
A Novel Tongue Squamous Cell Carcinoma Cell Line Escapes from Immune Recognition due to Genetic Alterations in HLA Class I Complex
 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Approval and Consent to Participate
2.2. Cell Culture
2.3. Short Tandem Repeat (STR) Analysis
2.4. Antibodies for Immunocytochemistry/Immunohistochemistry Staining
2.5. In Situ Hybridization of Human Papillomavirus
2.6. Karyotping of the Cell Line
2.7. Population Doubling Time
2.8. Spheroid Formation Assay
2.9. Orthotopic Transplantation Tumors of Tongue
2.10. Whole-Exome Sequencing (WES)
2.11. Western Blot
2.12. CFSE Labeling Proliferation Assay and Tumor Cell Killing Assay
2.13. Statistical Analysis
3. Results
3.1. Clinical History
3.2. Histopathology of the Donor
3.3. Establishment of WU-TSC-1 Cell Line
3.4. Neoplastic Characteristics of WU-TSC-1 In Vitro and In Vivo
3.5. WU-TSC-1 Was PD-L1 Negative
3.6. The Landscape of Chromosome Variations
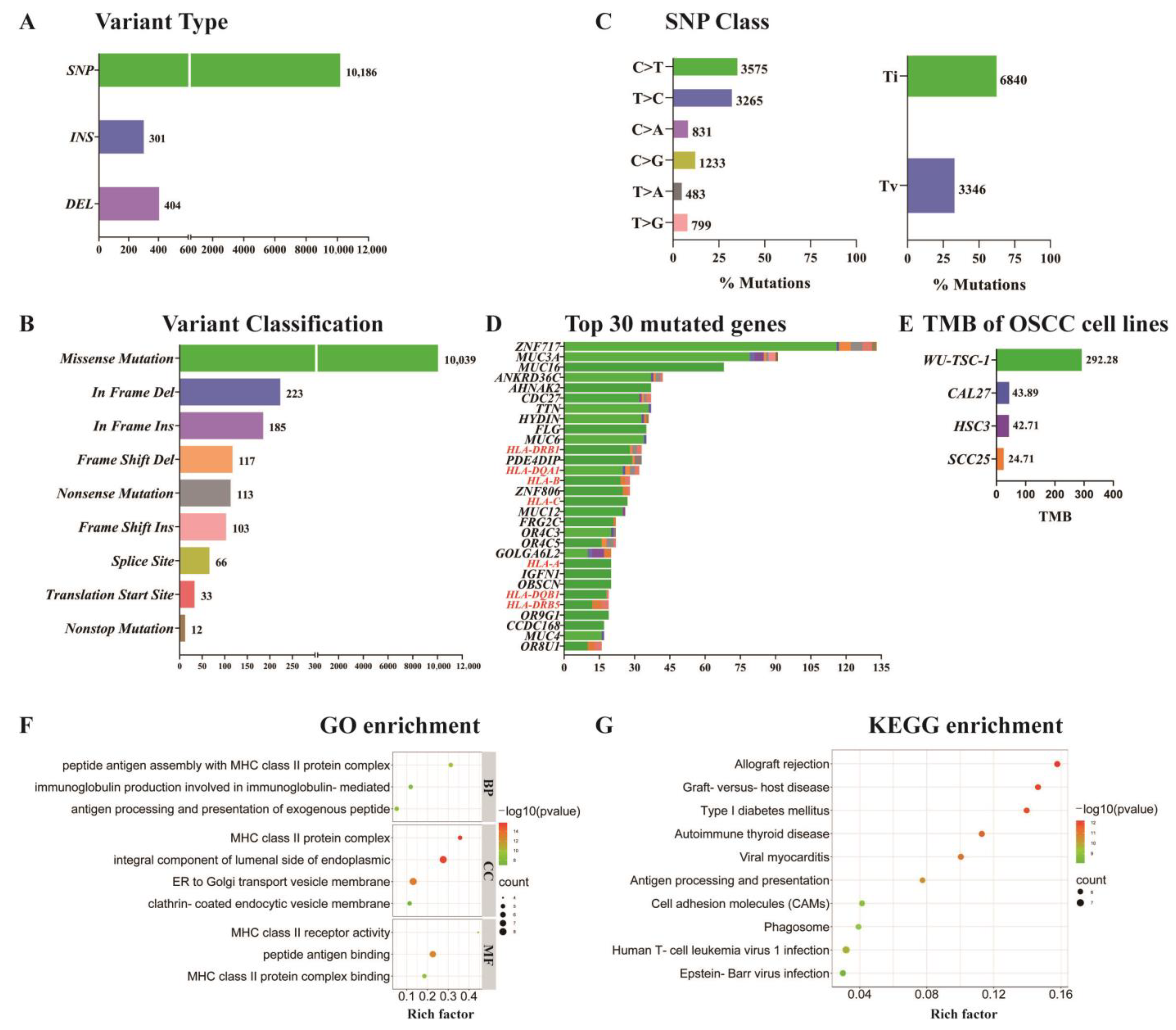
3.7. SNP/InDel Mutation Analysis
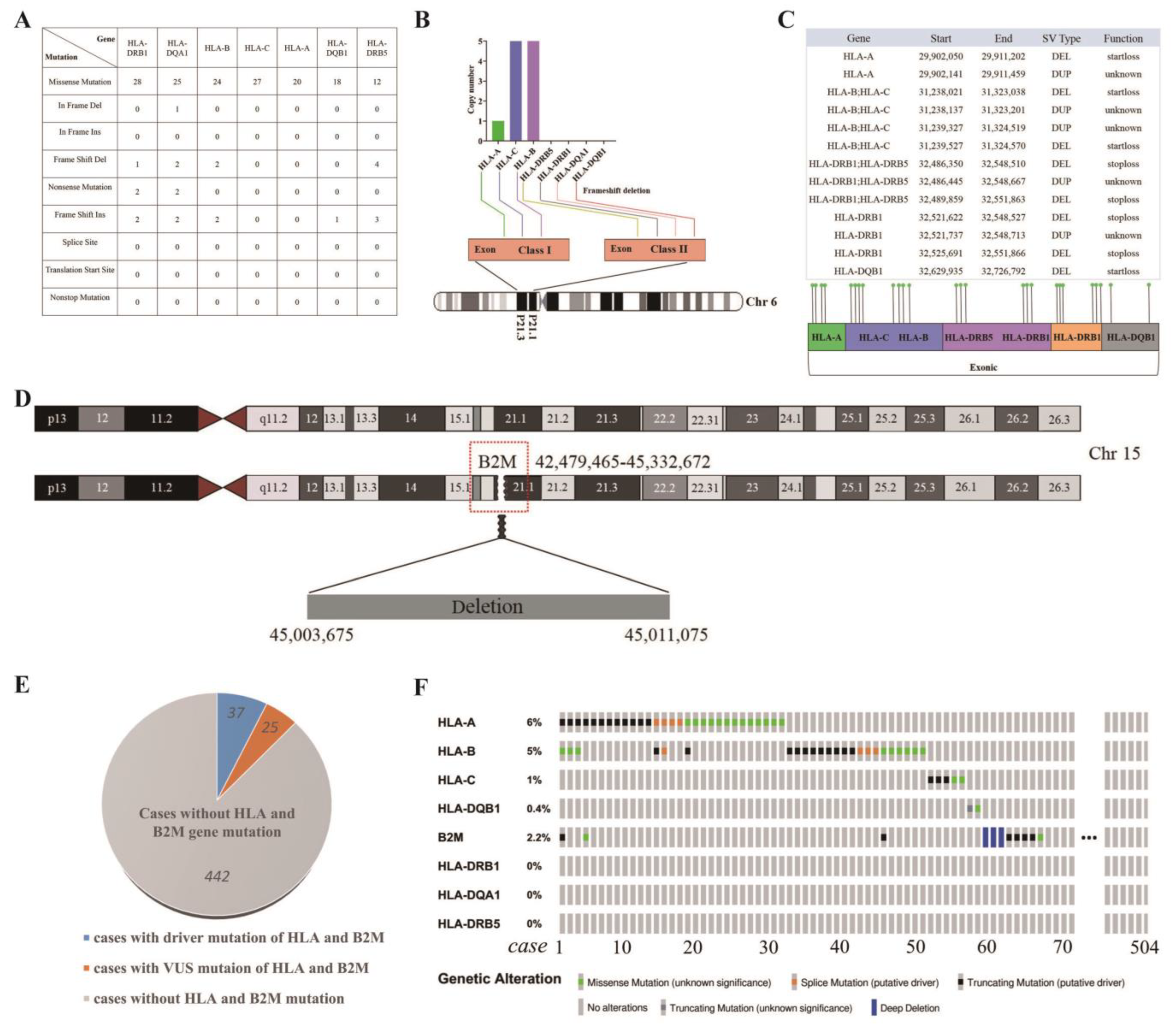
3.8. Distinctive Genetic Mutations in HLA Class I/II and B2M in WU-TSC-1
3.9. Defective Expression of HLA Class I Complex in WU-TSC-1
3.10. WU-TSC-1 Evades CD8+ T Cell Recognition
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vigneswaran, N.; Williams, M.D. Epidemiologic trends in head and neck cancer and aids in diagnosis. Oral Maxillofac. Surg. Clin. 2014, 26, 123–141. [Google Scholar] [CrossRef] [PubMed]
- Hussein, A.A.; Forouzanfar, T.; Bloemena, E.; De Visscher, J.; Brakenhoff, R.H.; Leemans, C.R.; Helder, M.N. A review of the most promising biomarkers for early diagnosis and prognosis prediction of tongue squamous cell carcinoma. Br. J. Cancer 2018, 119, 724–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sano, D.; Myers, J.N. Metastasis of squamous cell carcinoma of the oral tongue. Cancer Metastasis Rev. 2007, 26, 645–662. [Google Scholar] [CrossRef] [PubMed]
- Siriwardena, S.B.S.M.; Tsunematsu, T.; Qi, G.; Ishimaru, N.; Kudo, Y. Invasion-Related Factors as Potential Diagnostic and Therapeutic Targets in Oral Squamous Cell Carcinoma—A Review. Int. J. Mol. Sci. 2018, 19, 1462. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.E.; Burtness, B.; Leemans, C.R.; Lui, V.W.Y.; Bauman, J.E.; Grandis, J.R. Head and neck squamous cell carcinoma. Nat. Rev. Dis. Primers 2020, 6, 92. [Google Scholar] [CrossRef]
