
Astrocyte L-Lactate Signaling in the ACC Regulates Visceral Pain Aversive Memory in Rats

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiments
2.2. Cannula Implantation
2.3. Drugs Administration
2.4. Optogenetic Manipulation of ACC Astrocytes
2.5. Chemogenetic Manipulation of ACC Astrocytes
2.6. Behavioral Paradigm
2.7. In Vivo Microdialysis
2.7.1. Experiment 1: Measurement of the Training-Dependent Extracellular L-Lactate Level in the ACC
2.7.2. Experiment 2: Extracellular L-Lactate Level in Response to Bilateral Infusion of DAB into the ACC
2.7.3. Experiment 3: L-Lactate Level in Response to Optogenetic Activation of ACC Astrocytes
2.7.4. Experiment 4: L-Lactate level in response to chemogenetic manipulation of ACC astrocytes
2.7.5. L-Lactate measurement and analysis
2.8. Immunohistochemistry
2.9. Western Blot Analysis
2.10. Statistical Analysis
3. Results
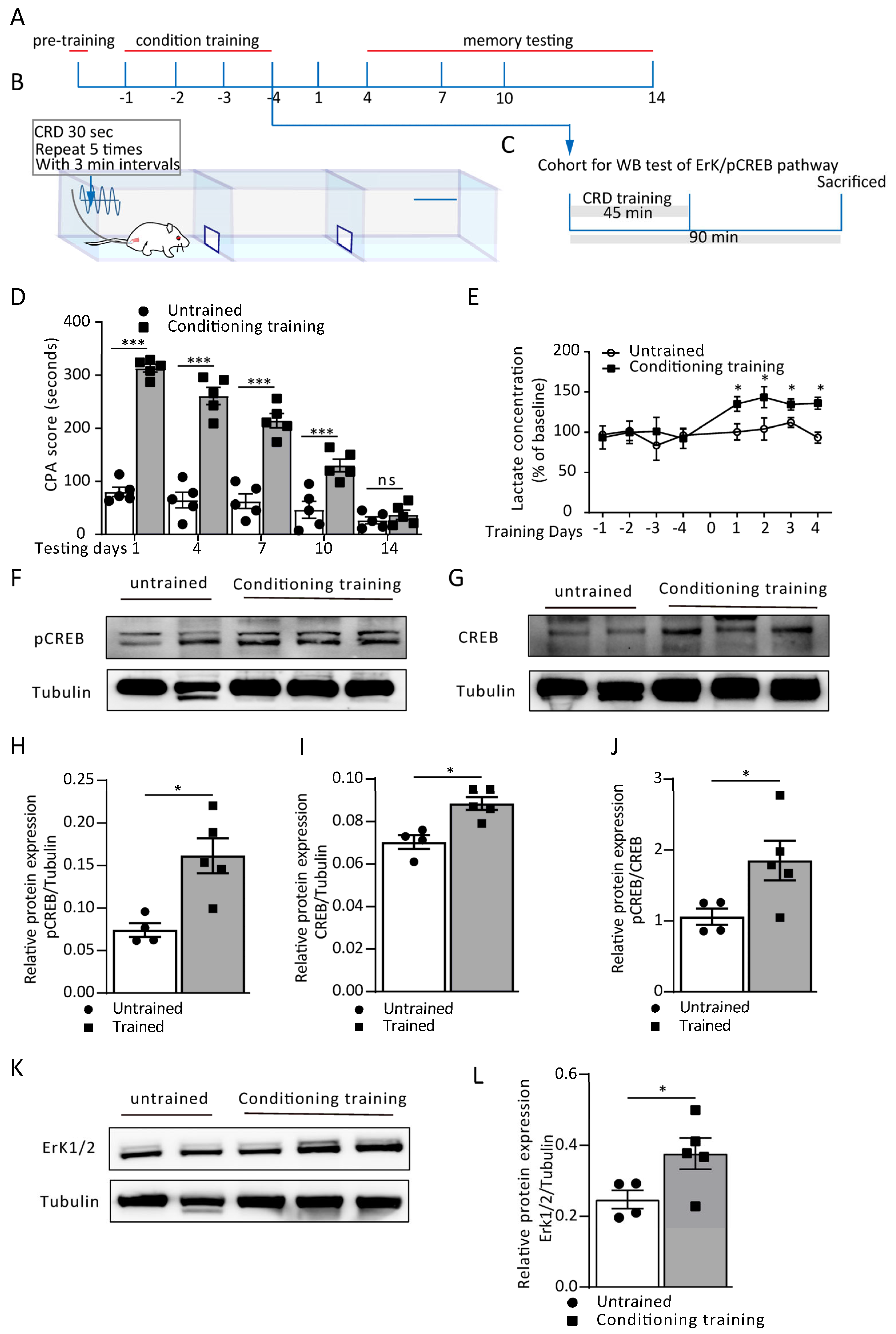
3.1. Conditioning Training-Induced Lactate Release from ACC Astrocytes Contributes to Aversive Memory Formation
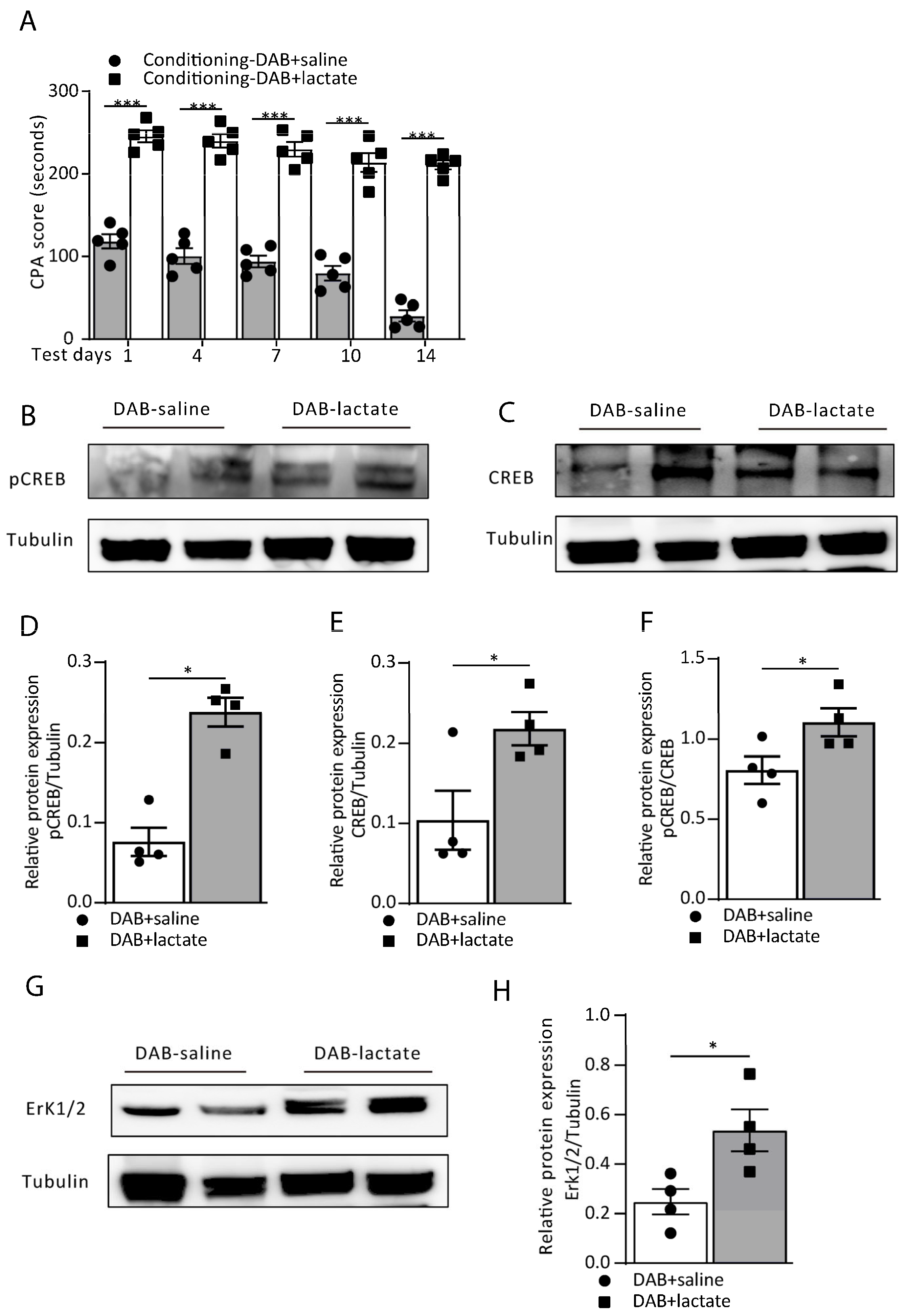
3.2. Blocking Astrocytic Glycogenolysis in the ACC Disrupts Aversive Memory by Decreasing the L-Lactate Level
3.3. Optogenetic Activation of ACC Astrocytes Reverses Aversive Memory Formation from Glycogenolysis Blocking by Increasing the L-Lactate Level
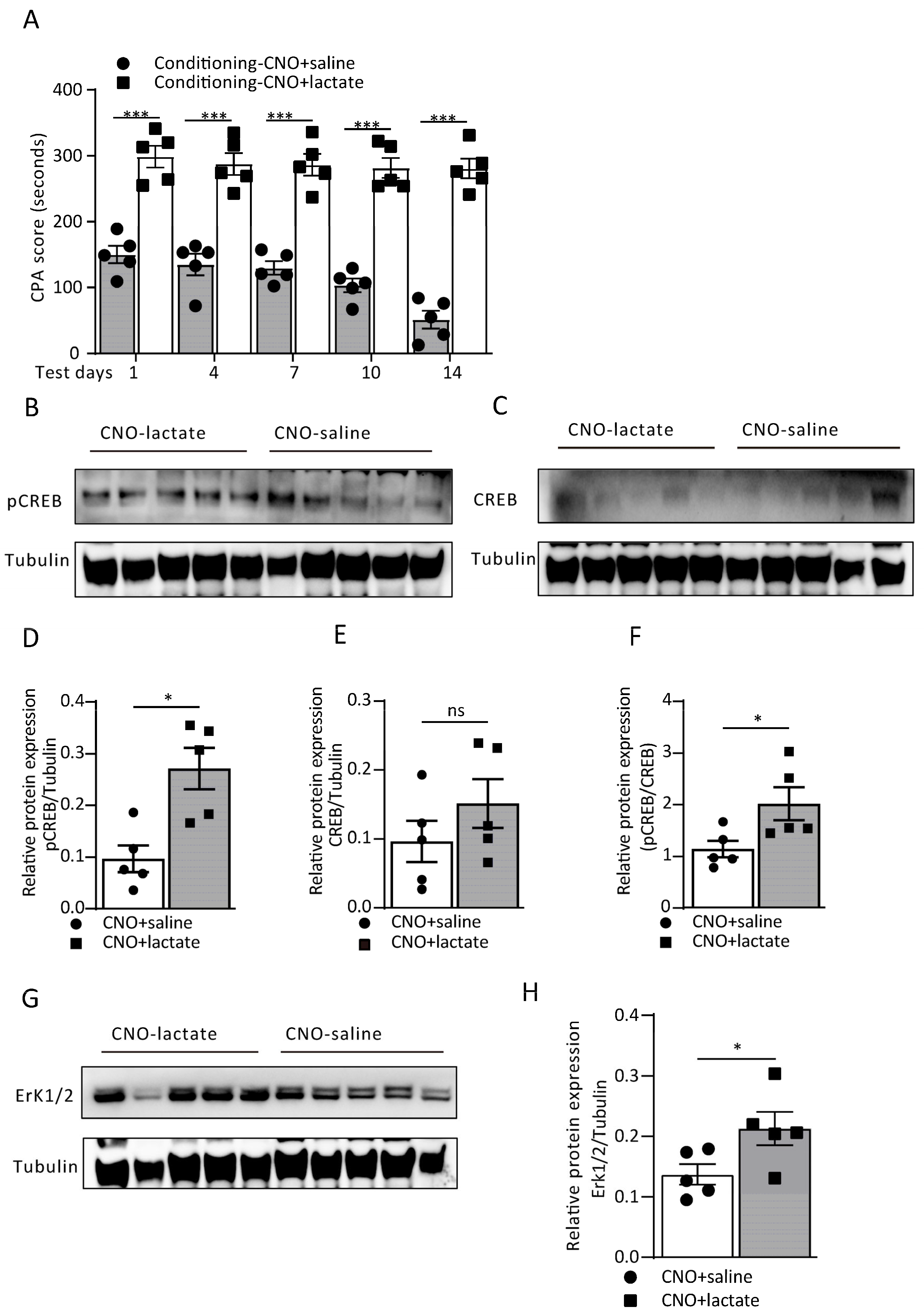
3.4. Astrocytic Gi Pathway Activation in the ACC Disrupts Aversive Memory Formation by Decreasing the L-Lactate Level
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Baliki, M.N.; Apkarian, A.V. Nociception, Pain, Negative Moods, and Behavior Selection. Neuron 2015, 87, 474–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogt, B.A.; Sikes, R.W.; Vogt, L.J. Anterior Cingulate Cortex and the Medial Pain System. In Neurobiology of Cingulate Cortex and Limbic Thalamus; Birkhäuser: Boston, MA, USA, 1993; pp. 313–344. [Google Scholar]
