
Basket to Purkinje Cell Inhibitory Ephaptic Coupling Is Abolished in Episodic Ataxia Type 1

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cerebellar Preparation
2.2. Cerebellar Cortex Recordings
2.3. Pinceau Imaging and Immunofluorescence
2.4. Statistical Analysis
3. Results
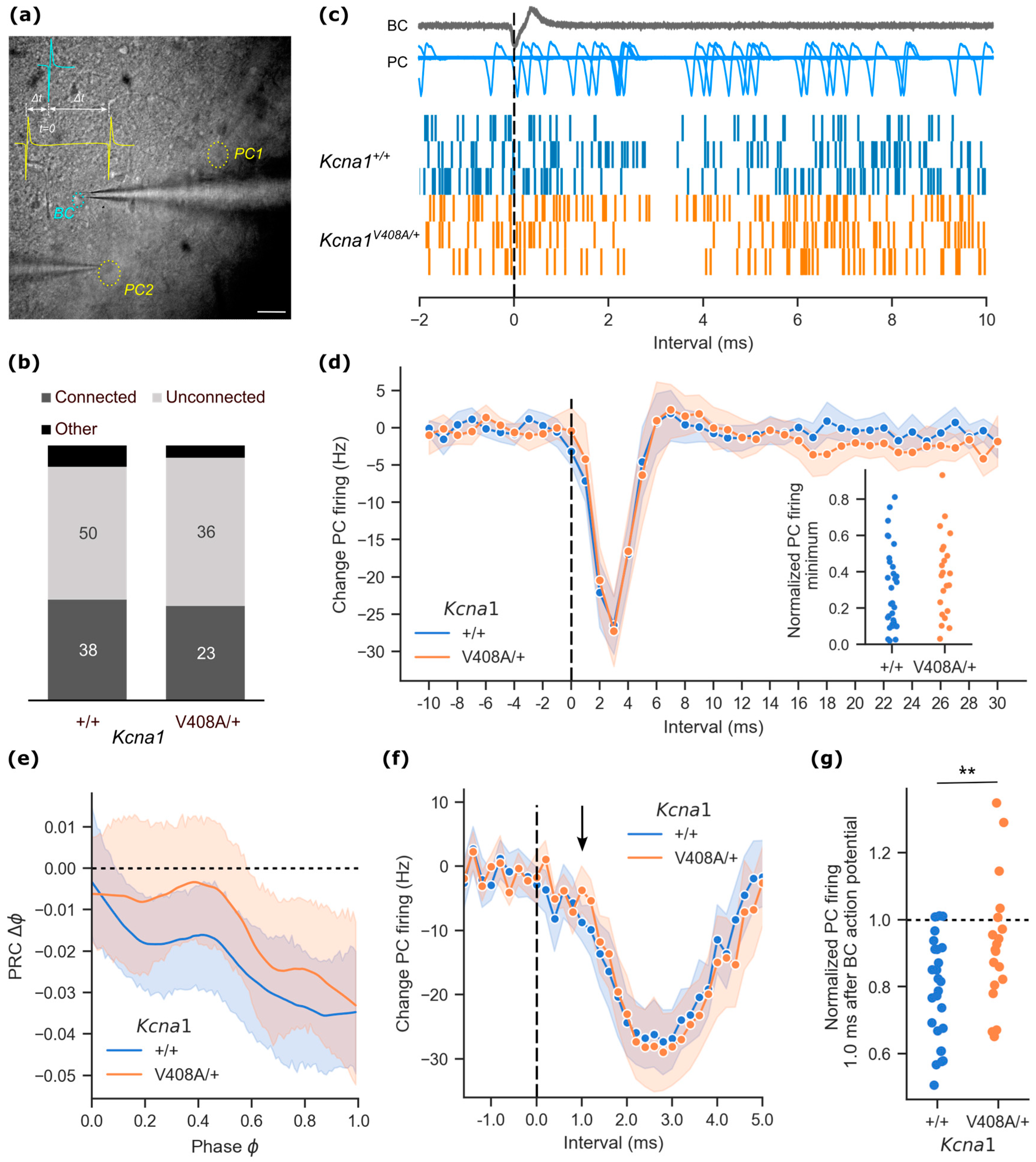
3.1. Impaired Ultra-Rapid Basket Cell Mediated Inhibition of Spontaneous Purkinje Cell Activity in EA1 Mutant Mice
3.2. Cerebellar Pinceaux Are Enlarged in EA1 Mutant Mice
3.3. Pinceau Visualization Correlates with Ephaptic Coupling
3.4. Evoked Basket Cell Firing Reveals Loss of Picrotoxin-Resistant Inhibition
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Browne, D.L.; Gancher, S.T.; Nutt, J.G.; Brunt, E.R.; Smith, E.A.; Kramer, P.; Litt, M. Episodic Ataxia/Myokymia Syndrome Is Associated with Point Mutations in the Human Potassium Channel Gene, KCNA1. Nat. Genet. 1994, 8, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Kunkel, D.D.; Martin, T.M.; Schwartzkroin, P.A.; Tempel, B.L. Heteromultimeric K+ Channels in Terminal and Juxtaparanodal Regions of Neurons. Nature 1993, 365, 75–79. [Google Scholar] [CrossRef]
- Laube, G.; Röper, J.; Pitt, J.C.; Sewing, S.; Kistner, U.; Garner, C.C.; Pongs, O.; Veh, R.W. Ultrastructural Localization of Shaker-Related Potassium Channel Subunits and Synapse-Associated Protein 90 to Septate-like Junctions in Rat Cerebellar Pinceaux. Brain Res. Mol. Brain Res. 1996, 42, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Kunkel, D.D.; Schwartzkroin, P.A.; Tempel, B.L. Localization of Kv1.1 and Kv1.2, Two K Channel Proteins, to Synaptic Terminals, Somata, and Dendrites in the Mouse Brain. J. Neurosci. 1994, 14, 4588–4599. [Google Scholar] [CrossRef] [PubMed]
- Begum, R.; Bakiri, Y.; Volynski, K.E.; Kullmann, D.M. Action Potential Broadening in a Presynaptic Channelopathy. Nat. Commun. 2016, 7, 12102. [Google Scholar] [CrossRef] [PubMed]
- Southan, A.P.; Robertson, B. Patch-Clamp Recordings from Cerebellar Basket Cell Bodies and Their Presynaptic Terminals Reveal an Asymmetric Distribution of Voltage-Gated Potassium Channels. J. Neurosci. 1998, 18, 948–955. [Google Scholar] [CrossRef]
- Herson, P.S.; Virk, M.; Rustay, N.R.; Bond, C.T.; Crabbe, J.C.; Adelman, J.P.; Maylie, J. A Mouse Model of Episodic Ataxia Type-1. Nat. Neurosci. 2003, 6, 378–383. [Google Scholar] [CrossRef] [PubMed]
- Adelman, J.P.; Bond, C.T.; Pessia, M.; Maylie, J. Episodic Ataxia Results from Voltage-Dependent Potassium Channels with Altered Functions. Neuron 1995, 15, 1449–1454. [Google Scholar] [CrossRef] [PubMed]