- Magnes, T.; Wagner, S.; Kiem, D.; Weiss, L.; Rinnerthaler, G.; Greil, R.; Melchardt, T. Prognostic and Predictive Factors in Advanced Head and Neck Squamous Cell Carcinoma. Int. J. Mol. Sci. 2021, 22, 4981. [Google Scholar] [CrossRef]
- Ghantous, Y.; Schussel, J.; Brait, M. Tobacco and alcohol-induced epigenetic changes in oral carcinoma. Curr. Opin. Oncol. 2018, 30, 152–158. [Google Scholar] [CrossRef]
- Li, Y.-C.; Cheng, A.-J.; Lee, L.-Y.; Huang, Y.-C.; Chang, J.T.-C. Multifaceted Mechanisms of Areca Nuts in Oral Carcinogenesis: The Molecular Pathology from Precancerous Condition to Malignant Transformation. J. Cancer 2019, 10, 4054–4062. [Google Scholar] [CrossRef] [Green Version]
- Taberna, M.; Mena, M.; Pavón, M.A.; Alemany, L.; Gillison, M.L.; Mesía, R. Human papillomavirus-related oropharyngeal cancer. Ann. Oncol. 2017, 28, 2386–2398. [Google Scholar] [CrossRef]
- Gupta, A.A.; Kheur, S.; Varadarajan, S.; Parveen, S.; Dewan, H.; Alhazmi, Y.A.; Raj, T.A.; Testaralli, L.; Patil, S. Chronic mechanical irritation and oral squamous cell carcinoma: A systematic review and meta-analysis. Bosn. J. Basic Med. Sci. 2021, 21, 647–658. [Google Scholar] [CrossRef]
- Farquhar, D.R.; Tanner, A.M.; Masood, M.M.; Patel, S.R.; Hackman, T.G.; Olshan, A.F.; Mazul, A.L.; Zevallos, J.P. Oral tongue carcinoma among young patients: An analysis of risk factors and survival. Oral Oncol. 2018, 84, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Lenze, N.R.; Farquhar, D.R.; Dorismond, D.; Sheth, S.; Zevallos, J.P.; Blumberg, J.; Lumley, C.; Patel, S.; Hackman, T.; Weissler, M.C.; et al. Age and risk of recurrence in oral tongue squamous cell carcinoma: Systematic review. Head Neck 2020, 42, 3755–3768. [Google Scholar] [CrossRef] [PubMed]
- Burtness, B.; Harrington, K.J.; Greil, R.; Soulières, D.; Tahara, M.; de Castro, G., Jr.; Psyrri, A.; Basté, N.; Neupane, P.; Bratland, A.; et al. Pembrolizumab alone or with chemotherapy versus cetuximab with chemotherapy for recurrent or metastatic squamous cell carcinoma of the head and neck (KEYNOTE-048): A randomised, open-label, phase 3 study. Lancet 2019, 394, 1915–1928. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, N.; Sento, S.; Yoshizawa, Y.; Sasabe, E.; Kudo, Y.; Yamamoto, T. Current Trends and Future Prospects of Molecular Targeted Therapy in Head and Neck Squamous Cell Carcinoma. Int. J. Mol. Sci. 2020, 22, 240. [Google Scholar] [CrossRef] [PubMed]
- Ferris, R.L. Immunology and Immunotherapy of Head and Neck Cancer. J. Clin. Oncol. 2015, 33, 3293–3304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoenfeld, A.J.; Hellmann, M.D. Acquired Resistance to Immune Checkpoint Inhibitors. Cancer Cell 2020, 37, 443–455. [Google Scholar] [CrossRef]