- Li, Y. Synaptic Plasticity and Synchrony in the Anterior Cingulate Cortex Circuitry: A Neural Network Approach to Causality of Chronic Visceral Pain and Associated Cognitive Deficits. Adv Neurobiol. 2018, 21, 219–245. [Google Scholar] [CrossRef]
- May, A. Chronic pain may change the structure of the brain. Pain 2008, 137, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Wu, X.; Owyang, C.; Li, Y. Enhanced responses of the anterior cingulate cortex neurones to colonic distension in viscerally hypersensitive rats. J. Physiol. 2006, 570 Pt 1, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, X.; Cao, B.; Liu, J.; Li, Y. Facilitation of synaptic transmission in the anterior cingulate cortex in viscerally hypersensitive rats. Cereb. Cortex 2015, 25, 859–868. [Google Scholar] [CrossRef] [Green Version]
- Cao, Z.; Wu, X.; Chen, S.; Fan, J.; Zhang, R.; Owyang, C.; Ying, L. Anterior cingulate cortex modulates visceral pain as measured by visceromotor responses in viscerally hypersensitive rats. Gastroenterology 2008, 134, 535–543. [Google Scholar] [CrossRef]
- Fan, J.; Wu, X.; Cao, Z.; Chen, S.; Owyang, C.; Li, Y. Up-regulation of anterior cingulate cortex NR2B receptors contributes to visceral pain responses in rats. Gastroenterology 2009, 136, 1732–1740.e3. [Google Scholar] [CrossRef] [Green Version]
- Cao, B.; Wang, J.; Mu, L.; Poon, D.C.-H.; Li, Y. Impairment of decision making associated with disruption of phase-locking in the anterior cingulate cortex in viscerally hypersensitive rats. Exp. Neurol. 2016, 286, 21–31. [Google Scholar] [CrossRef]
- Wang, J.; Tu, J.; Cao, B.; Mu, L.; Yang, X.; Cong, M.; Aruna, S.R.; Rosa, H.M.C.; Liping, W.; Ying, L. Astrocytic l-Lactate Signaling Facilitates Amygdala-Anterior Cingulate Cortex Synchrony and Decision Making in Rats. Cell Rep. 2017, 21, 2407–2418. [Google Scholar] [CrossRef]
