
Glycogene Expression Profile of Human Limbal Epithelial Cells with Distinct Clonogenic Potential



{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth-Arrested 3T3-J2 Fibroblasts
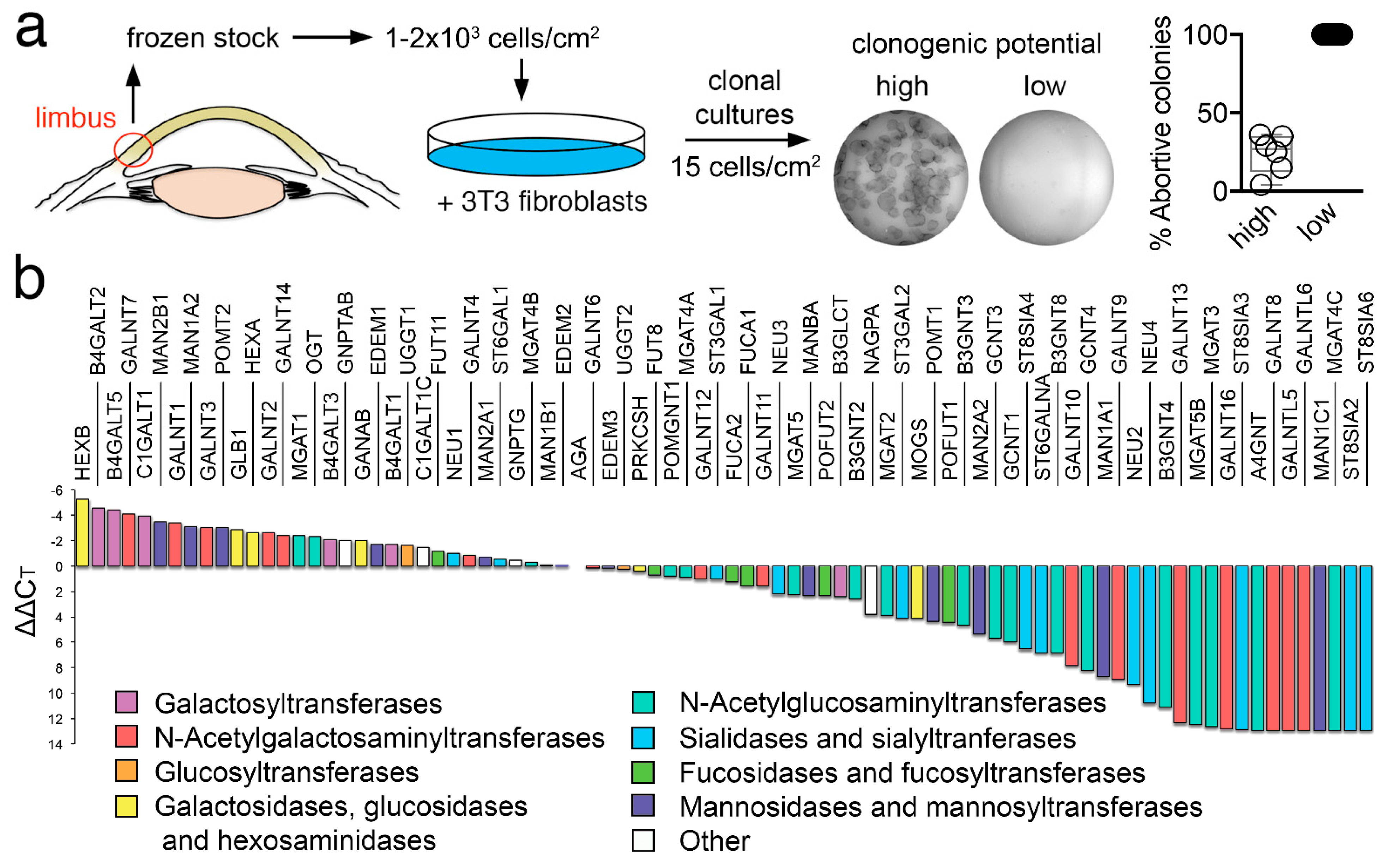
2.2. Human Limbal Epithelial Cell Culture
2.3. Isolation of Individual Clones
2.4. Clonal Classification
2.5. RNA Isolation and cDNA Synthesis
2.6. Human Glycosylation PCR Array
2.7. qPCR
2.8. Statistical Analyses
3. Results and Discussion
3.1. Glycogene Expression Profile in Human Limbal Epithelial Cells
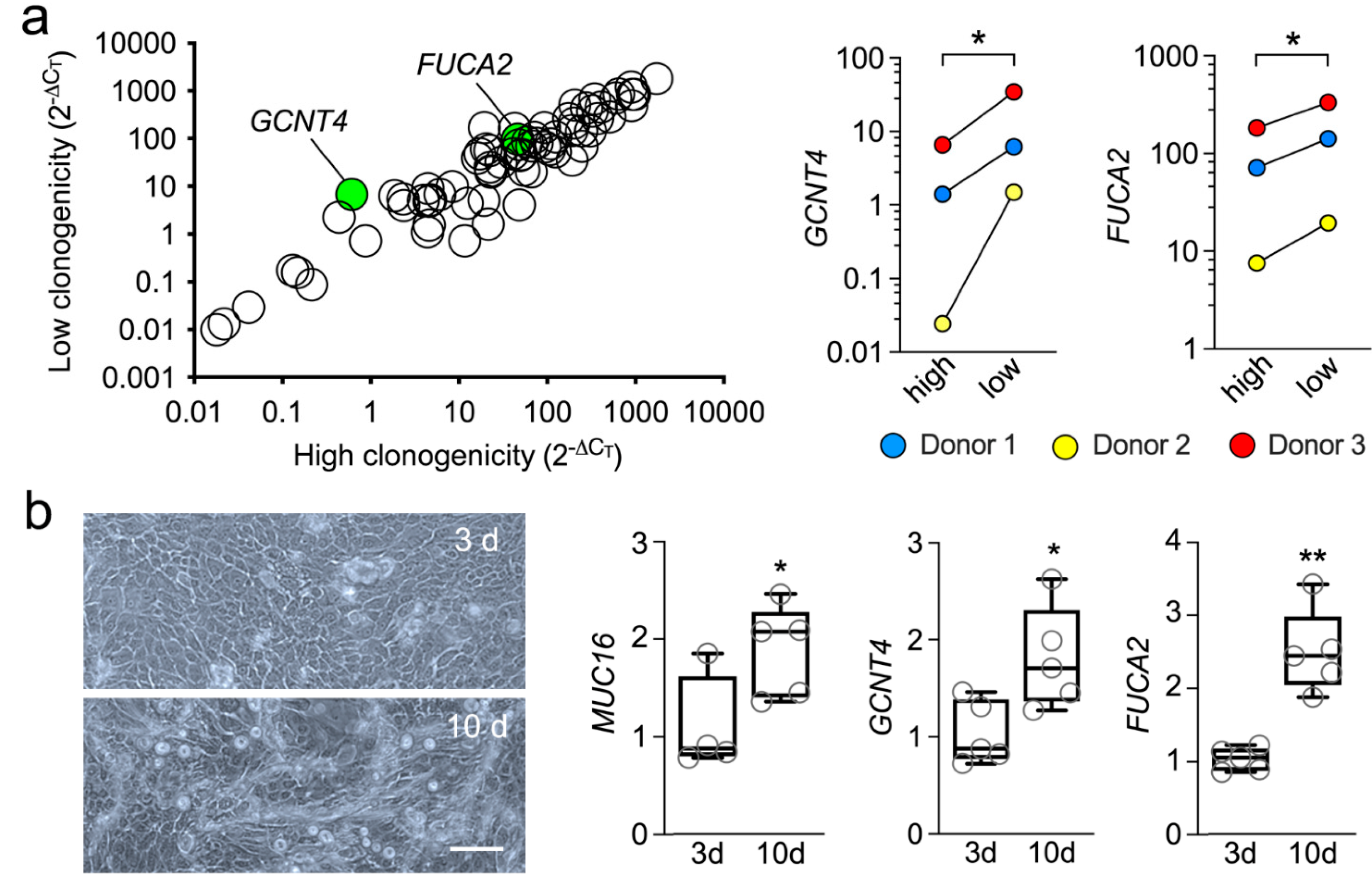
3.2. Abortive Clones Contain High Levels of GCNT4 and FUCA2
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Comelli, E.M.; Head, S.R.; Gilmartin, T.; Whisenant, T.; Haslam, S.M.; North, S.J.; Wong, N.K.; Kudo, T.; Narimatsu, H.; Esko, J.D.; et al. A focused microarray approach to functional glycomics: Transcriptional regulation of the glycome. Glycobiology 2006, 16, 117–131. [Google Scholar] [CrossRef]
- Pothukuchi, P.; Agliarulo, I.; Russo, D.; Rizzo, R.; Russo, F.; Parashuraman, S. Translation of genome to glycome: Role of the Golgi apparatus. FEBS Lett. 2019, 593, 2390–2411. [Google Scholar] [CrossRef] [Green Version]
- Freeze, H.H.; Chong, J.X.; Bamshad, M.J.; Ng, B.G. Solving glycosylation disorders: Fundamental approaches reveal complicated pathways. Am. J. Hum. Genet. 2014, 94, 161–175. [Google Scholar] [CrossRef] [Green Version]
- Handa, K.; Hakomori, S.I. Changes of glycoconjugate expression profiles during early development. Glycoconj. J. 2017, 34, 693–699. [Google Scholar] [CrossRef]
- Breimer, M.E.; Saljo, K.; Barone, A.; Teneberg, S. Glycosphingolipids of human embryonic stem cells. Glycoconj. J. 2017, 34, 713–723. [Google Scholar] [CrossRef] [Green Version]
- Mikami, T.; Kitagawa, H. Sulfated glycosaminoglycans: Their distinct roles in stem cell biology. Glycoconj. J. 2017, 34, 725–735. [Google Scholar] [CrossRef]
