
Identification of R-Spondin Gene Signature Predictive of Metastatic Progression in BRAFV600E-Positive Papillary Thyroid Cancer

 ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Sample Preparation and RNA Isolation
2.3. RNA Library Construction and Sequencing
2.4. Bioinformatic Analysis
2.5. Cell Culture
2.6. CRISPR-Cas9 for the Generation of RSPO4 and BRAF Knockout Cells
2.7. qRT-PCR
2.8. Western Blot Analysis
2.9. Cell Proliferation
2.10. Cell Motility and Invasion Assays
2.11. Immunohistochemistry (IHC) in the PTC Tissue Microarray (TMA)
2.12. Statistical Analysis
3. Results
3.1. Patient Description
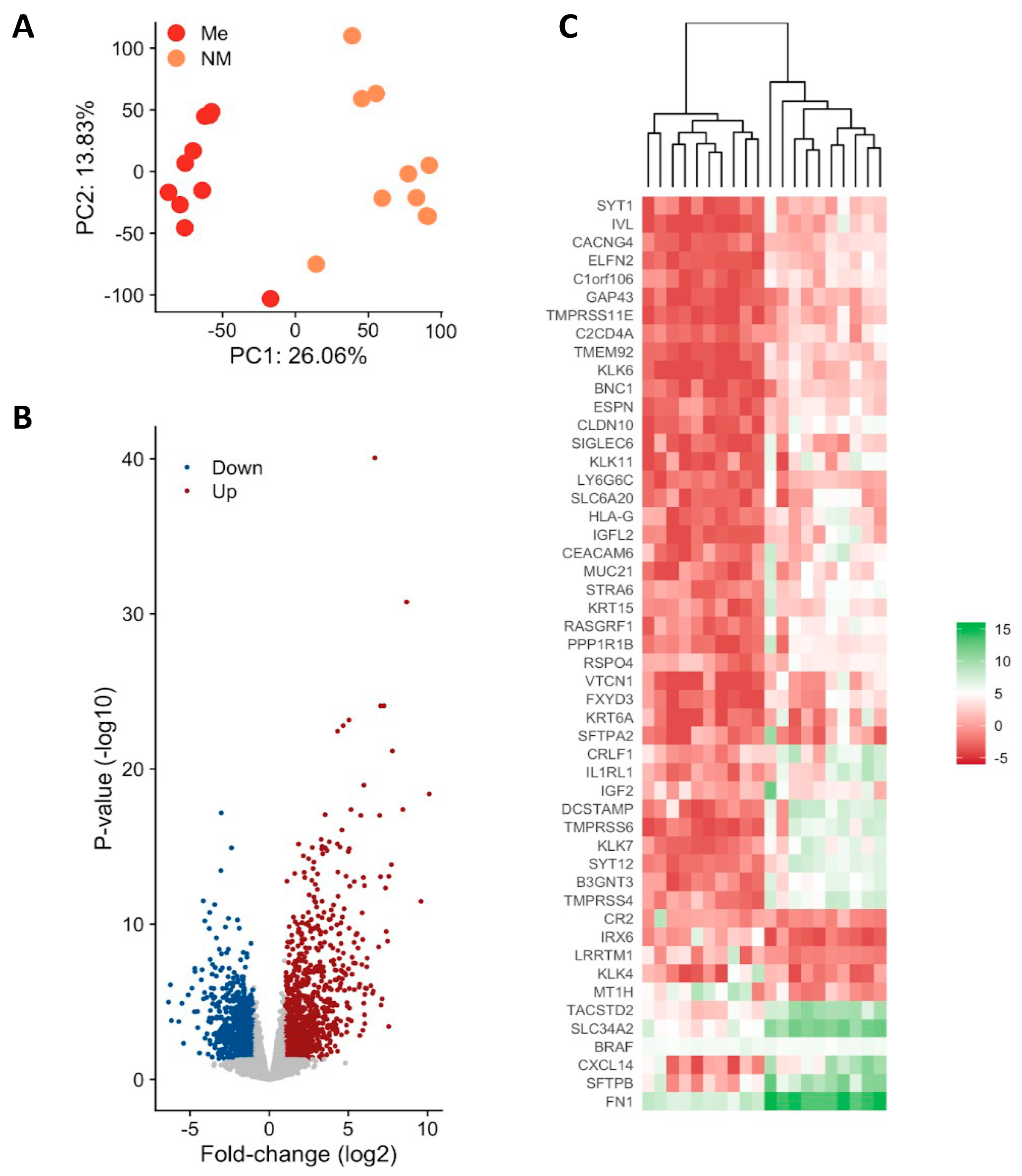
3.2. Gene Expression Profiling of Metastatic vs. Non-Metastatic PTC
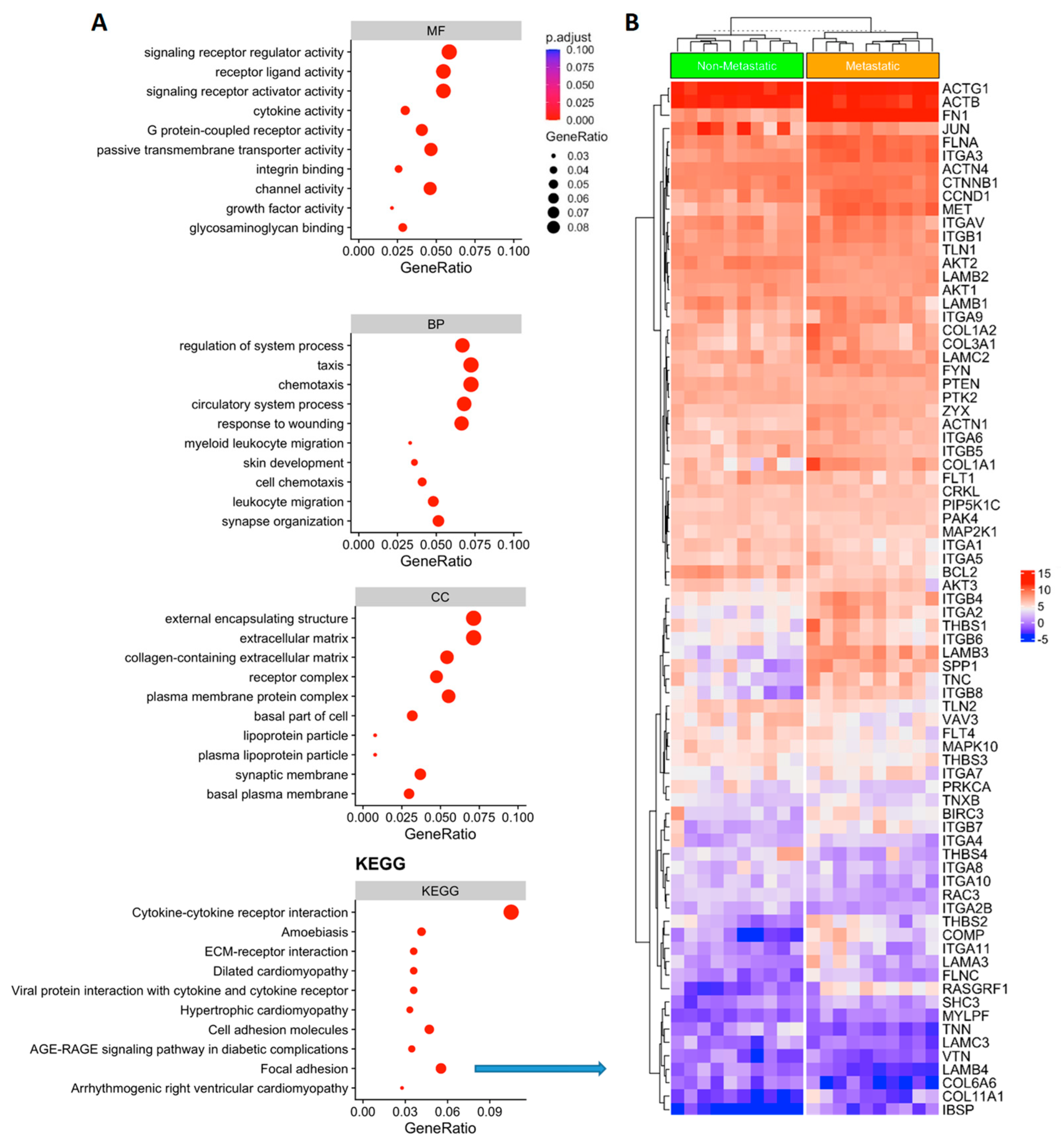