- Icenhour, A.; Langhorst, J.; Benson, S.; Schlamann, M.; Hampel, S.; Engler, H.; Forsting, M.; Elsenbruch, S. Neural circuitry of abdominal pain-related fear learning and reinstatement in irritable bowel syndrome. Neurogastroenterol. Motil. 2015, 27, 114–127. [Google Scholar] [CrossRef]
- Mayer, E.A.; Naliboff, B.D.; Craig, A.D. Neuroimaging of the brain-gut axis: From basic understanding to treatment of functional GI disorders. Gastroenterology 2006, 131, 1925–1942. [Google Scholar] [CrossRef]
- Bushnell, M.C.; Ceko, M.; Low, L.A. Cognitive and emotional control of pain and its disruption in chronic pain. Nat. Rev. Neurosci. 2013, 14, 502–511. [Google Scholar] [CrossRef] [Green Version]
- Woodworth, R.S.; Sherrington, C.S. A pseudaffective reflex and its spinal path. J. Physiol. 1904, 31, 234–243. [Google Scholar] [CrossRef] [Green Version]
- Cao, B.; Zhang, X.; Yan, N.; Chen, S.; Li, Y. Cholecystokinin enhances visceral pain-related affective memory via vagal afferent pathway in rats. Mol. Brain 2012, 5, 19. [Google Scholar] [CrossRef] [Green Version]
- Yan, N.; Cao, B.; Xu, J.; Hao, C.; Zhang, X.; Li, Y. Glutamatergic activation of anterior cingulate cortex mediates the affective component of visceral pain memory in rats. Neurobiol. Learn. Mem. 2012, 97, 156–164. [Google Scholar] [CrossRef]
- Bezzi, P.; Volterra, A. Astrocytes: Powering memory. Cell 2011, 144, 644–645. [Google Scholar] [CrossRef] [Green Version]
- Kol, A.; Adamsky, A.; Groysman, M.; Kreisel, T.; London, M.; Goshen, I. Astrocytes contribute to remote memory formation by modulating hippocampal-cortical communication during learning. Nat. Neurosci. 2020, 23, 1229–1239. [Google Scholar] [CrossRef]
- Suzuki, A.; Stern, S.A.; Bozdagi, O.; Huntley, G.W.; Walker, R.H.; Magistretti, P.J.; Cristina, M.A. Astrocyte-neuron lactate transport is required for long-term memory formation. Cell 2011, 144, 810–823. [Google Scholar] [CrossRef] [Green Version]
