adipogenesis and Reduced Osteogenesis in Isolated BMSCs
3.8. Treatment Effects on Osteoblast Density and Gene Expression of Key Regulatory Osteogenic Factors
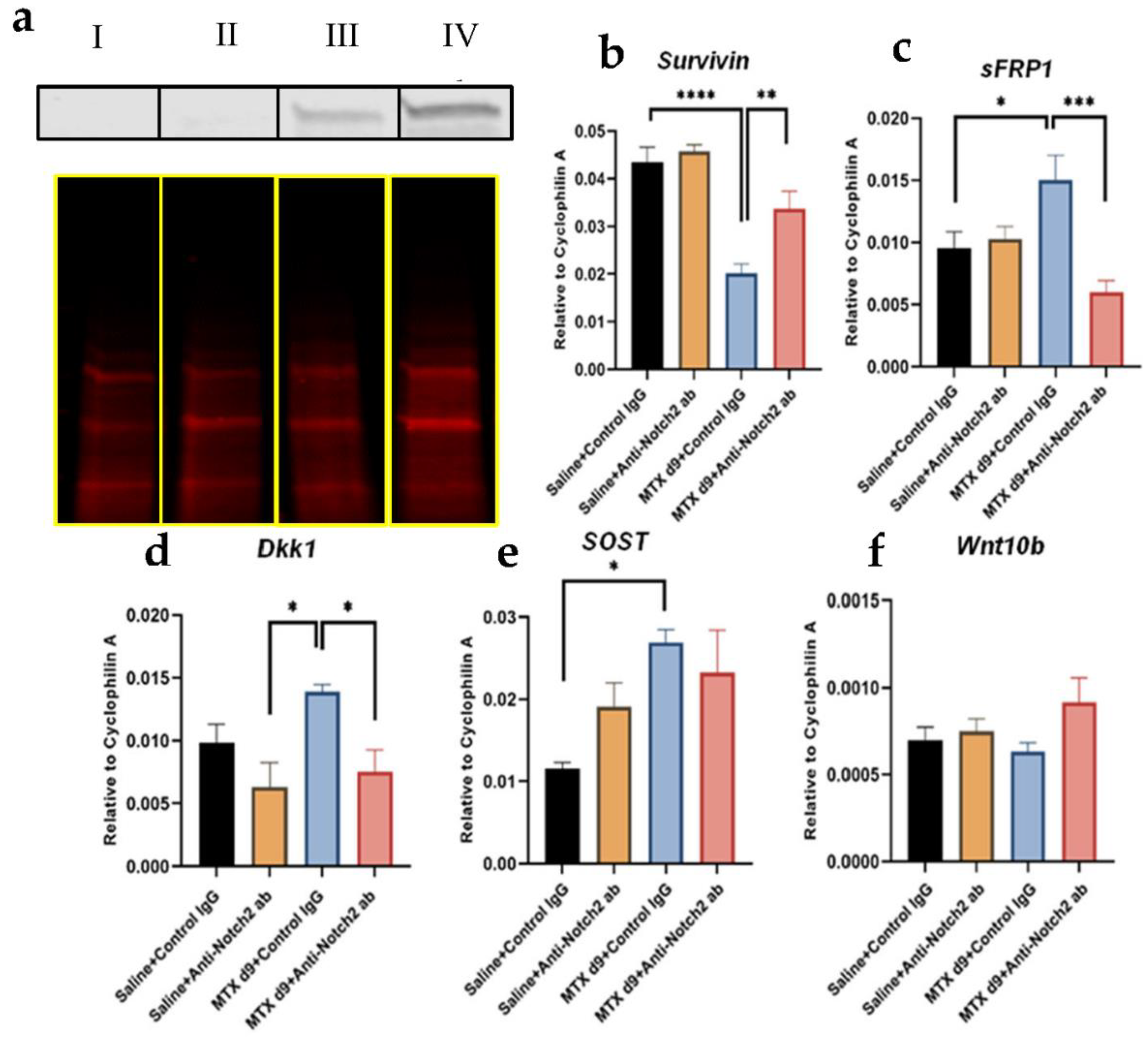
3.9. Notch2 Blockade in MTX-Treated Rats Was Associated with Changes in Activation of Wnt/β-Catenin Pathway and Expression of Key Wnt Signalling Mediators
4. Discussion
4.1. MTX Chemotherapy Bone Damage Is Associated with Alteration in Notch2 Signalling
4.2. Notch2 over Activation in Bone Is Associated with Imbalanced Bone Turnover Following MTX Chemotherapy
4.3. Over Activation of Notch2 in Bone Following MTX Treatment Is Associated with Osteogenesis/Adipogenesis Imbalance of BMSCs
4.4. Alteration in Notch2 Signalling Is Corelated with Changes in Wnt/β-Catenin Pathway Following MTX Treatment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kremer, J.M. Toward a better understanding of methotrexate. Arthritis Rheum. 2004, 50, 1370–1382. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.W.; Chen, K.M.; Hassanshahi, M.; Tang, Q.; Howe, P.R.; Xian, C.J. Childhood cancer chemotherapy-induced bone damage: Pathobiology and protective effects of resveratrol and other nutraceuticals. Ann. N. Y. Acad. Sci. 2017, 1403, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Meyers, P.A.; Schwartz, C.L.; Krailo, M.; Kleinerman, E.S.; Betcher, D.; Bernstein, M.L.; Conrad, E.; Ferguson, W.; Gebhardt, M.; Goorin, A.M.; et al. Osteosarcoma: A randomized, prospective trial of the addition of ifosfamide and/or muramyl tripeptide to cisplatin, doxorubicin, and high-dose methotrexate. J. Clin. Oncol. 2005, 23, 2004–2011. [Google Scholar] [CrossRef] [PubMed]
- Bonadonna, G.; Valagussa, P.; Moliterni, A.; Zambetti, M.; Brambilla, C. Adjuvant cyclophosphamide, methotrexate, and fluorouracil in node-positive breast cancer—The results of 20 years of follow-up. N. Engl. J. Med. 1995, 332, 901–906. [Google Scholar] [CrossRef] [PubMed]
- Jolivet, J.; Cowan, K.H.; Curt, G.A.; Clendeninn, N.J.; Chabner, B.A. The pharmacology and clinical use of methotrexate. N. Engl. J. Med. 1983, 309, 1094–1104. [Google Scholar] [CrossRef]
- Brody, J.P.; Krause, J.R.; Penchansky, L. Bone marrow response to chemotherapy in acute lymphocytic leukaemia and acute non-lymphocytic leukaemia. Scand. J. Haematol. 1985, 35, 240–245. [Google Scholar] [CrossRef]
- Gerard, E.L.; Ferry, J.A.; Amrein, P.C.; Harmon, D.C.; Mckinstry, R.C.; Hoppel, B.E.; Rosen, B.R. Compositional changes in vertebral bone marrow during treatment for acute leukemia: Assessment with quantitative chemical shift imaging. Radiology 1992, 183, 39–46. [Google Scholar] [CrossRef]