3.3. Functional Analysis of Genes Associated with Metastatic PTC
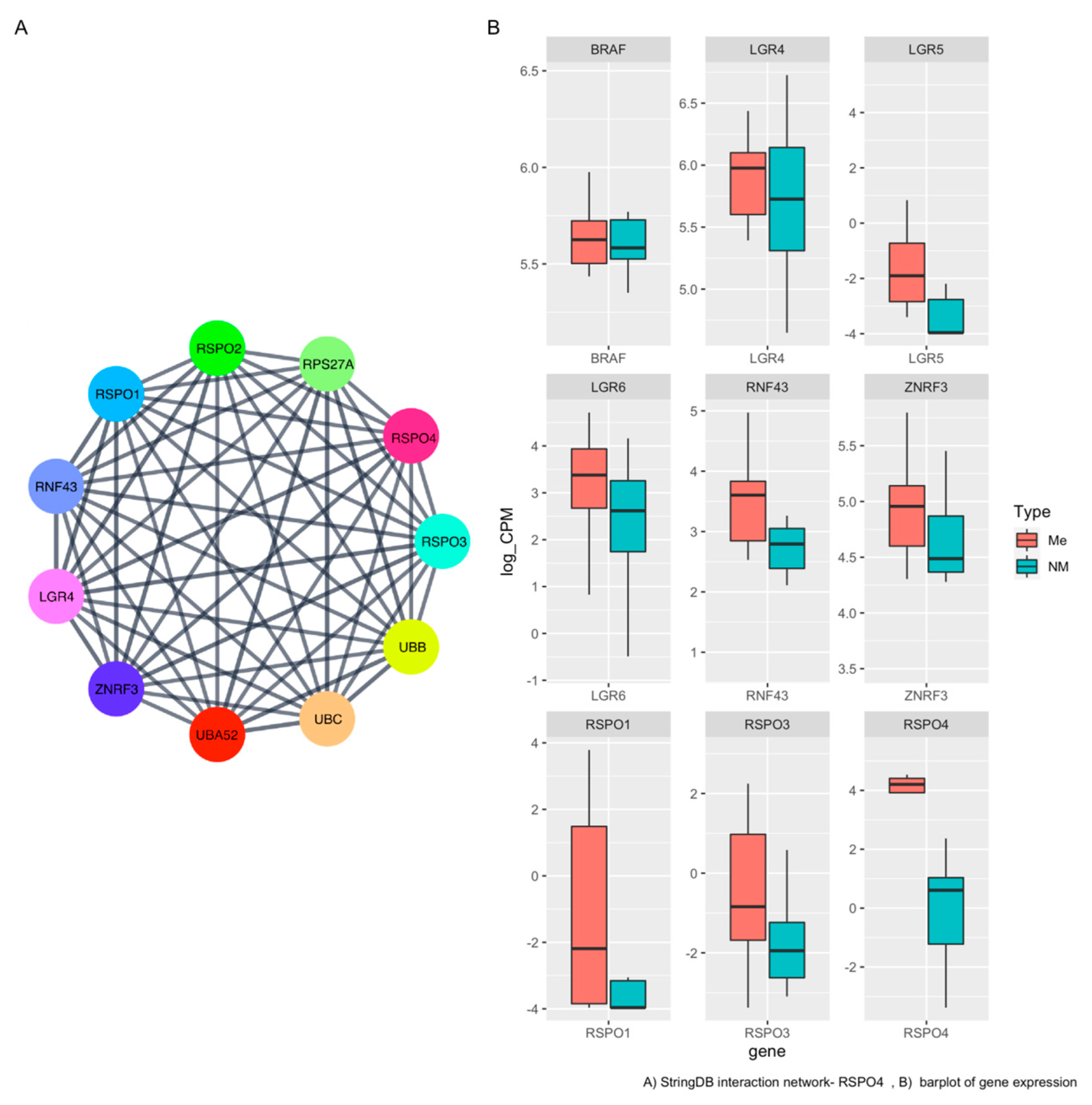
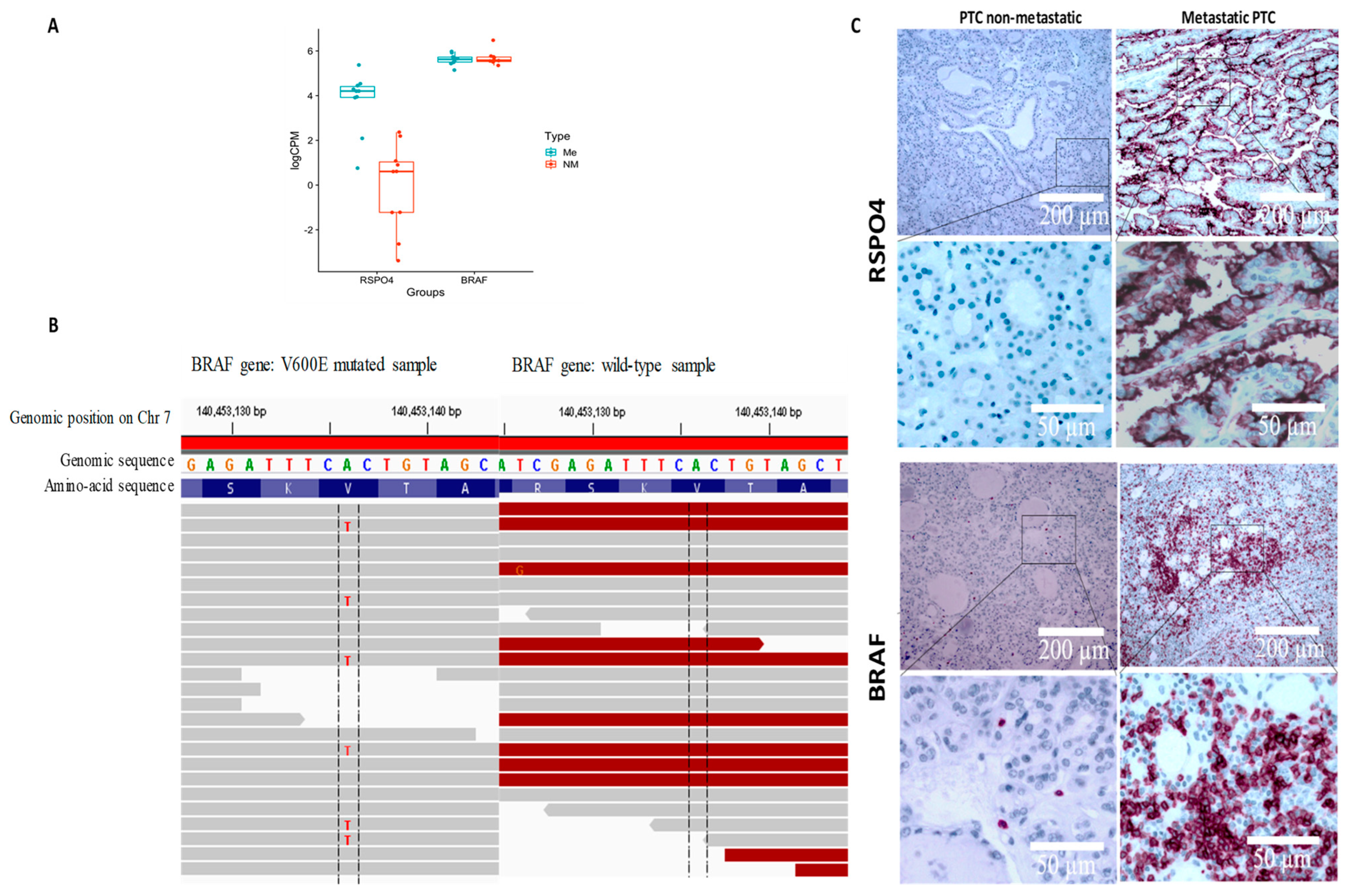
3.4. RSPO4 Is the Most Significantly Overexpressed Gene in Metastatic BRAF V600E PTC
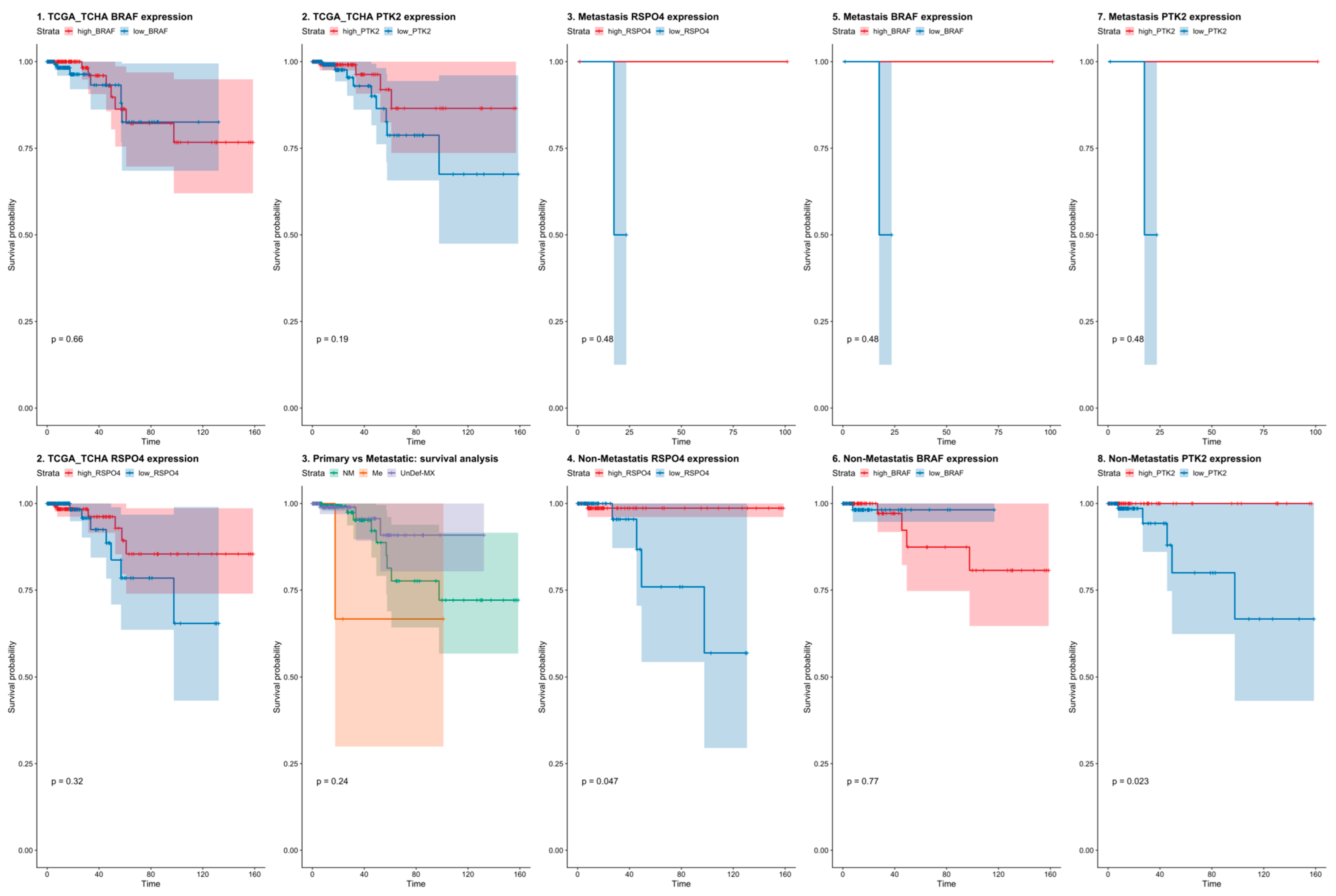
3.5. RSPO4 Can Serve as a Robust Biomarker of Metastatic PTC
3.6. RSPO4 Overexpression Occurs Selectively in BRAF V600E PTC
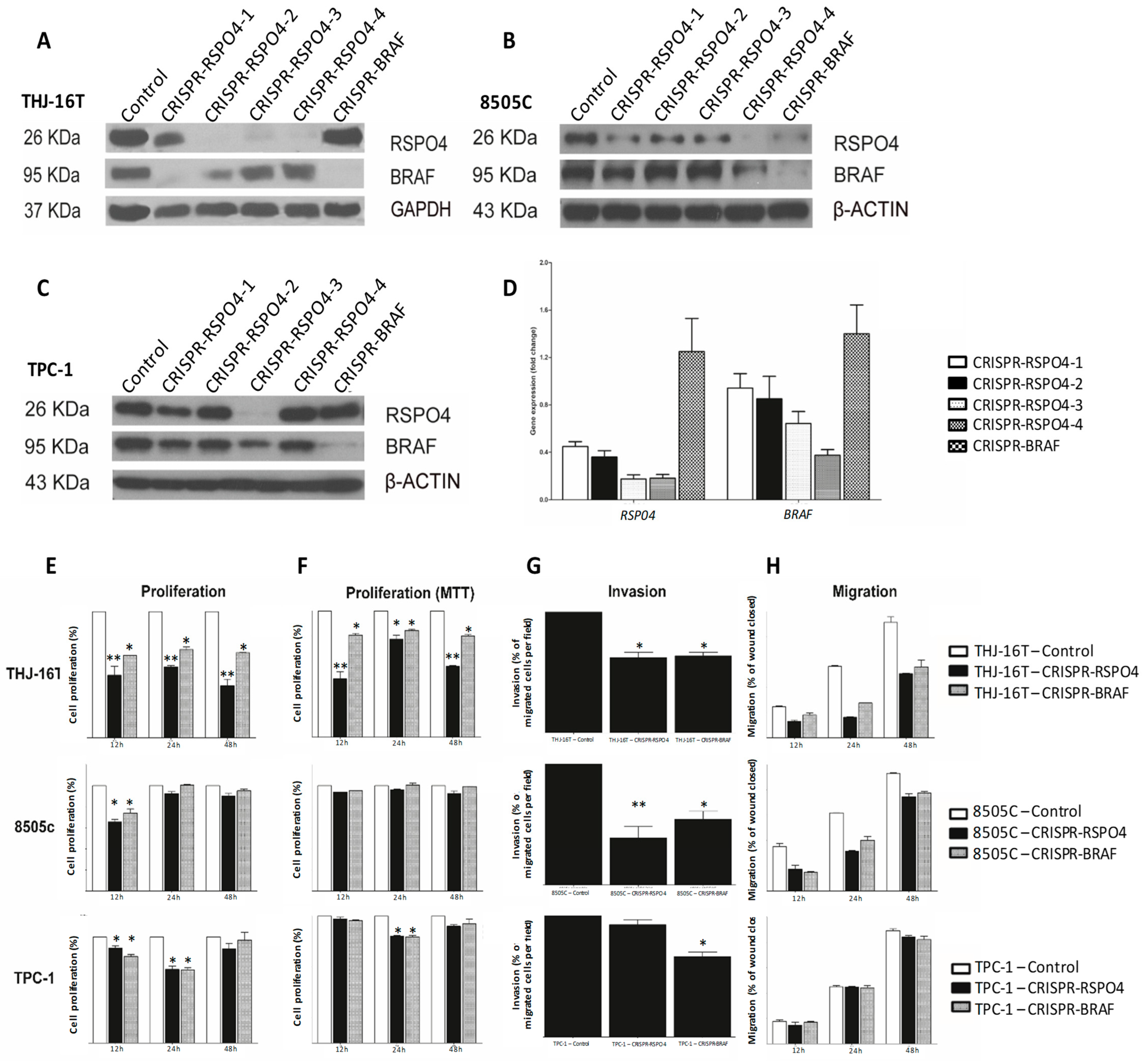