- Bagchi, S.; Yuan, R.; Engleman, E.G. Immune Checkpoint Inhibitors for the Treatment of Cancer: Clinical Impact and Mechanisms of Response and Resistance. Annu. Rev. Pathol. Mech. Dis. 2021, 16, 223–249. [Google Scholar] [CrossRef] [PubMed]
- Kok, V.C. Current Understanding of the Mechanisms Underlying Immune Evasion From PD-1/PD-L1 Immune Checkpoint Blockade in Head and Neck Cancer. Front. Oncol. 2020, 10, 268. [Google Scholar] [CrossRef] [Green Version]
- Rooney, M.S.; Shukla, S.A.; Wu, C.J.; Getz, G.; Hacohen, N. Molecular and Genetic Properties of Tumors Associated with Local Immune Cytolytic Activity. Cell 2015, 160, 48–61. [Google Scholar] [CrossRef] [Green Version]
- Shukla, S.A.; Rooney, M.S.; Rajasagi, M.; Tiao, G.; Dixon, P.M.; Lawrence, M.S.; Stevens, J.; Lane, W.J.; Dellagatta, J.L.; Steelman, S.; et al. Comprehensive analysis of cancer-associated somatic mutations in class I HLA genes. Nat. Biotechnol. 2015, 33, 1152–1158. [Google Scholar] [CrossRef]
- Del Campo, A.B.; Aptsiauri, N.; Méndez, R.; Zinchenko, S.; Vales, Á.; Paschen, A.; Ward, S.; Ruiz-Cabello, F.; González-Aseguinolaza, G.; Garrido, F. Efficient Recovery of HLA Class I Expression in Human Tumor Cells After Beta2-Microglobulin Gene Transfer Using Adenoviral Vector: Implications for Cancer Immunotherapy. Scand. J. Immunol. 2009, 70, 125–135. [Google Scholar] [CrossRef] [PubMed]
- del Campo, A.B.; Carretero, J.; Muñoz, J.A.; Zinchenko, S.; Ruiz-Cabello, F.; González-Aseguinolaza, G.; Garrido, F.; Aptsiauri, N. Adenovirus expressing β2-microglobulin recovers HLA class I expression and antitumor immunity by increasing T-cell recognition. Cancer Gene Ther. 2014, 21, 317–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, J.S.; Weissfeld, J.L.; Ragin, C.C.; Rossie, K.M.; Martin, C.L.; Shuster, M.; Ishwad, C.S.; Law, J.C.; MyersT, E.N.; Johnson, J.T.; et al. The influence of clinical and demographic risk factors on the establishment of head and neck squamous cell car-cinoma cell lines. Oral Oncol. 2007, 43, 701–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heo, D.S.; Snyderman, C.; Gollin, S.M.; Pan, S.; Walker, E.; Deka, R.; Barnes, E.L.; Johnson, J.T.; Herberman, R.B.; Whiteside, T.L. Biology, cytogenetics, and sensitivity to immunological effector cells of new head and neck squamous cell carcinoma lines. Cancer Res. 1989, 49, 5167–5175. [Google Scholar] [PubMed]