- Halton, J.M.; Atkinson, S.A.; Fraher, L.; Webber, C.; Gill, G.J.; Dawson, S.; Barr, R.D. Altered mineral metabolism and bone mass in children during treatment for acute lymphoblastic leukemia. J. Bone Min. Res. 1996, 11, 1774–1783. [Google Scholar] [CrossRef]
- Ahmed, S.F.; Wallace, W.H.; Kelnar, C.J. An anthropometric study of children during intensive chemotherapy for acute lymphoblastic leukaemia. Horm. Res. 1997, 48, 178–183. [Google Scholar] [CrossRef]
- Haddy, T.B.; Mosher, R.B.; Reaman, G.H. Osteoporosis in survivors of acute lymphoblastic leukemia. Oncologist 2001, 6, 278–285. [Google Scholar] [CrossRef] [Green Version]
- Kunstreich, M.; Kummer, S.; Laws, H.-J.; Borkhardt, A.; Kuhlen, M. Osteonecrosis in children with acute lymphoblastic leukemia. Haematologica 2016, 101, 1295–1305. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, B.; Kamps, W.; Jansen, H.; Hoekstra, H. The effect of chemotherapy on the growing skeleton. Cancer Treat. Rev. 2000, 26, 363–376. [Google Scholar] [CrossRef] [PubMed]
- Xian, C.J.; Cool, J.C.; Scherer, M.A.; Macsai, C.E.; Fan, C.; Covino, M.; Foster, B.K. Cellular mechanisms for methotrexate chemotherapy-induced bone growth defects. Bone 2007, 41, 842–850. [Google Scholar] [CrossRef] [PubMed]
- Xian, C.J.; Cool, J.C.; Scherer, M.A.; Fan, C.; Foster, B.K. Folinic acid attenuates methotrexate chemotherapy-induced damages on bone growth mechanisms and pools of bone marrow stromal cells. J. Cell Physiol. 2008, 214, 777–785. [Google Scholar] [CrossRef] [PubMed]
- Georgiou, K.R.; Hui, S.K.; Xian, C.J. Regulatory pathways associated with bone loss and bone marrow adiposity caused by aging, chemotherapy, glucocorticoid therapy and radiotherapy. Am. J. Stem Cells 2012, 1, 205–224. [Google Scholar]
- King, T.J.; Georgiou, K.R.; Cool, J.C.; Scherer, M.A.; Ang, E.S.M.; Foster, B.K.; Xu, J.; Xian, C.J. Methotrexate Chemotherapy Promotes Osteoclast Formation in the Long Bone of Rats via Increased Pro-Inflammatory Cytokines and Enhanced NF-κB Activation. Am. J. Pathol. 2012, 181, 121–129. [Google Scholar] [CrossRef]
- Chen, Q.; Shou, P.; Zheng, C.; Jiang, M.; Cao, G.; Yang, Q.; Cao, J.; Xie, N.; Velletri, T.; Zhang, X. Fate decision of mesenchymal stem cells: Adipocytes or osteoblasts? Cell Death Differ. 2016, 23, 1128–1139. [Google Scholar] [CrossRef] [Green Version]
- Fan, C.; Georgiou, K.R.; McKinnon, R.A.; Keefe, D.M.K.; Howe, P.R.C.; Xian, C.J. Combination chemotherapy with cyclophosphamide, epirubicin and 5-fluorouracil causes trabecular bone loss, bone marrow cell depletion and marrow adiposity in female rats. J. Bone Miner. Metab. 2016, 34, 277–290. [Google Scholar] [CrossRef]