3.7. BRAF Inactivation and RSPO4 Overexpression Influence PTC Cell Proliferation, Motility and Invasion
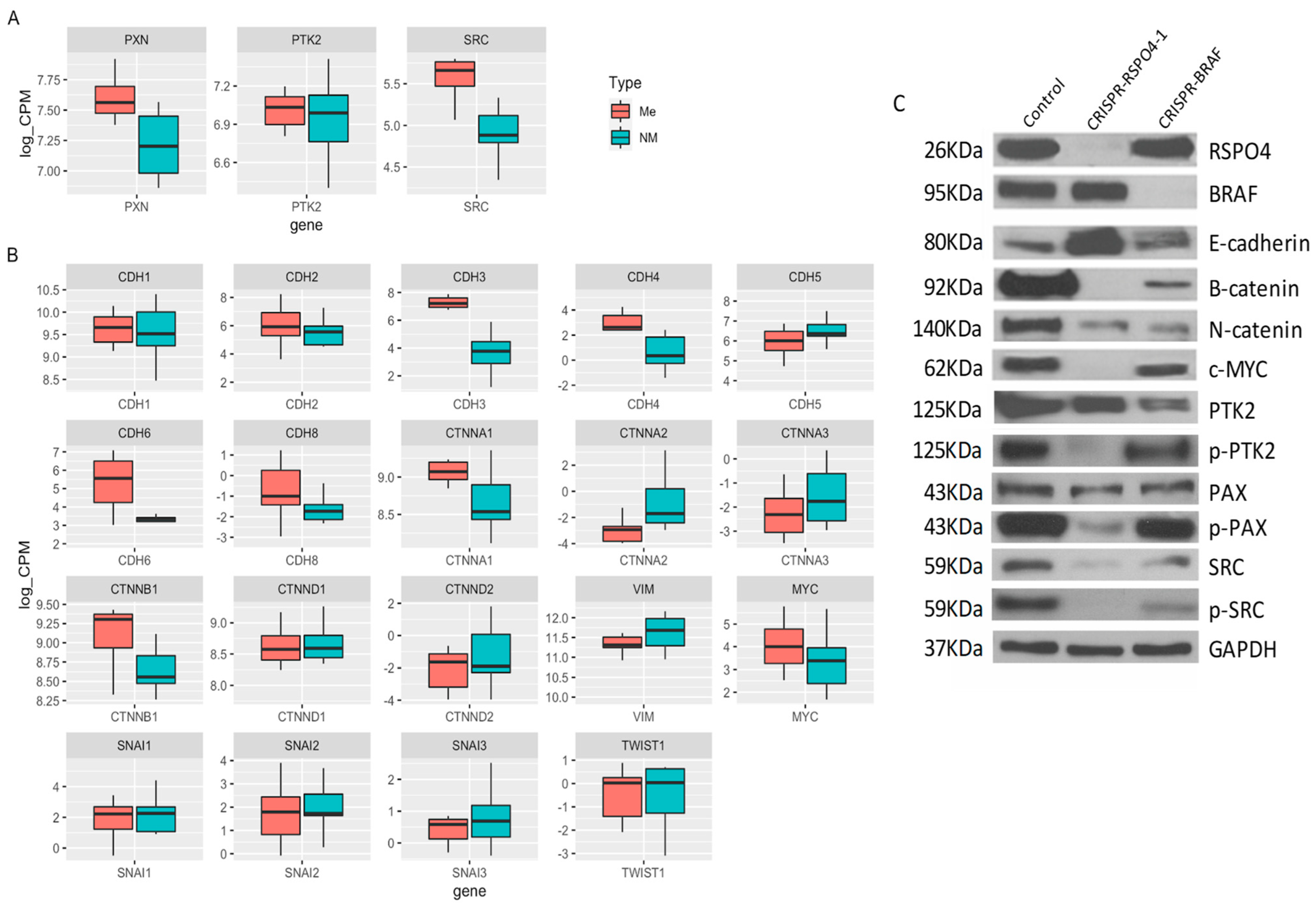
3.8. RSPO4 Inhibition in PTC Cells Prevents the Activation of Proteins Involved in Focal Adhesion Signaling
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Curado, M.P.; Edwards, B.; Shin, H.R.; Storm, H.; Ferlay, J.; Heanue, M.; Boyle, P. Cancer Incidence in Five Continents; IARC Scientific Publication No 160; IARC Press, International Agency for Research on Cancer: Lyon, France, 2007; Volume IX. [Google Scholar]
- Statistics. ACoC: Canadian Cancer Statistics 2013; Canadian Cancer Society: Toronto, ON, Canada, 2013; Available online: http://www.cancer.ca/Reference/?cceid=1700#ixzz2ZJNY0hbS (accessed on 1 September 2022).