- Verkhratsky, A.; Nedergaard, M. Physiology of Astroglia. Physiol. Rev. 2018, 98, 239–389. [Google Scholar] [CrossRef]
- Pellerin, L.; Magistretti, P.J. Glutamate uptake into astrocytes stimulates aerobic glycolysis: A mechanism coupling neuronal activity to glucose utilization. Proc. Natl. Acad. Sci. USA 1994, 91, 10625–10629. [Google Scholar] [CrossRef]
- Hu, Y.; Wilson, G.S. A temporary local energy pool coupled to neuronal activity: Fluctuations of extracellular lactate levels in rat brain monitored with rapid-response enzyme-based sensor. J. Neurochem. 1997, 69, 1484–1490. [Google Scholar] [CrossRef] [PubMed]
- Prichard, J.; Rothman, D.; Novotny, E.; Petroff, O.; Kuwabara, T.; Avison, M.; Howseman, A.; Hanstock, C.; Shulman, R. Lactate rise detected by 1H NMR in human visual cortex during physiologic stimulation. Proc. Natl. Acad. Sci. USA 1991, 88, 5829–5831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibbs, M.E.; Anderson, D.G.; Hertz, L. Inhibition of glycogenolysis in astrocytes interrupts memory consolidation in young chickens. Glia 2006, 54, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Walls, A.B.; Sickmann, H.M.; Brown, A.; Bouman, S.D.; Ransom, B.; Schousboe, A.; Waagepetersen, H.S. Characterization of 1,4-dideoxy-1,4-imino-d-arabinitol (DAB) as an inhibitor of brain glycogen shunt activity. J. Neurochem. 2008, 105, 1462–1470. [Google Scholar] [CrossRef] [PubMed]
- Adamsky, A.; Kol, A.; Kreisel, T.; Doron, A.; Ozeri-Engelhard, N.; Melcer, T.; Refaeli, R.; Horn, H.; Regev, L.; Groysman, M.; et al. Astrocytic Activation Generates De Novo Neuronal Potentiation and Memory Enhancement. Cell 2018, 174, 59–71.e14. [Google Scholar] [CrossRef] [Green Version]