- Furukawa, J.I.; Okada, K.; Shinohara, Y. Glycomics of human embryonic stem cells and human induced pluripotent stem cells. Glycoconj. J. 2017, 34, 807–815. [Google Scholar] [CrossRef]
- Satomaa, T.; Heiskanen, A.; Mikkola, M.; Olsson, C.; Blomqvist, M.; Tiittanen, M.; Jaatinen, T.; Aitio, O.; Olonen, A.; Helin, J.; et al. The N-glycome of human embryonic stem cells. BMC Cell Biol. 2009, 10, 42. [Google Scholar] [CrossRef] [Green Version]
- Hasehira, K.; Tateno, H.; Onuma, Y.; Ito, Y.; Asashima, M.; Hirabayashi, J. Structural and quantitative evidence for dynamic glycome shift on production of induced pluripotent stem cells. Mol. Cell Proteom. 2012, 11, 1913–1923. [Google Scholar] [CrossRef] [Green Version]
- Lanctot, P.M.; Gage, F.H.; Varki, A.P. The glycans of stem cells. Curr. Opin. Chem. Biol. 2007, 11, 373–380. [Google Scholar] [CrossRef]
- Gurusamy, N.; Alsayari, A.; Rajasingh, S.; Rajasingh, J. Adult Stem Cells for Regenerative Therapy. Prog. Mol. Biol. Transl. Sci. 2018, 160, 1–22. [Google Scholar] [CrossRef]
- Cotsarelis, G.; Cheng, S.Z.; Dong, G.; Sun, T.T.; Lavker, R.M. Existence of slow-cycling limbal epithelial basal cells that can be preferentially stimulated to proliferate: Implications on epithelial stem cells. Cell 1989, 57, 201–209. [Google Scholar] [CrossRef]
- Lindberg, K.; Brown, M.E.; Chaves, H.V.; Kenyon, K.R.; Rheinwald, J.G. In vitro propagation of human ocular surface epithelial cells for transplantation. Investig. Ophthalmol. Vis. Sci. 1993, 34, 2672–2679. [Google Scholar]
- Barrandon, Y.; Green, H. Three clonal types of keratinocyte with different capacities for multiplication. Proc. Natl. Acad. Sci. USA 1987, 84, 2302–2306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellegrini, G.; Golisano, O.; Paterna, P.; Lambiase, A.; Bonini, S.; Rama, P.; De Luca, M. Location and clonal analysis of stem cells and their differentiated progeny in the human ocular surface. J. Cell Biol. 1999, 145, 769–782. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Li, M.; Qi, X.; Li, J. β1,4-Galactosyltransferase V Modulates Breast Cancer Stem Cells through Wnt/beta-catenin Signaling Pathway. Cancer Res. Treat. 2020, 52, 1084–1102. [Google Scholar] [CrossRef] [PubMed]