- Georgiou, K.R.; King, T.J.; Scherer, M.A.; Zhou, H.; Foster, B.K.; Xian, C.J. Attenuated Wnt/β-catenin signalling mediates methotrexate chemotherapy-induced bone loss and marrow adiposity in rats. Bone 2012, 50, 1223–1233. [Google Scholar] [CrossRef]
- Steeve, K.T.; Marc, P.; Sandrine, T.; Dominique, H.; Yannick, F. IL-6, RANKL, TNF-alpha/IL-1: Interrelations in bone resorption pathophysiology. Cytokine Growth Factor Rev. 2004, 15, 49–60. [Google Scholar] [CrossRef]
- Gronthos, S.; Zannettino, A.C. The role of the chemokine CXCL12 in osteoclastogenesis. Trends Endocrinol. Metab. 2007, 18, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Raghu Nadhanan, R.; Fan, C.-M.; Su, Y.-W.; Howe, P.R.C.; Xian, C.J. Fish oil in comparison to folinic acid for protection against adverse effects of methotrexate chemotherapy on bone. J. Orthop. Re. 2014, 32, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Zanotti, S.; Canalis, E. Notch Signaling and the Skeleton. Endocr Rev. 2016, 37, 223–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindsell, C.E.; Boulter, J.; diSibio, G.; Gossler, A.; Weinmaster, G. Expression patterns of Jagged, Delta1, Notch1, Notch2, and Notch3 genes identify ligand-receptor pairs that may function in neural development. Mol. Cell Neurosci. 1996, 8, 14–27. [Google Scholar] [CrossRef]
- Baron, M. An overview of the Notch signalling pathway. Semin. Cell Dev. Biol. 2003, 14, 113–119. [Google Scholar] [CrossRef]
- Zanotti, S.; Canalis, E. Notch and its ligands. In Principles of Bone Biology; Elsevier: Amsterdam, The Netherlands, 2020; pp. 1083–1112. [Google Scholar]
- Bulman, M.P.; Kusumi, K.; Frayling, T.M.; McKeown, C.; Garrett, C.; Lander, E.S.; Krumlauf, R.; Hattersley, A.T.; Ellard, S.; Turnpenny, P.D. Mutations in the human delta homologue, DLL3, cause axial skeletal defects in spondylocostal dysostosis. Nat. Genet. 2000, 24, 438–441. [Google Scholar] [CrossRef]
- Sciaudone, M.; Gazzerro, E.; Priest, L.; Delany, A.M.; Canalis, E. Notch 1 impairs osteoblastic cell differentiation. Endocrinology 2003, 144, 5631–5639. [Google Scholar] [CrossRef] [Green Version]
- Deregowski, V.; Gazzerro, E.; Priest, L.; Rydziel, S.; Canalis, E. Notch 1 overexpression inhibits osteoblastogenesis by suppressing Wnt/β-catenin but not bone morphogenetic protein signaling. J. Biol. Chem. 2006, 281, 6203–6210. [Google Scholar] [CrossRef] [Green Version]
- Hilton, M.J.; Tu, X.; Wu, X.; Bai, S.; Zhao, H.; Kobayashi, T.; Kronenberg, H.M.; Teitelbaum, S.L.; Ross, F.P.; Kopan, R.; et al. Notch signaling maintains bone marrow mesenchymal progenitors by suppressing osteoblast differentiation. Nat. Med. 2008, 14, 306–314. [Google Scholar] [CrossRef] [Green Version]