- Davies, L.; Welch, H.G. Increasing incidence of thyroid cancer in the United States, 1973–2002. JAMA 2006, 295, 2164–2167. [Google Scholar] [CrossRef]
- Aschebrook-Kilfoy, B.; Kaplan, E.L.; Chiu, B.C.; Angelos, P.; Grogan, R.H. The Acceleration in Papillary Thyroid Cancer Incidence Rates is Similar among Racial and Ethnic Groups in the United States. Ann. Surg. Oncol. 2013, 20, 2746–2753. [Google Scholar] [CrossRef]
- Dom, G.; Tarabichi, M.; Unger, K.; Thomas, G.; Oczko-Wojciechowska, M.; Bogdanova, T.; Jarzab, B.; Dumont, J.E.; Detours, V.; Maenhaut, C. A gene expression signature distinguishes normal tissues of sporadic and radiation-induced papillary thyroid carcinomas. Br. J. Cancer 2012, 107, 994–1000. [Google Scholar] [CrossRef]
- Mazzaferri, E.L. Management of a solitary thyroid nodule. N. Engl. J. Med. 1993, 328, 553–559. [Google Scholar]
- Schlumberger, M.J. Papillary and follicular thyroid carcinoma. N. Engl. J. Med. 1998, 338, 297–306. [Google Scholar] [CrossRef]
- Ito, Y.; Miyauchi, A.; Kihara, M.; Fukushima, M.; Higashiyama, T.; Miya, A. Overall survival of papillary thyroid carcinoma patients: A single-institution long-term follow-up of 5897 patients. World J. Surg. 2018, 42, 615–622. [Google Scholar] [CrossRef]
- Ywata de Carvalho, A.; Kohler, H.F.; Gomes, C.C.; Vartanian, J.G.; Kowalski, L.P. Predictive factors for recurrence of papillary thyroid carcinoma: Analysis of 4085 patients. Acta Otorhinolaryngol. Ital. 2021, 41, 236–242. [Google Scholar] [CrossRef]
- Cooper, D.S.; Doherty, G.M.; Haugen, B.R.; Kloos, R.T.; Lee, S.L.; Mandel, S.J.; Mazzaferri, E.L.; McIver, B.; Pacini, F.; Schlumberger, M. Revised American Thyroid Association management guidelines for patients with thyroid nodules and differentiated thyroid cancer: The American Thyroid Association (ATA) guidelines taskforce on thyroid nodules and differentiated thyroid cancer. Thyroid Off. J. Am. Thyroid Assoc. 2009, 19, 1167–1214. [Google Scholar] [CrossRef]
- Haugen, B.R.; Sawka, A.M.; Alexander, E.K.; Bible, K.C.; Caturegli, P.; Doherty, G.M.; Mandel, S.J.; Morris, J.C.; Nassar, A.; Pacini, F. American Thyroid Association guidelines on the management of thyroid nodules and differentiated thyroid cancer task force review and recommendation on the proposed renaming of encapsulated follicular variant papillary thyroid carcinoma without invasion to noninvasive follicular thyroid neoplasm with papillary-like nuclear features. Thyroid Off. J. Am. Thyroid Assoc. 2017, 27, 481–483. [Google Scholar]
- Xing, M.; Haugen, B.R.; Schlumberger, M. Progress in molecular-based management of differentiated thyroid cancer. Lancet 2013, 381, 1058–1069. [Google Scholar] [CrossRef]
- Rossi, E.D.; Locantore, P.; Bruno, C.; Dell’Aquila, M.; Tralongo, P.; Curatolo, M.; Revelli, L.; Raffaelli, M.; Larocca, L.M.; Pantanowitz, L.; et al. Molecular Characterization of Thyroid Follicular Lesions in the Era of “Next-Generation” Techniques. Front. Endocrinol. 2022, 13, 834456. [Google Scholar] [CrossRef]
- Agrawal, N.; Akbani, R.; áArman Aksoy, B.; Ally, A.; Arachchi, H.; Asa, S.; áTodd Auman, J.; Balasundaram, M.; Balu, S.; Baylin, S.; et al. Integrated genomic characterization of papillary thyroid carcinoma. Cell 2014, 159, 676–690. [Google Scholar] [CrossRef]
- Boufraqech, M.; Nilubol, N. Multi-omics Signatures and Translational Potential to Improve Thyroid Cancer Patient Outcome. Cancers 2019, 11, 1988. [Google Scholar] [CrossRef]
- Morand, G.B.; da Silva, S.D.; Hier, M.P.; Alaoui-Jamali, M.A. Insights into genetic and epigenetic determinants with impact on vitamin d signaling and cancer association studies: The case of thyroid cancer. Front. Oncol. 2014, 4, 309. [Google Scholar] [CrossRef]
- Nucera, C.; Porrello, A.; Antonello, Z.A.; Mekel, M.; Nehs, M.A.; Giordano, T.J.; Gerald, D.; Benjamin, L.E.; Priolo, C.; Puxeddu, E.; et al. B-Raf(V600E) and thrombospondin-1 promote thyroid cancer progression. Proc. Natl. Acad. Sci. USA 2010, 107, 10649–10654. [Google Scholar] [CrossRef]
- Li, C.; Lee, K.C.; Schneider, E.B.; Zeiger, M.A. BRAF V600E mutation and its association with clinicopathological features of papillary thyroid cancer: A meta-analysis. J. Clin. Endocrinol. Metab. 2012, 97, 4559–4570. [Google Scholar] [CrossRef]
- Xing, M.; Alzahrani, A.S.; Carson, K.A.; Shong, Y.K.; Kim, T.Y.; Viola, D.; Elisei, R.; Bendlova, B.; Yip, L.; Mian, C.; et al. Association between BRAF V600E Mutation and Recurrence of Papillary Thyroid Cancer. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2015, 33, 42–50. [Google Scholar] [CrossRef]
- Morand, G.B.; da Silva, S.D.; Mlynarek, A.M.; Black, M.J.; Payne, R.J.; Hier, M.P. Clinicopathological relevance of antithyroglobulin antibodies in low-risk papillary thyroid cancer. Clin. Otolaryngol. Off. J. ENT-UK Off. J. Neth. Soc. Oto-Rhino-Laryngol. Cervico-Facial Surg. 2017, 42, 1130–1134. [Google Scholar] [CrossRef]