- Johansen, J.P.; Fields, H.L.; Manning, B.H. The affective component of pain in rodents: Direct evidence for a contribution of the anterior cingulate cortex. Proc. Natl. Acad. Sci. USA 2001, 98, 8077–8082. [Google Scholar] [CrossRef] [Green Version]
- Llorca-Torralba, M.; Suárez-Pereira, I.; Bravo, L.; Camarena-Delgado, C.; Garcia-Partida, J.A.; Mico, J.A.; Berrocoso, E. Chemogenetic Silencing of the Locus Coeruleus-Basolateral Amygdala Pathway Abolishes Pain-Induced Anxiety and Enhanced Aversive Learning in Rats. Biol. Psychiatry 2019, 85, 1021–1035. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Gao, J.; Yan, J.; Fan, J.; Owyang, C.; Li, Y. Role for NMDA receptors in visceral nociceptive transmission in the anterior cingulate cortex of viscerally hypersensitive rats. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G918–G927. [Google Scholar] [CrossRef] [Green Version]
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: Biology and pathology. Acta Neuropathol. 2010, 119, 7–35. [Google Scholar] [CrossRef] [Green Version]
- Santello, M.; Toni, N.; Volterra, A. Astrocyte function from information processing to cognition and cognitive impairment. Nat. Neurosci. 2019, 22, 154–166. [Google Scholar] [CrossRef]
- Attwell, D.; Laughlin, S.B. An energy budget for signaling in the grey matter of the brain. J. Cereb. Blood Flow Metab. 2001, 21, 1133–1145. [Google Scholar] [CrossRef]
- Tang, F.; Lane, S.; Korsak, A.; Paton, J.F.R.; Gourine, A.V.; Kasparov, S.; Teschemacher, A.G. Lactate-mediated glia-neuronal signalling in the mammalian brain. Nat. Commun. 2014, 5, 3284. [Google Scholar] [CrossRef] [Green Version]
- Cataldo, A.M.; Broadwell, R.D. Cytochemical identification of cerebral glycogen and glucose-6-phosphatase activity under normal and experimental conditions. II. Choroid plexus and ependymal epithelia, endothelia and pericytes. J. Neurocytol. 1986, 15, 511–524. [Google Scholar] [CrossRef]