- Iwakura, A.; Uchigashima, M.; Miyazaki, T.; Yamasaki, M.; Watanabe, M. Lack of Molecular-Anatomical Evidence for GABAergic Influence on Axon Initial Segment of Cerebellar Purkinje Cells by the Pinceau Formation. J. Neurosci. 2012, 32, 9438–9448. [Google Scholar] [CrossRef]
- Sotelo, C.; Llinás, R. Specialized Membrane Junctions between Neurons in the Vertebrate Cerebellar Cortex. J. Cell Biol. 1972, 53, 271–289. [Google Scholar] [CrossRef] [PubMed]
- Blot, A.; Barbour, B. Ultra-Rapid Axon-Axon Ephaptic Inhibition of Cerebellar Purkinje Cells by the Pinceau. Nat. Neurosci. 2014, 17, 289–295. [Google Scholar] [CrossRef]
- Korn, H.; Axelrad, H. Electrical Inhibition of Purkinje Cells in the Cerebellum of the Rat. Proc. Natl. Acad. Sci. USA 1980, 77, 6244–6247. [Google Scholar] [CrossRef] [PubMed]
- Blot, A.; Barbour, B. Analysis of the Study of the Cerebellar Pinceau by Korn and Axelrad. bioRxiv 2013, 001123. [Google Scholar] [CrossRef]
- Kole, M.J.; Qian, J.; Waase, M.P.; Klassen, T.L.; Chen, T.T.; Augustine, G.J.; Noebels, J.L. Selective Loss of Presynaptic Potassium Channel Clusters at the Cerebellar Basket Cell Terminal Pinceau in Adam11 Mutants Reveals Their Role in Ephaptic Control of Purkinje Cell Firing. J. Neurosci. 2015, 35, 11433–11444. [Google Scholar] [CrossRef]
- Klein, S.; Seidler, B.; Kettenberger, A.; Sibaev, A.; Rohn, M.; Feil, R.; Allescher, H.-D.; Vanderwinden, J.-M.; Hofmann, F.; Schemann, M.; et al. Interstitial Cells of Cajal Integrate Excitatory and Inhibitory Neurotransmission with Intestinal Slow-Wave Activity. Nat. Commun. 2013, 4, 1630. [Google Scholar] [CrossRef] [PubMed]
- Amat, S.B.; Rowan, M.J.M.; Gaffield, M.A.; Bonnan, A.; Kikuchi, C.; Taniguchi, H.; Christie, J.M. Using C-Kit to Genetically Target Cerebellar Molecular Layer Interneurons in Adult Mice. PLoS ONE 2017, 12, e0179347. [Google Scholar] [CrossRef]
- Ting, J.T.; Daigle, T.L.; Chen, Q.; Feng, G. Acute Brain Slice Methods for Adult and Aging Animals: Application of Targeted Patch Clamp Analysis and Optogenetics. In Patch-Clamp Methods and Protocols; Martina, M., Taverna, S., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2014; pp. 221–242. ISBN 978-1-4939-1096-0. [Google Scholar]
- Perkins, K.L. Cell-Attached Voltage-Clamp and Current-Clamp Recording and Stimulation Techniques in Brain Slices. J. Neurosci. Methods 2006, 154, 1–18. [Google Scholar] [CrossRef]