- Canalis, E.; Schilling, L.; Yee, S.P.; Lee, S.K.; Zanotti, S. Hajdu Cheney Mouse Mutants Exhibit Osteopenia, Increased Osteoclastogenesis, and Bone Resorption. J. Biol. Chem. 2016, 291, 1538–1551. [Google Scholar] [CrossRef] [Green Version]
- Zanotti, S.; Yu, J.; Sanjay, A.; Schilling, L.; Schoenherr, C.; Economides, A.N.; Canalis, E. Sustained Notch2 signaling in osteoblasts, but not in osteoclasts, is linked to osteopenia in a mouse model of Hajdu-Cheney syndrome. J. Biol. Chem. 2017, 292, 12232–12244. [Google Scholar] [CrossRef] [Green Version]
- Tu, X.; Chen, J.; Lim, J.; Karner, C.M.; Lee, S.Y.; Heisig, J.; Wiese, C.; Surendran, K.; Kopan, R.; Gessler, M.; et al. Physiological notch signaling maintains bone homeostasis via RBPjk and Hey upstream of NFATc1. PLoS Genet. 2012, 8, e1002577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Axelrod, J.D.; Matsuno, K.; Artavanis-Tsakonas, S.; Perrimon, N. Interaction between Wingless and Notch signaling pathways mediated by dishevelled. Science 1996, 271, 1826–1832. [Google Scholar] [CrossRef] [PubMed]
- De Strooper, B.; Annaert, W. Where Notch and Wnt signaling meet. Presenilin Hub. J. Cell Biol. 2001, 152, F17–F20. [Google Scholar] [CrossRef] [Green Version]
- Lin, G.L.; Hankenson, K.D. Integration of BMP, Wnt, and notch signaling pathways in osteoblast differentiation. J. Cell Biochem. 2011, 112, 3491–3501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanotti, S.; Smerdel-Ramoya, A.; Stadmeyer, L.; Durant, D.; Radtke, F.; Canalis, E. Notch inhibits osteoblast differentiation and causes osteopenia. Endocrinology 2008, 149, 3890–3899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muguruma, Y.; Hozumi, K.; Warita, H.; Yahata, T.; Uno, T.; Ito, M.; Ando, K. Maintenance of Bone Homeostasis by DLL1-Mediated Notch Signaling. J. Cell Physiol. 2017, 232, 2569–2580. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Mabuchi, Y.; Miyamoto, K.; Araki, D.; Niibe, K.; Houlihan, D.D.; Morikawa, S.; Nakagawa, T.; Nakajima, T.; Akazawa, C.; et al. Notch2 Signaling Regulates the Proliferation of Murine Bone Marrow-Derived Mesenchymal Stem/Stromal Cells via c-Myc Expression. PLoS ONE 2016, 11, e0165946. [Google Scholar] [CrossRef]
- Lacey, D.L.; Timms, E.; Tan, H.L.; Kelley, M.J.; Dunstan, C.R.; Burgess, T.; Elliott, R.; Colombero, A.; Elliott, G.; Scully, S.; et al. Osteoprotegerin ligand is a cytokine that regulates osteoclast differentiation and activation. Cell 1998, 93, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Fukushima, H.; Nakao, A.; Okamoto, F.; Shin, M.; Kajiya, H.; Sakano, S.; Elliott, R.; Colombero, A.; Elliott, G.; Scully, S.; et al. The association of Notch2 and NF-kappaB accelerates RANKL-induced osteoclastogenesis. Mol. Cell. Biol. 2008, 28, 6402–6412. [Google Scholar] [CrossRef] [Green Version]
