

The Mechanoreception in Drosophila melanogaster Oocyte under Modeling Micro- and Hypergravity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
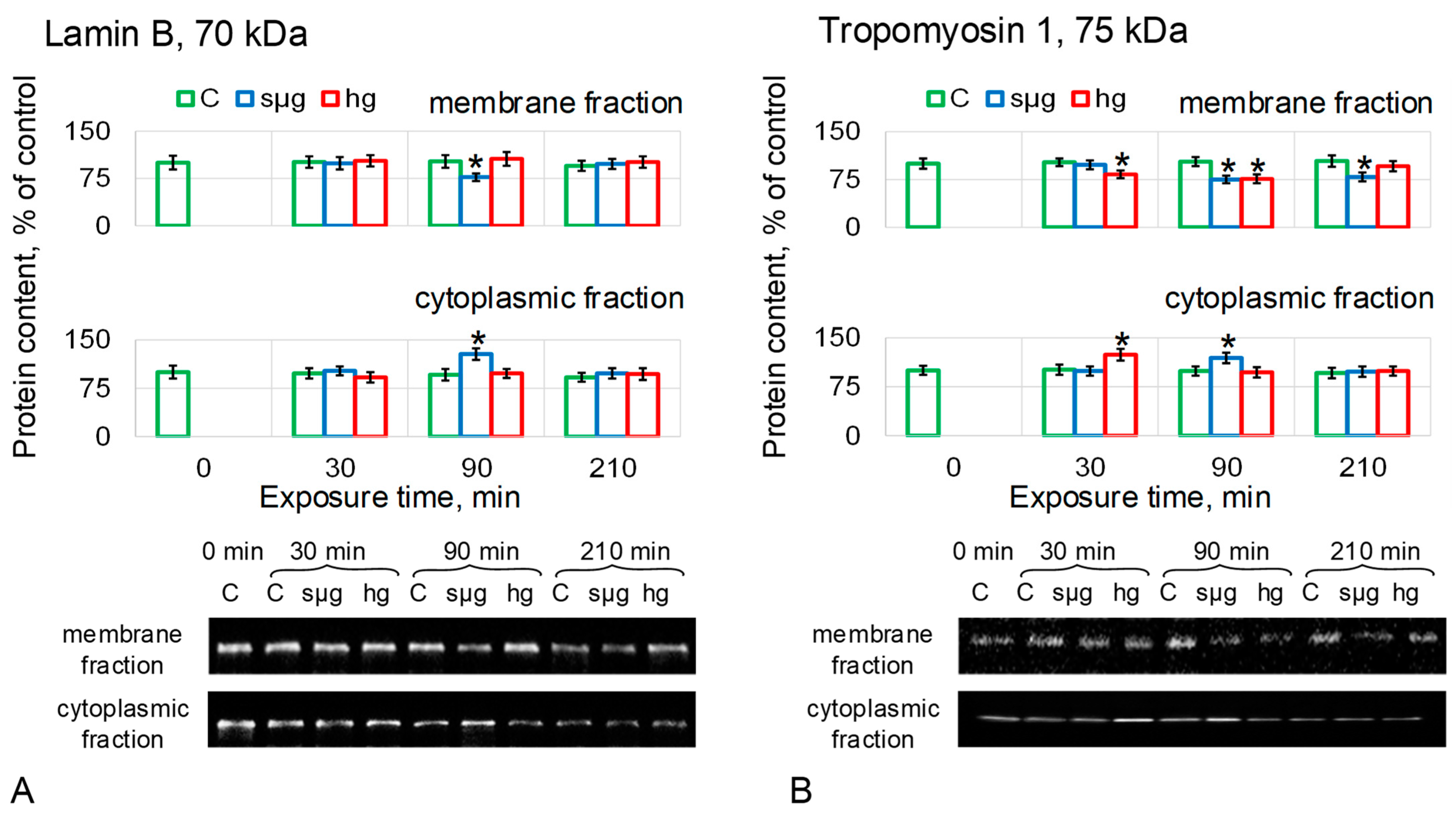{kind=link}
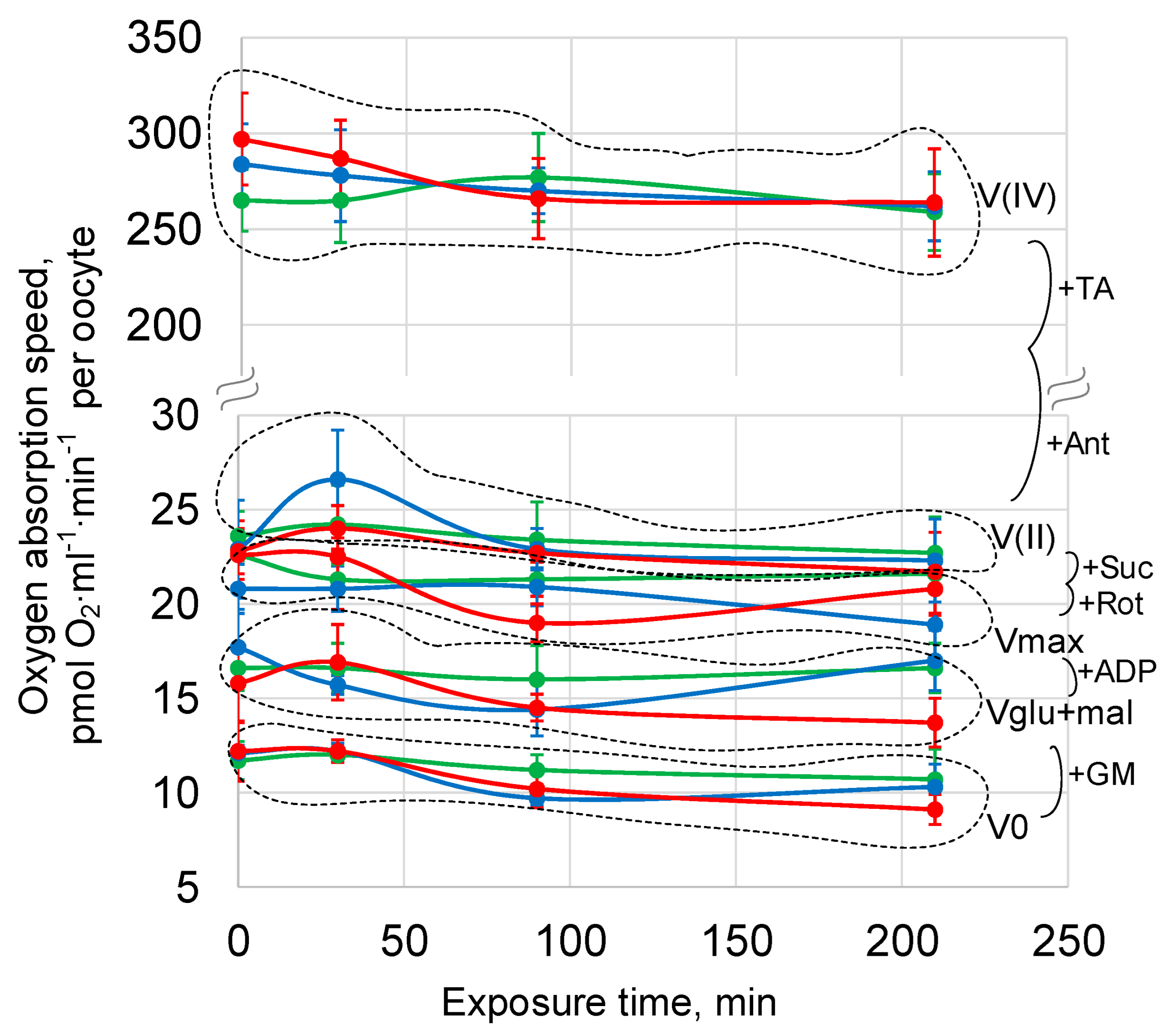{kind=link}
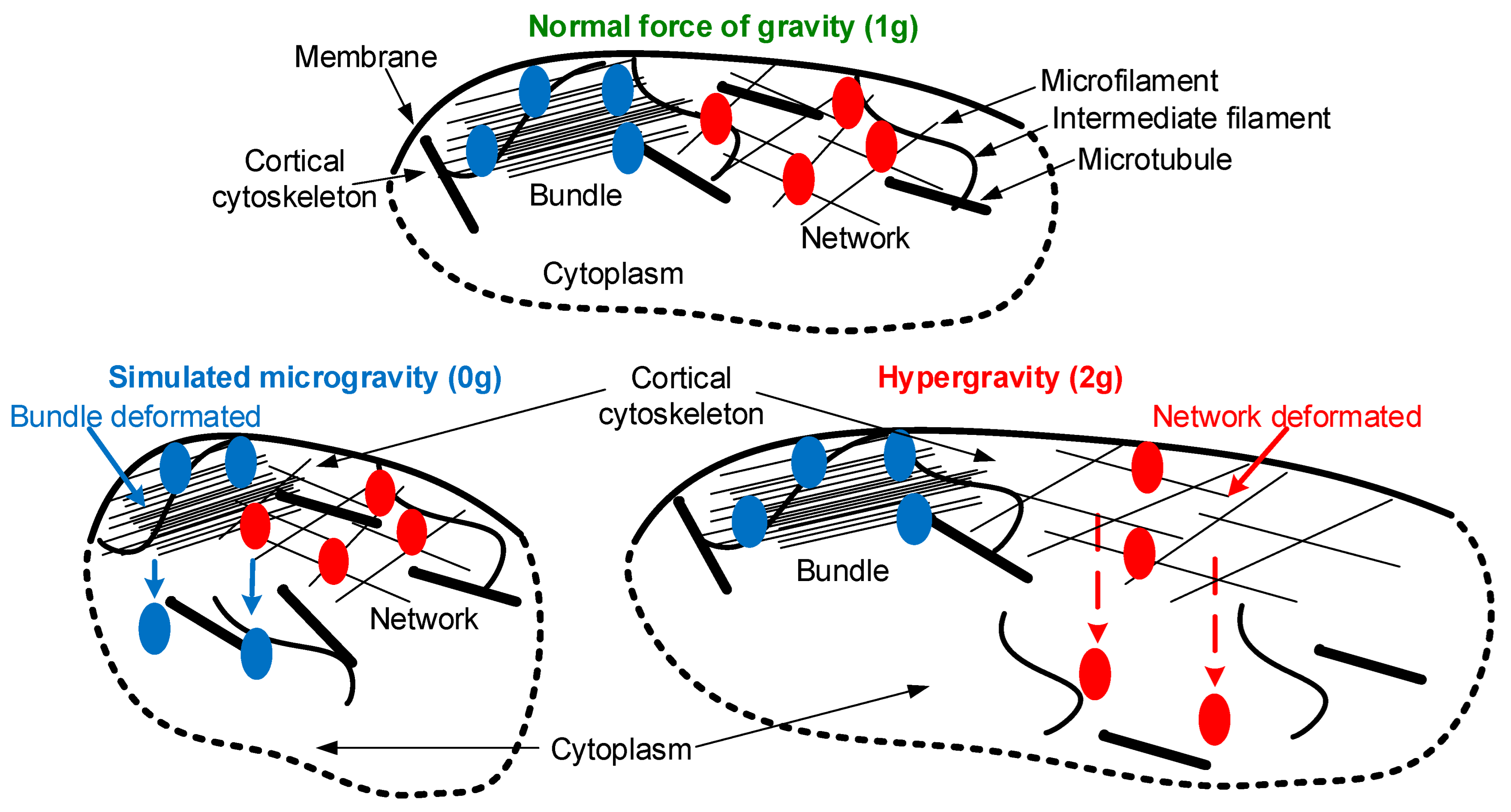{kind=link}
Abstract
Share and Cite
Ogneva, I.V. The Mechanoreception in Drosophila melanogaster Oocyte under Modeling Micro- and Hypergravity. Cells 2023, 12, 1819. https://doi.org/10.3390/cells12141819
Ogneva IV. The Mechanoreception in Drosophila melanogaster Oocyte under Modeling Micro- and Hypergravity. Cells. 2023; 12(14):1819. https://doi.org/10.3390/cells12141819
Chicago/Turabian StyleOgneva, Irina V. 2023. "The Mechanoreception in Drosophila melanogaster Oocyte under Modeling Micro- and Hypergravity" Cells 12, no. 14: 1819. https://doi.org/10.3390/cells12141819
APA StyleOgneva, I. V. (2023). The Mechanoreception in Drosophila melanogaster Oocyte under Modeling Micro- and Hypergravity. Cells, 12(14), 1819. https://doi.org/10.3390/cells12141819