- Phoka, E.; Cuntz, H.; Roth, A.; Häusser, M. A New Approach for Determining Phase Response Curves Reveals That Purkinje Cells Can Act as Perfect Integrators. PLoS Comput. Biol. 2010, 6, e1000768. [Google Scholar] [CrossRef] [PubMed]
- Häusser, M.; Clark, B.A. Tonic Synaptic Inhibition Modulates Neuronal Output Pattern and Spatiotemporal Synaptic Integration. Neuron 1997, 19, 665–678. [Google Scholar] [CrossRef]
- Zerr, P.; Adelman, J.P.; Maylie, J. Episodic Ataxia Mutations in Kv1.1 Alter Potassium Channel Function by Dominant Negative Effects or Haploinsufficiency. J. Neurosci. 1998, 18, 2842–2848. [Google Scholar] [CrossRef]
- Bishop, G.A. An Analysis of HRP-Filled Basket Cell Axons in the Cat’s Cerebellum. Anat. Embryol. 1993, 188, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Eccles, J.C.; Llinás, R.; Sasaki, K. The Inhibitory Interneurones within the Cerebellar Cortex. Exp. Brain Res. 1966, 1, 1–16. [Google Scholar] [CrossRef]
- Todorov, B.; Kros, L.; Shyti, R.; Plak, P.; Haasdijk, E.D.; Raike, R.S.; Frants, R.R.; Hess, E.J.; Hoebeek, F.E.; De Zeeuw, C.I.; et al. Purkinje Cell-Specific Ablation of CaV2.1 Channels Is Sufficient to Cause Cerebellar Ataxia in Mice. Cerebellum 2012, 11, 246–258. [Google Scholar] [CrossRef]
- Du, X.; Wang, J.; Zhu, H.; Rinaldo, L.; Lamar, K.-M.; Palmenberg, A.C.; Hansel, C.; Gomez, C.M. Second Cistron in CACNA1A Gene Encodes a Transcription Factor Mediating Cerebellar Development and SCA6. Cell 2013, 154, 118–133. [Google Scholar] [CrossRef] [PubMed]
- Irie, T.; Matsuzaki, Y.; Sekino, Y.; Hirai, H. Kv3.3 Channels Harbouring a Mutation of Spinocerebellar Ataxia Type 13 Alter Excitability and Induce Cell Death in Cultured Cerebellar Purkinje Cells. J. Physiol. 2014, 592, 229–247. [Google Scholar] [CrossRef]
- Yan, H.; Pablo, J.L.; Pitt, G.S. FGF14 Regulates Presynaptic Ca2+ Channels and Synaptic Transmission. Cell Rep. 2013, 4, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Vivekananda, U.; Novak, P.; Bello, O.D.; Korchev, Y.E.; Krishnakumar, S.S.; Volynski, K.E.; Kullmann, D.M. Kv1.1 Channelopathy Abolishes Presynaptic Spike Width Modulation by Subthreshold Somatic Depolarization. Proc. Natl. Acad. Sci. USA 2017, 114, 2395–2400. [Google Scholar] [CrossRef]
- Zang, Y.; Dieudonné, S.; De Schutter, E. Voltage- and Branch-Specific Climbing Fiber Responses in Purkinje Cells. Cell Rep. 2018, 24, 1536–1549. [Google Scholar] [CrossRef]