- Dringen, R.; Gebhardt, R.; Hamprecht, B. Glycogen in astrocytes: Possible function as lactate supply for neighboring cells. Brain Res. 1993, 623, 208–214. [Google Scholar] [CrossRef]
- Shulman, R.G.; Hyder, F.; Rothman, D.L. Cerebral energetics and the glycogen shunt: Neurochemical basis of functional imaging. Proc. Natl. Acad. Sci. USA 2001, 98, 6417–6422. [Google Scholar] [CrossRef] [Green Version]
- Rouach, N.; Koulakoff, A.; Abudara, V.; Willecke, K.; Giaume, C. Astroglial metabolic networks sustain hippocampal synaptic transmission. Science 2008, 322, 1551–1555. [Google Scholar] [CrossRef]
- Magistretti, P.J.; Allaman, I. Lactate in the brain: From metabolic end-product to signalling molecule. Nat. Rev. Neurosci. 2018, 19, 235–249. [Google Scholar] [CrossRef]
- Lauritzen, K.H.; Morland, C.; Puchades, M.; Holm-Hansen, S.; Hagelin, E.M.; Lauritzen, F.; Attramadal, H.; Storm-Mathisen, J.; Gjedde, A.; Bergersen, L.H. Lactate receptor sites link neurotransmission, neurovascular coupling, and brain energy metabolism. Cereb. Cortex 2014, 24, 2784–2795. [Google Scholar] [CrossRef]
- Morland, C.; Lauritzen, K.H.; Puchades, M.; Holm-Hansen, S.; Andersson, K.; Gjedde, A.; Attramadal, H.; Storm-Mathisen, J.; Bergersen, L.H. The lactate receptor, G-protein coupled receptor81/hydroxycarboxylic acid receptor 1: Expression and action in brain. J. Neurosci. Res. 2015, 93, 1043–1055. [Google Scholar] [CrossRef]
- Briquet, M.; Rocher, A.B.; Alessandri, M.; Rosenberg, N.; de Castro Abrantes, H.; Wellbourne-Wood, J.; Schmuziger, C.; Ginet, V.; Puyal, J.; Pralong, E.; et al. Activation of lactate receptor HCAR1 down-modulates neuronal activity in rodent and human brain tissue. J. Cereb. Blood Flow Metab. 2022, 42, 1650–1665. [Google Scholar] [CrossRef]