- Peymanfar, Y.; Su, Y.-W.; Hassanshahi, M.; Xian, C.J. Methotrexate treatment suppresses osteoblastic differentiation by inducing Notch2 signaling and blockade of Notch2 rescues osteogenesis by preserving Wnt/β-catenin signaling. J. Orthop. Res. 2021. [CrossRef] [PubMed]
- Friedlaender, G.E.; Tross, R.B.; Doganis, A.C.; Kirkwood, J.M.; Baron, R. Effects of chemotherapeutic agents on bone. I. Short-term methotrexate and doxorubicin (adriamycin) treatment in a rat model. J. Bone Jt. Surg. Am. 1984, 66, 602–607. [Google Scholar] [CrossRef]
- Wu, Y.; Cain-Hom, C.; Choy, L.; Hagenbeek, T.J.; de Leon, G.P.; Chen, Y.; Finkle, D.; Venook, R.; Wu, X.; Ridgway, J.; et al. Therapeutic antibody targeting of individual Notch receptors. Nature 2010, 464, 1052–1057. [Google Scholar] [CrossRef] [PubMed]
- Lafkas, D.; Shelton, A.; Chiu, C.; de Leon Boenig, G.; Chen, Y.; Stawicki, S.S.; Siltanen, C.; Reichelt, M.; Zhou, M.; Wu, X.; et al. Therapeutic antibodies reveal Notch control of transdifferentiation in the adult lung. Nature 2015, 528, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Canalis, E.; Sanjay, A.; Yu, J.; Zanotti, S. An Antibody to Notch2 Reverses the Osteopenic Phenotype of Hajdu-Cheney Mutant Male Mice. Endocrinology 2017, 158, 730–742. [Google Scholar] [CrossRef]
- Fan, C.; Cool, J.C.; Scherer, M.A.; Foster, B.K.; Shandala, T.; Tapp, H.; Xian, C.J. Damaging effects of chronic low-dose methotrexate usage on primary bone formation in young rats and potential protective effects of folinic acid supplementary treatment. Bone 2009, 44, 61–70. [Google Scholar] [CrossRef]
- Dishowitz, M.I.; Terkhorn, S.P.; Bostic, S.A.; Hankenson, K.D. Notch signaling components are upregulated during both endochondral and intramembranous bone regeneration. J. Orthop. Res. 2012, 30, 296–303. [Google Scholar] [CrossRef] [Green Version]
- Gregory, C.A.; Grady Gunn, W.; Peister, A.; Prockop, D.J. An Alizarin red-based assay of mineralization by adherent cells in culture: Comparison with cetylpyridinium chloride extraction. Anal. Biochem. 2004, 329, 77–84. [Google Scholar] [CrossRef]
- Ochotny, N.M. Role of the V-ATPase a3 Subunit in Osteoclast Maturation and Function. Doctoral Dissertation, University of Toronto, Toronto, ON, Canada, 2013. [Google Scholar]
- Georgiou, K.R.; Scherer, M.A.; Fan, C.M.; Cool, J.C.; King, T.J.; Foster, B.K.; Xian, C.J. Methotrexate chemotherapy reduces osteogenesis but increases adipogenic potential in the bone marrow. J. Cell Physiol. 2012, 227, 909–918. [Google Scholar] [CrossRef]