- Buttermore, E.D.; Piochon, C.; Wallace, M.L.; Philpot, B.D.; Hansel, C.; Bhat, M.A. Pinceau Organization in the Cerebellum Requires Distinct Functions of Neurofascin in Purkinje and Basket Neurons during Postnatal Development. J. Neurosci. 2012, 32, 4724–4742. [Google Scholar] [CrossRef]
- Couto, J.; Linaro, D.; Schutter, E.D.; Giugliano, M. On the Firing Rate Dependency of the Phase Response Curve of Rat Purkinje Neurons In Vitro. PLoS Comput. Biol. 2015, 11, e1004112. [Google Scholar] [CrossRef]
- Zang, Y.; Hong, S.; De Schutter, E. Firing Rate-Dependent Phase Responses of Purkinje Cells Support Transient Oscillations. eLife 2020, 9, e60692. [Google Scholar] [CrossRef] [PubMed]
- Somogyi, P.; Hámori, J. A Quantitative Electron Microscopic Study of the Purkinje Cell Axon Initial Segment. Neuroscience 1976, 1, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Connor, J.X.; McCormack, K.; Pletsch, A.; Gaeta, S.; Ganetzky, B.; Chiu, S.-Y.; Messing, A. Genetic Modifiers of the Kvβ2-Null Phenotype in Mice. Genes Brain Behav. 2005, 4, 77–88. [Google Scholar] [CrossRef]
- Zhou, H.; Lin, Z.; Voges, K.; Ju, C.; Gao, Z.; Bosman, L.W.; Ruigrok, T.J.; Hoebeek, F.E.; De Zeeuw, C.I.; Schonewille, M. Cerebellar Modules Operate at Different Frequencies. eLife 2014, 3, e02536. [Google Scholar] [CrossRef]
- Zhou, J.; Brown, A.M.; Lackey, E.P.; Arancillo, M.; Lin, T.; Sillitoe, R.V. Purkinje Cell Neurotransmission Patterns Cerebellar Basket Cells into Zonal Modules Defined by Distinct Pinceau Sizes. eLife 2020, 9, e55569. [Google Scholar] [CrossRef]
- Kozareva, V.; Martin, C.; Osorno, T.; Rudolph, S.; Guo, C.; Vanderburg, C.; Nadaf, N.; Regev, A.; Regehr, W.G.; Macosko, E. A Transcriptomic Atlas of Mouse Cerebellar Cortex Comprehensively Defines Cell Types. Nature 2021, 598, 214–219. [Google Scholar] [CrossRef]
- Kim, J.; Lee, S.; Tsuda, S.; Zhang, X.; Asrican, B.; Gloss, B.; Feng, G.; Augustine, G.J. Optogenetic Mapping of Cerebellar Inhibitory Circuitry Reveals Spatially Biased Coordination of Interneurons via Electrical Synapses. Cell Rep. 2014, 7, 1601–1613. [Google Scholar] [CrossRef]
- Han, K.-S.; Guo, C.; Chen, C.H.; Witter, L.; Osorno, T.; Regehr, W.G. Ephaptic Coupling Promotes Synchronous Firing of Cerebellar Purkinje Cells. Neuron 2018, 100, 564–578.e3. [Google Scholar] [CrossRef] [PubMed]
- Han, K.-S.; Chen, C.H.; Khan, M.M.; Guo, C.; Regehr, W.G. Climbing Fiber Synapses Rapidly and Transiently Inhibit Neighboring Purkinje Cells via Ephaptic Coupling. Nat. Neurosci. 2020, 23, 1399–1409. [Google Scholar] [CrossRef]