- Pecori, F.; Akimoto, Y.; Hanamatsu, H.; Furukawa, J.I.; Shinohara, Y.; Ikehara, Y.; Nishihara, S. Mucin-type O-glycosylation controls pluripotency in mouse embryonic stem cells via Wnt receptor endocytosis. J. Cell Sci. 2020, 133, jcs245845. [Google Scholar] [CrossRef]
- Argueso, P.; Spurr-Michaud, S.; Russo, C.L.; Tisdale, A.; Gipson, I.K. MUC16 mucin is expressed by the human ocular surface epithelia and carries the H185 carbohydrate epitope. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2487–2495. [Google Scholar] [CrossRef] [Green Version]
- Brockhausen, I.; Elimova, E.; Woodward, A.M.; Argueso, P. Glycosylation pathways of human corneal and conjunctival epithelial cell mucins. Carbohydr. Res. 2018, 470, 50–56. [Google Scholar] [CrossRef]
- Nairn, A.V.; Aoki, K.; dela Rosa, M.; Porterfield, M.; Lim, J.M.; Kulik, M.; Pierce, J.M.; Wells, L.; Dalton, S.; Tiemeyer, M.; et al. Regulation of glycan structures in murine embryonic stem cells: Combined transcript profiling of glycan-related genes and glycan structural analysis. J. Biol. Chem. 2012, 287, 37835–37856. [Google Scholar] [CrossRef] [Green Version]
- Woodward, A.M.; Lehoux, S.; Mantelli, F.; Di Zazzo, A.; Brockhausen, I.; Bonini, S.; Argueso, P. Inflammatory Stress Causes N-Glycan Processing Deficiency in Ocular Autoimmune Disease. Am. J. Pathol. 2019, 189, 283–294. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, T.; Woodward, A.M.; Magnelli, P.; McColgan, N.M.; Lehoux, S.; Jacobo, S.M.P.; Mauris, J.; Argueso, P. N-Glycosylation affects the stability and barrier function of the MUC16 mucin. J. Biol. Chem. 2017, 292, 11079–11090. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Dong, H.T.; Zhao, T.; Yao, F.; Xu, Y.; Chen, B.; Wu, Y.; Jin, F.; Xing, P. Cancer-associated adipocytes release FUCA2 to promote aggressiveness in TNBC. Endocr. Relat. Cancer 2022, 29, 139–149. [Google Scholar] [CrossRef]