- King, T.J.; Shandala, T.; Lee, A.M.; Foster, B.K.; Chen, K.-M.; Howe, P.R.; Xian, C.J. Potential Effects of Phytoestrogen Genistein in Modulating Acute Methotrexate Chemotherapy-Induced Osteoclastogenesis and Bone Damage in Rats. Int. J. Mol. Sci. 2015, 16, 18293–18311. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Yu, S.; Myers, J.; Wang, Y.; Xin, W.W.; Albakri, M.; Xin, A.W.; Li, M.; Huang, A.Y.; Xin, W. Notch2 blockade enhances hematopoietic stem cell mobilization and homing. Haematologica 2017, 102, 1785–1795. [Google Scholar] [CrossRef] [Green Version]
- Canalis, E.; Adams, D.J.; Boskey, A.; Parker, K.; Kranz, L.; Zanotti, S. Notch signaling in osteocytes differentially regulates cancellous and cortical bone remodeling. J. Biol. Chem. 2013, 288, 25614–25625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Li, Y.; Ahmad, A.; Azmi, A.S.; Banerjee, S.; Kong, D.; Sarkar, F.H. Targeting Notch signaling pathway to overcome drug resistance for cancer therapy. Biochim. Biophys. Acta 2010, 1806, 258–267. [Google Scholar]
- Bellavia, D.; Palermo, R.; Felli, M.P.; Screpanti, I.; Checquolo, S. Notch signaling as a therapeutic target for acute lymphoblastic leukemia. Expert Opin. Ther. Targets 2018, 22, 331–342. [Google Scholar] [CrossRef]
- Lee, A.M.C.; Shandala, T.; Soo, P.P.; Su, Y.-W.; King, T.J.; Chen, K.-M.; Sarkar, F.H. Effects of Resveratrol Supplementation on Methotrexate Chemotherapy-Induced Bone Loss. Nutrients 2017, 9, 255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Fujiwara, T.; Ye, S.; Li, X.; Zhao, H. Ubiquitin E3 Ligase LNX2 is Critical for Osteoclastogenesis In Vitro by Regulating M-CSF/RANKL Signaling and Notch2. Calcif. Tissue Int. 2015, 96, 465–475. [Google Scholar] [CrossRef] [Green Version]
- Jin, W.J.; Kim, B.; Kim, J.-W.; Kim, H.-H.; Ha, H.; Lee, Z.H. Notch2 signaling promotes osteoclast resorption via activation of PYK2. Cell. Signal. 2016, 28, 357–365. [Google Scholar] [CrossRef]
- Song, B.; Chi, Y.; Li, X.; Du, W.; Han, Z.B.; Tian, J.; Li, J.-J.; Chen, F.; Wu, H.; Han, L.-X.; et al. Inhibition of Notch Signaling Promotes the Adipogenic Differentiation of Mesenchymal Stem Cells Through Autophagy Activation and PTEN-PI3K/AKT/mTOR Pathway. Cell. Physiol. Biochem. 2015, 36, 1991–2002. [Google Scholar] [CrossRef]
- Engin, F.; Yao, Z.; Yang, T.; Zhou, G.; Bertin, T.; Jiang, M.M.; Chen, Y.; Wang, L.; Zheng, H.; Sutton, R.E.; et al. Dimorphic effects of Notch signaling in bone homeostasis. Nat. Med. 2008, 14, 299–305. [Google Scholar] [CrossRef]