- Zheng, X.; Wu, K.; Liao, S.; Pan, Y.; Sun, Y.; Chen, X.; Zhang, Y.; Xia, S.; Hu, Y.; Zhang, J. MicroRNA-transcription factor network analysis reveals miRNAs cooperatively suppress RORA in oral squamous cell carcinoma. Oncogenesis 2018, 7, 79. [Google Scholar] [CrossRef] [Green Version]
- De Ruiter, E.J.; Mulder, F.J.; Koomen, B.M.; Speel, E.-J.; Hout, M.F.C.M.V.D.; de Roest, R.H.; Bloemena, E.; Devriese, L.A.; Willems, S.M. Comparison of three PD-L1 immunohistochemical assays in head and neck squamous cell carcinoma (HNSCC). Mod. Pathol. 2020, 34, 1125–1132. [Google Scholar] [CrossRef]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Ali, R.; Al Zahrani, H.; Barhoumi, T.; Alhallaj, A.; Mashhour, A.; Alshammari, M.; Alshawakir, Y.; Baz, O.; Alanazi, A.; Khan, A.; et al. Isolation and Establishment of a Highly Proliferative, Cancer Stem Cell-Like, and Naturally Immortalized Triple-Negative Breast Cancer Cell Line, KAIMRC2. Cells 2021, 10, 1303. [Google Scholar] [CrossRef]
- Blanchard, P.; Belkhir, F.; Temam, S.; El Khoury, C.; De Felice, F.; Casiraghi, O.; Patrikidou, A.; Mirghani, H.; Levy, A.; Even, C.; et al. Outcomes and prognostic factors for squamous cell carcinoma of the oral tongue in young adults: A single-institution case-matched analysis. Eur. Arch. Oto-Rhino-Laryngol. 2016, 274, 1683–1690. [Google Scholar] [CrossRef]
- Wang, S.J.; Asthana, S.; van Zante, A.; Heaton, C.M.; Phuchareon, J.; Stein, L.; Higuchi, S.; Kishimoto, T.; Chiu, C.Y.; Olshen, A.B.; et al. Establishment and characterization of an oral tongue squamous cell carcinoma cell line from a never-smoking patient. Oral Oncol. 2017, 69, 1–10. [Google Scholar] [CrossRef]
- Vipparthi, K.; Patel, A.K.; Ghosh, S.; Das, S.; Das, C.; Das, K.; Sarkar, A.; Thatikonda, V.; Pal, B.; Remani, A.S.K.N.; et al. Two novel cell culture models of buccal mucosal oral cancer from patients with no risk-habits of tobacco smoking or chewing. Oral Oncol. 2020, 113, 105131. [Google Scholar] [CrossRef] [PubMed]
- Fasano, M.; Della Corte, C.M.; Di Liello, R.; Viscardi, G.; Sparano, F.; Iacovino, M.L.; Paragliola, F.; Piccolo, A.; Napolitano, S.; Martini, G.; et al. Immunotherapy for head and neck cancer: Present and future. Crit. Rev. Oncol. 2022, 174, 103679. [Google Scholar] [CrossRef] [PubMed]