- Rodriguez Benavente, M.C.; Argueso, P. Glycosylation pathways at the ocular surface. Biochem. Soc. Trans. 2018, 46, 343–350. [Google Scholar] [CrossRef]
- Cao, Z.; Zhao, Z.; Mohan, R.; Alroy, J.; Stanley, P.; Panjwani, N. Role of the Lewis(x) glycan determinant in corneal epithelial cell adhesion and differentiation. J. Biol. Chem. 2001, 276, 21714–21723. [Google Scholar] [CrossRef] [Green Version]
- Gesteira, T.F.; Sun, M.; Coulson-Thomas, Y.M.; Yamaguchi, Y.; Yeh, L.K.; Hascall, V.; Coulson-Thomas, V.J. Hyaluronan Rich Microenvironment in the Limbal Stem Cell Niche Regulates Limbal Stem Cell Differentiation. Investig. Ophthalmol. Vis. Sci. 2017, 58, 4407–4421. [Google Scholar] [CrossRef] [Green Version]
- Yazdani, M.; Shahdadfar, A.; Jackson, C.J.; Utheim, T.P. Hyaluronan-Based Hydrogel Scaffolds for Limbal Stem Cell Transplantation: A Review. Cells 2019, 8, 245. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guindolet, D.; Woodward, A.M.; Gabison, E.E.; Argüeso, P. Glycogene Expression Profile of Human Limbal Epithelial Cells with Distinct Clonogenic Potential. Cells 2022, 11, 1575. https://doi.org/10.3390/cells11091575
Guindolet D, Woodward AM, Gabison EE, Argüeso P. Glycogene Expression Profile of Human Limbal Epithelial Cells with Distinct Clonogenic Potential. Cells. 2022; 11(9):1575. https://doi.org/10.3390/cells11091575
Chicago/Turabian StyleGuindolet, Damien, Ashley M. Woodward, Eric E. Gabison, and Pablo Argüeso. 2022. "Glycogene Expression Profile of Human Limbal Epithelial Cells with Distinct Clonogenic Potential" Cells 11, no. 9: 1575. https://doi.org/10.3390/cells11091575
APA StyleGuindolet, D., Woodward, A. M., Gabison, E. E., & Argüeso, P. (2022). Glycogene Expression Profile of Human Limbal Epithelial Cells with Distinct Clonogenic Potential. Cells, 11(9), 1575. https://doi.org/10.3390/cells11091575