- Canalis, E.; Bridgewater, D.; Schilling, L.; Zanotti, S. Canonical Notch activation in osteocytes causes osteopetrosis. Am. J. Physiol.-Endocrinol. Metab. 2016, 310, E171–E182. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.; Wu, H.; Xu, H.; Xiong, H.; Chu, Q.; Yu, S.; Wu, G.S.; Wu, K. Notch signaling: An emerging therapeutic target for cancer treatment. Cancer Lett. 2015, 369, 20–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takebe, N.; Miele, L.; Harris, P.J.; Jeong, W.; Bando, H.; Kahn, M.; Yang, S.X.; Ivy, S.P. Targeting Notch, Hedgehog, and Wnt pathways in cancer stem cells: Clinical update. Nat. Rev. Clin. Oncol. 2015, 12, 445–464. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Genes | Forward Sequence (5′-3′) | Reverse Sequence (5′-3′) |
|---|---|---|---|
| Cyclophilin A | PPIA | GAGCTGTTTGCAGACAAAGTTC | CCCTGGCACATGAATCCTG |
| Dkk-1 | DKK1 | GGTTCTTGGTCGTGCTTTCA | CTTGATCGCGTTGGAATTGA |
| Nfatc1 | NFATC1 | GTGCAAGCCAAATTCCCTGG | CTTGGACGGGGCTGGTTATT |
| RANKL | TNFSF11 | CCGTGCAAAGGGAATTACAAC | GAGCCACGAACCTTCCATCA |
| OPG | TNFRSF11B | CACAGCTCGCAAGAGCAAACT | ATATGCCGTTGCACACTGCTT |
| Runx2 | RUNX2 | TCACAAATCCTCCCCAAGTGG | GAATGCGCCCTAAATCACTGA |
| Osteocalcin | BGLAP | GCTGGCCCTGACTGCATTCTG | ATTCACCACCTTACTGCCCTCCTG |
| Osterix | SP7 | GCTTTTCTGTGGCAAGAGGTTC | CTGATGTTTGCTCAAGTGGTCG |
| SOST | SOST | CAACCAGACCATGAACCGGG | AAGCGGGTGTAGTGCAGCTC |
| PPARγ | PPARG | AACGTGAAGCCCATCGAGGACATC | CTTGGCGAACAGCTGGGAGGAC |
| sFRP-1 | SFRP1 | CCCGAGATGCTCAAATGTGAC | AGATGTTCGATGATGGCCTCC |
| β-Catenin | CTNB1 | CTTGGCTGAACCGTCACAGAT | TCCTCGTCATTTAGCAGTTTGG |
| Survivin | BIRC5 | AACTGGCCCTTCCTGGAG | TCAGGCTCGTTCTCGGTAG |
| Wnt10b | WNT10B | AGAATGCGGATCCACAACAAC | TCCAACAGGTCTTGAATTGGC |
| Notch 1 | NOTCH1 | CCAGGGTGGTCAGGAAAGTC | GGTTCTGGCTGCACTCGTTA |
| Notch2 | NOTCH2 | ATGCCGGGTTTCAAAGGTGT | ATGTCGATCTGGCACACTGG |
| Hes1 | HES1 | GACACCGGACAAACCAAAGAC | AATGCCGGGAGCTATCTTTCT |
| HeyL | HEYL | GTCCCCACTGCCTTTGAGAA | CATCAAAGAACCCTGCGCCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peymanfar, Y.; Su, Y.-W.; Xian, C.J. Notch2 Blockade Mitigates Methotrexate Chemotherapy-Induced Bone Loss and Marrow Adiposity. Cells 2022, 11, 1521. https://doi.org/10.3390/cells11091521
Peymanfar Y, Su Y-W, Xian CJ. Notch2 Blockade Mitigates Methotrexate Chemotherapy-Induced Bone Loss and Marrow Adiposity. Cells. 2022; 11(9):1521. https://doi.org/10.3390/cells11091521
Chicago/Turabian StylePeymanfar, Yaser, Yu-Wen Su, and Cory J. Xian. 2022. "Notch2 Blockade Mitigates Methotrexate Chemotherapy-Induced Bone Loss and Marrow Adiposity" Cells 11, no. 9: 1521. https://doi.org/10.3390/cells11091521
APA StylePeymanfar, Y., Su, Y.-W., & Xian, C. J. (2022). Notch2 Blockade Mitigates Methotrexate Chemotherapy-Induced Bone Loss and Marrow Adiposity. Cells, 11(9), 1521. https://doi.org/10.3390/cells11091521