- Semsar-Kazerooni, K.; Morand, G.B.; Payne, A.E.; da Silva, S.D.; Forest, V.-I.; Hier, M.P.; Pusztaszeri, M.P.; Tamilia, M.; Payne, R.J. Mutational status may supersede tumor size in predicting the presence of aggressive pathologic features in well differentiated thyroid cancer. J. Otolaryngol.—Head Neck Surg. 2022, 51, 9. [Google Scholar] [CrossRef]
- Yip, L.; Gooding, W.E.; Nikitski, A.; Wald, A.I.; Carty, S.E.; Karslioglu-French, E.; Seethala, R.R.; Zandberg, D.P.; Ferris, R.L.; Nikiforova, M.N. Risk assessment for distant metastasis in differentiated thyroid cancer using molecular profiling: A matched case-control study. Cancer 2021, 127, 1779–1787. [Google Scholar] [CrossRef]
- Bourgey, M.; Dali, R.; Eveleigh, R.; Chen, K.C.; Letourneau, L.; Fillon, J.; Michaud, M.; Caron, M.; Sandoval, J.; Lefebvre, F.; et al. GenPipes: An open-source framework for distributed and scalable genomic analyses. Gigascience 2019, 8, giz037. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Roberts, A.; Pimentel, H.; Trapnell, C.; Pachter, L. Identification of novel transcripts in annotated genomes using RNA-Seq. Bioinformatics 2011, 27, 2325–2329. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Bustin, S.A.; Beaulieu, J.F.; Huggett, J.; Jaggi, R.; Kibenge, F.S.; Olsvik, P.A.; Penning, L.C.; Toegel, S. MIQE precis: Practical implementation of minimum standard guidelines for fluorescence-based quantitative real-time PCR experiments. BMC Mol. Biol. 2010, 11, 74. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- de Lima, J.M.; Morand, G.B.; Macedo, C.C.S.; Diesel, L.; Hier, M.P.; Mlynarek, A.; Kowalski, L.P.; Maschietto, M.; Alaoui-Jamali, M.A.; da Silva, S.D. NDRG1 deficiency is associated with regional metastasis in oral cancer by inducing epithelial-mesenchymal transition. Carcinogenesis 2020, 41, 769–777. [Google Scholar] [CrossRef]
- da Silva, S.D.; Marchi, F.A.; Su, J.; Yang, L.; Valverde, L.; Hier, J.; Bijian, K.; Hier, M.; Mlynarek, A.; Kowalski, L.P. Co-overexpression of TWIST1-CSF1 is a common event in metastatic oral cancer and drives biologically aggressive phenotype. Cancers 2021, 13, 153. [Google Scholar] [CrossRef]
- Blaydon, D.C.; Ishii, Y.; O’Toole, E.A.; Unsworth, H.C.; Teh, M.T.; Rüschendorf, F.; Sinclair, C.; Hopsu-Havu, V.K.; Tidman, N.; Moss, C.; et al. The gene encoding R-spondin 4 (RSPO4), a secreted protein implicated in Wnt signaling, is mutated in inherited anonychia. Nat. Genet. 2006, 38, 1245–1247. [Google Scholar] [CrossRef]
- Sulzmaier, F.J.; Jean, C.; Schlaepfer, D.D. FAK in cancer: Mechanistic findings and clinical applications. Nat. Rev. Cancer 2014, 14, 598–610. [Google Scholar] [CrossRef]
- Spano, D.; Zollo, M. Tumor microenvironment: A main actor in the metastasis process. Clin. Exp. Metastasis 2012, 29, 381–395. [Google Scholar] [CrossRef]
- Wong, G.S.; Rustgi, A.K. Matricellular proteins: Priming the tumour microenvironment for cancer development and metastasis. Br. J. Cancer 2013, 108, 755–761. [Google Scholar] [CrossRef]
- Fagin, J.A.; Wells, S.A., Jr. Biologic and clinical perspectives on thyroid cancer. N. Engl. J. Med. 2016, 375, 1054–1067. [Google Scholar] [CrossRef]
- Cabanillas, M.E.; McFadden, D.G.; Durante, C. Thyroid cancer. Lancet 2016, 388, 2783–2795. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Hanahan, D.; Coussens, L.M. Accessories to the crime: Functions of cells recruited to the tumor microenvironment. Cancer Cell 2012, 21, 309–322. [Google Scholar] [CrossRef]
- Kim, K.A.; Wagle, M.; Tran, K.; Zhan, X.; Dixon, M.A.; Liu, S.; Gros, D.; Korver, W.; Yonkovich, S.; Tomasevic, N.; et al. R-Spondin family members regulate the Wnt pathway by a common mechanism. Mol. Biol. Cell 2008, 19, 2588–2596. [Google Scholar] [CrossRef]
- Carmon, K.S.; Gong, X.; Lin, Q.; Thomas, A.; Liu, Q. R-spondins function as ligands of the orphan receptors LGR4 and LGR5 to regulate Wnt/beta-catenin signaling. Proc. Natl. Acad. Sci. USA 2011, 108, 11452–11457. [Google Scholar] [CrossRef] [PubMed]
- De Lau, W.; Barker, N.; Low, T.Y.; Koo, B.K.; Li, V.S.; Teunissen, H.; Kujala, P.; Haegebarth, A.; Peters, P.J.; Van De Wetering, M.; et al. Lgr5 homologues associate with Wnt receptors and mediate R-spondin signalling. Nature 2011, 476, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Glinka, A.; Dolde, C.; Kirsch, N.; Huang, Y.L.; Kazanskaya, O.; Ingelfinger, D.; Boutros, M.; Cruciat, C.M.; Niehrs, C. LGR4 and LGR5 are R-spondin receptors mediating Wnt/β-catenin and Wnt/PCP signalling. EMBO Rep. 2011, 12, 1055–1061. [Google Scholar] [CrossRef] [PubMed]
- Tejeda-Muñoz, N.; Morselli, M.; Moriyama, Y.; Sheladiya, P.; Pellegrini, M.; De Robertis, E.M. Canonical Wnt signaling induces focal adhesion and Integrin beta-1 endocytosis. iScience 2022, 25, 104123. [Google Scholar] [CrossRef] [PubMed]