- De Castro Abrantes, H.; Briquet, M.; Schmuziger, C.; Restivo, L.; Puyal, J.; Rosenberg, N.; Rocher, A.B.; Offermanns, S.; Chatton, J.V. The lactate receptor HCAR1 modulates neuronal network activity through the activation of Gα and Gβγ subunits. J. Neurosci. 2019, 39, 4422–4433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallenius, K.; Thalen, P.; Bjorkman, J.A.; Petra Johannesson, P.; Wiseman, J.; Böttcher, G.; Fjellström, O.; Oakes, N.D. Involvement of the metabolic sensor GPR81 in cardiovascular control. JCI Insight 2017, 2, e92564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Ruchti, E.; Petit, J.M.; Jourdain, P.; Grenningloh, G.; Allaman, I.; Magistretti, P.J. Lactate promotes plasticity gene expression by potentiating NMDA signaling in neurons. Proc. Natl. Acad. Sci. USA 2014, 111, 12228–12233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosienko, V.; Teschemacher, A.G.; Kasparov, S. Is L-lactate a novel signaling molecule in the brain? J. Cereb. Blood Flow Metab. 2015, 35, 1069–1075. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iqbal, Z.; Liu, S.; Lei, Z.; Ramkrishnan, A.S.; Akter, M.; Li, Y. Astrocyte L-Lactate Signaling in the ACC Regulates Visceral Pain Aversive Memory in Rats. Cells 2023, 12, 26. https://doi.org/10.3390/cells12010026
Iqbal Z, Liu S, Lei Z, Ramkrishnan AS, Akter M, Li Y. Astrocyte L-Lactate Signaling in the ACC Regulates Visceral Pain Aversive Memory in Rats. Cells. 2023; 12(1):26. https://doi.org/10.3390/cells12010026
Chicago/Turabian StyleIqbal, Zafar, Shu Liu, Zhuogui Lei, Aruna Surendran Ramkrishnan, Mastura Akter, and Ying Li. 2023. "Astrocyte L-Lactate Signaling in the ACC Regulates Visceral Pain Aversive Memory in Rats" Cells 12, no. 1: 26. https://doi.org/10.3390/cells12010026
APA StyleIqbal, Z., Liu, S., Lei, Z., Ramkrishnan, A. S., Akter, M., & Li, Y. (2023). Astrocyte L-Lactate Signaling in the ACC Regulates Visceral Pain Aversive Memory in Rats. Cells, 12(1), 26. https://doi.org/10.3390/cells12010026