- Durgeau, A.; Virk, Y.; Corgnac, S.; Mami-Chouaib, F. Recent Advances in Targeting CD8 T-Cell Immunity for More Effective Cancer Immunotherapy. Front. Immunol. 2018, 9, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jhunjhunwala, S.; Hammer, C.; Delamarre, L. Antigen presentation in cancer: Insights into tumour immunogenicity and immune evasion. Nat. Rev. Cancer 2021, 21, 298–312. [Google Scholar] [CrossRef]
- Sade-Feldman, M.; Jiao, Y.J.; Chen, J.H.; Rooney, M.S.; Barzily-Rokni, M.; Eliane, J.-P.; Bjorgaard, S.L.; Hammond, M.R.; Vitzthum, H.; Blackmon, S.M.; et al. Resistance to checkpoint blockade therapy through inactivation of antigen presentation. Nat. Commun. 2017, 8, 1136. [Google Scholar] [CrossRef] [Green Version]
- Wieczorek, M.; Abualrous, E.T.; Sticht, J.; Álvaro-Benito, M.; Stolzenberg, S.; Noé, F.; Freund, C. Major Histocompatibility Complex (MHC) Class I and MHC Class II Proteins: Conformational Plasticity in Antigen Presentation. Front. Immunol. 2017, 8, 292. [Google Scholar] [CrossRef] [Green Version]
- Zaretsky, J.M.; Garcia-Diaz, A.; Shin, D.S.; Escuin-Ordinas, H.; Hugo, W.; Hu-Lieskovan, S.; Torrejon, D.Y.; Abril-Rodriguez, G.; Sandoval, S.; Barthly, L.; et al. Mutations Associated with Acquired Resistance to PD-1 Blockade in Melanoma. N. Engl. J. Med. 2016, 375, 819–829. [Google Scholar] [CrossRef]
- Bernal, M.; Ruiz-Cabello, F.; Concha, A.; Paschen, A.; Garrido, F. Implication of the β2-microglobulin gene in the generation of tumor escape phenotypes. Cancer Immunol. Immunother. 2012, 61, 1359–1371. [Google Scholar] [CrossRef]
- Maleno, I.; Aptsiauri, N.; Cabrera, T.; Gallego, A.; Paschen, A.; López-Nevot, M.A.; Garrido, F. Frequent loss of heterozygosity in the β2-microglobulin region of chromosome 15 in primary human tumors. Immunogenetics 2010, 63, 65–71. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, W.; Zhang, W.; Wu, S.; Yan, Z. Candida albicans induces upregulation of programmed death ligand 1 in oral squamous cell carcinoma. J. Oral Pathol. Med. 2022, 51, 444–453. [Google Scholar] [CrossRef]
- Lanzel, E.A.; Paula Gomez Hernandez, M.; Bates, A.M.; Treinen, C.N.; Starman, E.E.; Fischer, C.L.; Parashar, D.; Guthmiller, J.M.; Johnson, G.K.; Abbasi, T.; et al. Predicting PD-L1 expression on human cancer cells using next-generation sequencing information in computational simulation models. Cancer Immunol. Immunother. 2016, 65, 1511–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, S.; Itai, S.; Kaneko, M.K.; Kato, Y. Detection of high PD-L1 expression in oral cancers by a novel monoclonal antibody L1Mab-4. Biochem. Biophys. Rep. 2018, 13, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Philip, M.; Schietinger, A. CD8+ T cell differentiation and dysfunction in cancer. Nat. Rev. Immunol. 2021, 22, 209–223. [Google Scholar] [CrossRef] [PubMed]