- Provenzano, P.P.; Inman, D.R.; Eliceiri, K.W.; Keely, P.J. Matrix density-induced mechanoregulation of breast cell phenotype, signaling and gene expression through a FAK-ERK linkage. Oncogene 2009, 28, 4326–4343. [Google Scholar] [CrossRef]
- Klein, E.A.; Yin, L.; Kothapalli, D.; Castagnino, P.; Byfield, F.J.; Xu, T.; Levental, I.; Hawthorne, E.; Janmey, P.A.; Assoian, R.K. Cell-cycle control by physiological matrix elasticity and in vivo tissue stiffening. Curr. Biol. 2009, 19, 1511–1518. [Google Scholar] [CrossRef]
- Bjerke, M.A.; Dzamba, B.J.; Wang, C.; DeSimone, D.W. FAK is required for tension-dependent organization of collective cell movements in Xenopus mesendoderm. Dev. Biol. 2014, 394, 340–356. [Google Scholar] [CrossRef]
- Sun, Z.; Guo, S.S.; Fässler, R. Integrin-mediated mechanotransduction. J. Cell Biol. 2016, 215, 445–456. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group Variable | Non-Metastatic | Metastatic | p-Value * |
|---|---|---|---|
| Histological subtype | 10/10 papillary | 10/10 papillary | 0.99 |
| Age (mean, SE) | 45.9 (4.7) | 46.3 (4.8) | 0.95 |
| Gender (f/m) | 7/3 | 6/4 | 0.99 |
| pT category → (T3/T2-1) | 4/6 | 4/6 | 0.99 |
| pN category | pN0 (10/10) | pN+ (10/10) | <0.001 |
| Gene | NM | Clone |
|---|---|---|
| BRAF | NM_004333 | TRCN0000231130 |
| RSPO4-1 | NM_001029871 | TRCN0000139566 |
| RSPO4-2 | NM_001029871 | TRCN0000139399 |
| RSPO4-3 | NM_001029871 | TRCN0000139901 |
| RSPO4-4 | NM_001029871 | TRCN0000139862 |
| Gene Symbol | Forward Sequence (5′–3′) | Reverse Sequence (5′–3′) |
|---|---|---|
| ACTB | GCACCCAGCACAATGAAG- | GCACCCAGCACAATGAAG- |
| BRAF | GAAGGTGAAGGTCGGA | GGGTCATTGATGGCAAC |
| CDH1 | CCTGGGACTCCACCTACAGA | CCTGGGACTCCACCTACAGA |
| CTNNB1 | ATTGTCCACGCTGGATTTTC | TCGAGGACGGTCGGACT |
| GAPDH | AATACACCAGCAAGCTAGATGC | AATCAGTTCCGTTCCCCAGAG |
| HPRT1 | GAACGTCTTGCTCGAGATGTGA | TCCAGCAGGTCAGCAAAGAAT |
| MYC | ATCCAGCGTCTAAGCAGCTG | TACAACACCCGAGCAAGGAC |
| PXN | AGCTAGCGCGACCCTGA | TGTGGGAGGTGGTAGACTCC |
| PTK2 | CCTGGTCCACTTGATCAGCTA | GCCAAAAGGATTTCTAAACCAG |
| RSPO4 | GTGGAACAGCCGTTCTCCTCT | GAAGGAAGAAGCAAGTGGGC |
| SNAIL | CCAGTGCCTCGACCACTATG | CTGCTGGAAGGTAAACTCTGGA |
| TWIST | TCCATTTTCTCCTTCTCTGGAA | CCTTCTCGGTCTGGAGGAT |
| VIM | CTTCAGAGAGAGGAAGCCGA | ATTCCACTTTGCGTTCAAGG |
| Characteristics | Benign Cases (n = 15) | PTC Matched from Seq Patients (n = 20) | PTC Long Follow-Up (n = 136) | All Patients (n = 171) | p-Value |
|---|---|---|---|---|---|
| Age (years, mean ± SD) | 57.3 (14.6) | 46.0 (14.7) | 46.0 (14.4) | 47.0 (14.6) | 0.025 |
| Gender Female Male | 9 (60.0%) 6 (40.0%) | 13 (65.0%) 7 (35.0%) | 112 (82.4%) 24 (17.6%) | 134 (78.4%) 37 (21.6%) | 0.041 |
| Surgery Total thyroidectomy Hemithyroidectomy | 12 (80%) 3 (20%) | 15 (75%) 5 (25%) | 126 (92.6%) 10 (7.4%) | 153 (89.5%) 18 (10.5%) | 0.027 |
| pT category pT1 pT2 pT3/4 | na * | 4 (20.0%) 5 (25.0%) 11(55.0%) | 21 (15.4%) 33 (24.3%) 82 (60.3%) | 25 (16.0%) 38 (24.4%) 93 (59.6%) | 0.001 |
| pN category pN0 pN1a pN1b pNx | na * | 10 (50.0%) 6 (30.0%) 4 (20.0%) 0 | 74 (54.4%) 29 (21.3%) 18 (13.2%) 15 (11.0%) | 84 (53.8%) 35 (22.4%) 22 (14.2%) 15 (9.6%) | 0.001 |
| Follow-up (months, mean ± SD) | 14.5 (11.2) | 15.7 (7.3%) | 68.1 (30.9) | 59.8 (34.5) | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva, S.D.; Morand, G.B.; Diesel, L.; de Lima, J.M.; Bijian, K.; Kailasam, S.; Lefebvre, F.; Bourque, G.; Hier, M.; Alaoui-Jamali, M.A. Identification of R-Spondin Gene Signature Predictive of Metastatic Progression in BRAFV600E-Positive Papillary Thyroid Cancer. Cells 2023, 12, 139. https://doi.org/10.3390/cells12010139
da Silva SD, Morand GB, Diesel L, de Lima JM, Bijian K, Kailasam S, Lefebvre F, Bourque G, Hier M, Alaoui-Jamali MA. Identification of R-Spondin Gene Signature Predictive of Metastatic Progression in BRAFV600E-Positive Papillary Thyroid Cancer. Cells. 2023; 12(1):139. https://doi.org/10.3390/cells12010139
Chicago/Turabian Styleda Silva, Sabrina Daniela, Grégoire B. Morand, Luciana Diesel, Jefferson Muniz de Lima, Krikor Bijian, Senthilkumar Kailasam, Francois Lefebvre, Guillaume Bourque, Michael Hier, and Moulay A. Alaoui-Jamali. 2023. "Identification of R-Spondin Gene Signature Predictive of Metastatic Progression in BRAFV600E-Positive Papillary Thyroid Cancer" Cells 12, no. 1: 139. https://doi.org/10.3390/cells12010139
APA Styleda Silva, S. D., Morand, G. B., Diesel, L., de Lima, J. M., Bijian, K., Kailasam, S., Lefebvre, F., Bourque, G., Hier, M., & Alaoui-Jamali, M. A. (2023). Identification of R-Spondin Gene Signature Predictive of Metastatic Progression in BRAFV600E-Positive Papillary Thyroid Cancer. Cells, 12(1), 139. https://doi.org/10.3390/cells12010139