- Anastassiou, C.A.; Koch, C. Ephaptic Coupling to Endogenous Electric Field Activity: Why Bother? Curr. Opin. Neurobiol. 2015, 31, 95–103. [Google Scholar] [CrossRef]
- Rajakulendran, S.; Schorge, S.; Kullmann, D.M.; Hanna, M.G. Episodic Ataxia Type 1: A Neuronal Potassium Channelopathy. Neurotherapeutics 2007, 4, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Mittmann, W.; Koch, U.; Häusser, M. Feed-Forward Inhibition Shapes the Spike Output of Cerebellar Purkinje Cells. J. Physiol. 2005, 563, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Zang, Y.; Schutter, E.D. The Cellular Electrophysiological Properties Underlying Multiplexed Coding in Purkinje Cells. J. Neurosci. 2021, 41, 1850–1863. [Google Scholar] [CrossRef] [PubMed]
- Andersen, P.; Eccles, J.C.; Voorhoeve, P.E. Postsynaptic Inhibition of Cerebellar Purkinje Cells. J. Neurophysiol. 1964, 27, 1138–1153. [Google Scholar] [CrossRef] [PubMed]
- Blot, A.; de Solages, C.; Ostojic, S.; Szapiro, G.; Hakim, V.; Léna, C. Time-Invariant Feed-Forward Inhibition of Purkinje Cells in the Cerebellar Cortex in Vivo. J. Physiol. 2016, 594, 2729–2749. [Google Scholar] [CrossRef] [PubMed]
- Walter, J.T.; Alviña, K.; Womack, M.D.; Chevez, C.; Khodakhah, K. Decreases in the Precision of Purkinje Cell Pacemaking Cause Cerebellar Dysfunction and Ataxia. Nat. Neurosci. 2006, 9, 389–397. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Epitope | Source | Number | Species | IF dilution |
|---|---|---|---|---|
| KV1.1 | NeuroMab | K36/15 | Monoclonal | 1:1000 |
| KV1.1 | Alomone labs | APC-161 | Rabbit | 1:500 |
| Calbindin | Swant | D-28k | Monoclonal | 1:1000 |
| Parvalbumin | Sigma | P3088 | Monoclonal | 1:1000 |
| PSD-95 | Abcam | AB2723 | Monoclonal | 1:1000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin, H.G.S.; Kullmann, D.M. Basket to Purkinje Cell Inhibitory Ephaptic Coupling Is Abolished in Episodic Ataxia Type 1. Cells 2023, 12, 1382. https://doi.org/10.3390/cells12101382
Martin HGS, Kullmann DM. Basket to Purkinje Cell Inhibitory Ephaptic Coupling Is Abolished in Episodic Ataxia Type 1. Cells. 2023; 12(10):1382. https://doi.org/10.3390/cells12101382
Chicago/Turabian StyleMartin, Henry G. S., and Dimitri M. Kullmann. 2023. "Basket to Purkinje Cell Inhibitory Ephaptic Coupling Is Abolished in Episodic Ataxia Type 1" Cells 12, no. 10: 1382. https://doi.org/10.3390/cells12101382
APA StyleMartin, H. G. S., & Kullmann, D. M. (2023). Basket to Purkinje Cell Inhibitory Ephaptic Coupling Is Abolished in Episodic Ataxia Type 1. Cells, 12(10), 1382. https://doi.org/10.3390/cells12101382