- Rossjohn, J.; Gras, S.; Miles, J.J.; Turner, S.J.; Godfrey, D.I.; McCluskey, J. T Cell Antigen Receptor Recognition of Antigen-Presenting Molecules. Annu. Rev. Immunol. 2015, 33, 169–200. [Google Scholar] [CrossRef]
- Van Kaer, L.; Parekh, V.V.; Postoak, J.L.; Wu, L. Role of autophagy in MHC class I-restricted antigen presentation. Mol. Immunol. 2017, 113, 2–5. [Google Scholar] [CrossRef]
- Hazini, A.; Fisher, K.; Seymour, L. Deregulation of HLA-I in cancer and its central importance for immunotherapy. J. Immunother. Cancer 2021, 9, e002899. [Google Scholar] [CrossRef]
- Kawazu, M.; Ueno, T.; Saeki, K.; Sax, N.; Togashi, Y.; Kanaseki, T.; Chida, K.; Kishigami, F.; Sato, K.; Kojima, S.; et al. HLA Class I Analysis Provides Insight Into the Genetic and Epigenetic Background of Immune Evasion in Colorectal Cancer With High Microsatellite Instability. Gastroenterology 2022, 162, 799–812. [Google Scholar] [CrossRef]
- Gettinger, S.; Choi, J.; Hastings, K.; Truini, A.; Datar, I.; Sowell, R.; Wurtz, A.; Dong, W.; Cai, G.; Melnick, M.A.; et al. Impaired HLA Class I Antigen Processing and Presentation as a Mechanism of Acquired Resistance to Immune Checkpoint Inhibitors in Lung Cancer. Cancer Discov. 2017, 7, 1420–1435. [Google Scholar] [CrossRef] [Green Version]
- Pereira, C.; Gimenez-Xavier, P.; Pros, E.; Pajares, M.J.; Moro, M.; Gomez, A.; Navarro, A.; Condom, E.; Moran, S.; Gomez-Lopez, G.; et al. Genomic Profiling of Patient-Derived Xenografts for Lung Cancer Identifies B2M Inactivation Impairing Immunorecognition. Clin. Cancer Res. 2017, 23, 3203–3213. [Google Scholar] [CrossRef] [Green Version]
- Snahnicanova, Z.; Kasubova, I.; Kalman, M.; Grendar, M.; Mikolajcik, P.; Gabonova, E.; Laca, L.; Caprnda, M.; Rodrigo, L.; Ciccocioppo, R.; et al. Genetic and epigenetic analysis of the beta-2-microglobulin gene in microsatellite instable colorectal cancer. Clin. Exp. Med. 2019, 20, 87–95. [Google Scholar] [CrossRef]
- Karabekian, Z.; Idrees, S.; Ding, H.; Jamshidi, A.; Posnack, N.; Sarvazyan, N. Downregulation of beta-microglobulin to diminish T-lymphocyte lysis of non-syngeneic cell sources of engineered heart tissue constructs. Biomed. Mater. 2015, 10, 034101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thongsin, N.; Wattanapanitch, M. CRISPR/Cas9 Ribonucleoprotein Complex-Mediated Efficient B2M Knockout in Human Induced Pluripotent Stem Cells (iPSCs). In Induced Pluripotent Stem (iPS) Cells. Methods in Molecular Biology; Nagy, A., Turksen, K., Eds.; Humana: New York, NY, USA, 2021; Volume 2454, pp. 607–624. [Google Scholar] [CrossRef]
- Couture, A.; Garnier, A.; Docagne, F.; Boyer, O.; Vivien, D.; Le-Mauff, B.; Latouche, J.-B.; Toutirais, O. HLA-Class II Artificial Antigen Presenting Cells in CD4+ T Cell-Based Immunotherapy. Front. Immunol. 2019, 10, 1081. [Google Scholar] [CrossRef] [PubMed]
- Axelrod, M.L.; Cook, R.S.; Johnson, D.B.; Balko, J.M. Biological Consequences of MHC-II Expression by Tumor Cells in Cancer. Clin. Cancer Res. 2019, 25, 2392–2402. [Google Scholar] [CrossRef] [PubMed]
- Kreiter, S.; Vormehr, M.; Van De Roemer, N.; Diken, M.; Löwer, M.; Diekmann, J.; Boegel, S.; Schrörs, B.; Vascotto, F.; Castle, J.C.; et al. Mutant MHC class II epitopes drive therapeutic immune responses to cancer. Nature 2015, 520, 692–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeves, E.; James, E. Antigen processing and immune regulation in the response to tumours. Immunology 2017, 150, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Garrido, F.; Aptsiauri, N.; Doorduijn, E.M.; Lora, A.M.G.; van Hall, T. The urgent need to recover MHC class I in cancers for effective immunotherapy. Curr. Opin. Immunol. 2016, 39, 44–51. [Google Scholar] [CrossRef] [Green Version]
- Ni, J.; Miller, M.; Stojanovic, A.; Garbi, N.; Cerwenka, A. Sustained effector function of IL-12/15/18–preactivated NK cells against established tumors. J. Exp. Med. 2012, 209, 2351–2365. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, X.; Sun, Y.; Li, Y.; Ma, J.; Lv, Y.; Hu, Y.; Zhou, Y.; Zhang, J. A Novel Tongue Squamous Cell Carcinoma Cell Line Escapes from Immune Recognition due to Genetic Alterations in HLA Class I Complex. Cells 2023, 12, 35. https://doi.org/10.3390/cells12010035
Zheng X, Sun Y, Li Y, Ma J, Lv Y, Hu Y, Zhou Y, Zhang J. A Novel Tongue Squamous Cell Carcinoma Cell Line Escapes from Immune Recognition due to Genetic Alterations in HLA Class I Complex. Cells. 2023; 12(1):35. https://doi.org/10.3390/cells12010035
Chicago/Turabian StyleZheng, Xiaofeng, Yanan Sun, Yiwei Li, Jiyuan Ma, Yinan Lv, Yaying Hu, Yi Zhou, and Jiali Zhang. 2023. "A Novel Tongue Squamous Cell Carcinoma Cell Line Escapes from Immune Recognition due to Genetic Alterations in HLA Class I Complex" Cells 12, no. 1: 35. https://doi.org/10.3390/cells12010035