1. Introduction
Plants in their environment are continuously subjected to biotic and abiotic stresses, which limit plant growth and crop yield [
1,
2]. Therefore, plants have evolved various physiological and biochemical mechanisms for stress adaptation that rely mainly on changes in gene expression patterns [
3,
4]. In this process, transcription factors (TFs) represent the master switches that target stress-responsive genes and regulate their expression [
5]. The calmodulin-binding transcription activators (CAMTAs) family is one of the well-characterized calmodulin (CaM)-binding TFs that work downstream of the calcium signaling pathway to elicit plant responses to various biotic and abiotic stresses [
6,
7]. CAMTAs have been identified in plants, humans, Drosophila as well as worms and are evolutionarily conserved from plants to humans [
8,
9,
10,
11,
12,
13]. A typical CAMTA protein has various ordered functional domains with amino acid sequence stretches that are evolutionarily conserved [
8,
14,
15]. These domains include a DNA-binding domain (CG-1), a non-specific DNA-binding domain, a Transcription-associated Immuno Globulin domain (TIG), ankyrin repeats (ANK) for protein–protein interactions, followed by five tandem repeats of a Ca
2+-independent CaM-binding domain (IQ), and a Ca
2+-dependent CaM-binding domain (CaMBD).
In
Arabidopsis thaliana (referred to as
Arabidopsis henceforth), there are six CAMTAs (AtCAMTA1 to AtCAMTA6, referred to as CAMTA1
to CAMTA6 henceforth), which are differentially expressed in response to multiple stresses [
6,
10,
16]. Among these, CAMTA3 (also known as signal-responsive1, SR1), is the most studied member and regulates diverse biotic and abiotic stress responses, as well as thigmomorphogenesis [
8,
17,
18,
19,
20,
21,
22]. Several transcriptomic studies comparing WT and
camta3 mutants revealed a dual regulatory role for CAMTA3 in multiple cellular pathways [
23,
24,
25]. Mutants of
camta3 exhibit up-regulation of plant immune system, salicylic acid (SA) biosynthesis, and salt stress-responsive genes [
17,
23,
25], while genes involved in insect tolerance and low temperature stress response showed down-regulation [
25,
26]. In line with this,
camta3 mutants exhibited autoimmune phenotypes including stunted growth, chlorosis and leaf lesions, SA accumulation, constitutive expression of defense genes and enhanced resistance against bacterial and fungal pathogens [
14,
17,
23,
27]. Our studies have shown that CAMTA3 functions as a negative regulator for salt tolerance in
Arabidopsis [
25]. On the other hand, CAMTA3 acts as a positive regulator for cold and drought stress response as well as defense against insect herbivory and wound-induced response [
20,
28,
29]. CAMTA3 works in concert with CAMTA1, CAMTA2 and CAMTA5 to increase plant freezing tolerance by inducing the expression of the
CRT/DRE-Binding Factor (CBF) genes and other cold-induced genes [
24,
26,
30]. The
camta3 mutants were also more susceptible to herbivore attack and showed decreased levels of glucosinolates, as compared to the wild-type (WT) plants (Laluk et al., 2012). Moreover, CAMTA3 has been recently reported to be a positive regulator for the suppression of plant viral infection by activating the RNAi defense machinery [
22]. Therefore, CAMTA3 functions both as a transcriptional repressor for defense- and salt stress-related genes as well as a transcriptional activator for genes involved in cold, drought, glucosinolate metabolism, herbivory, wound-induced and general stress-mediated responses [
25,
26,
28,
29,
31].
The CAMTA proteins have a CG-1 domain, which binds to
CGCG or
CGTG core motifs with the
CGCG-core consensus motif (
A/
C)
CGCG(
C/
G/
T), whereas the
CGTG-core consensus motif is (
A/
C)
CGTGT [
10,
17,
23,
26,
32]. Furthermore, CAMTA3 has been shown to act as a transcriptional activator of early general stress response genes by binding to the
Rapid Stress Response Element (
RSRE) [
33], which is
CGCGTT motif [
31,
34]. Previous studies have shown that CAMTA3 regulates plant immunity by modulating the expression of the defense genes
EDS1,
NDR1,
EIN3 and
NPRI by binding to the
CGCG elements in their promoters [
17,
19,
27]. It is thought that CAMTA3 regulates plant immunity by regulating the expression of defense genes. However, Lolle et al. [
35] have shown that the enhanced defense phenotype observed in
camta3 is primarily because of the activation of the NLRs (
Nucleotide-binding,
Leucine-rich
Repeat Receptors) - DOMINANT SUPPRESSOR OF camta3 NUMBER 1 (DSC1) and DOMINANT SUPPRESSOR OF camta3 NUMBER (2 DSC2), rather than the loss of the transcriptional activity of CAMTA3 as a negative regulator of gene expression. In a genetic screen, they identified two dominant negative NLRs,
DSC1-DN and
DSC2-DN, that suppressed the autoimmune phenotype of the
camta3 mutant [
35]. The expression of
DSC1-DN or
DSC2-DN in
camta3 mutants suppressed the autoimmune phenotype, restored the expression of the defense genes (
PR1 and
EDS1) to WT levels and repressed plant resistance to fungal and bacterial pathogens [
35]. Furthermore, the expression of
DSC1 or
DSC2 in
Nicotiana benthamiana triggered the HR cell death response, but co-expression of
CAMTA3 prevented this [
35]. Furthermore, BiFC and FRET assays showed CAMTA3 and DSCs to be a part of a nuclear-localized complex [
35]. Together these results led to the suggestion that DSC1/DSC2 and CAMTA3 form a guard–guardee complex, similar to that observed with RESISTANCE TO
P. SYRINGAE PV MACULICOLA 1 (RPM1) and RESISTANT TO
P. SYRINGAE 2 (RPS2) NLRs and RPM1-INTERACTING PROTEIN 4 (RIN4) [
36]. These results, further suggest that modification of CAMTA3 by effectors or loss of CAMTA3 triggers the corresponding NLR guards to activate plant defense genes [
35]. Here, we tested if the DNA-binding and transcriptional activity of CAMTA3 is necessary for its function in plant immunity. We generated point mutations in the conserved amino acids of the CAMTA3 CG-1 domain, evaluated the DNA-binding and transcriptional activity of the mutated versions, and tested their ability to complement the
camta3 mutant phenotype. Our results show that the CAMTA3 DNA-binding activity of the CG-1 and its modulation of transcription is essential for its function in plant immunity. Furthermore, our study identified six amino acids in the CG-1 domain that are critical for its DNA-binding activity.
2. Materials and Methods
2.1. Identification and Selection of the Conserved Amino Acids in the CG-1 Domain for Mutational Studies
The genomic and full-length protein sequences of 1343 CAMTA transcription factors (including the splice variants) belonging to 165 plant species obtained from the “Plant Transcription Factor Database” (
http://planttfdb.gao-lab.org, accessed on 16 January 2020) were used for analysis (
File S1). A full-length protein sequence alignment across the various plant species was generated using the “T-Coffee” software, as described at
http://planttfdb.gao-lab.org, accessed on 16 January 2020. The protein sequences from multiple species were truncated to include only the CG-1 domain, which were then aligned using the Hidden Markov Model-guided method (
http://planttfdb.gao-lab.org, accessed on 16 January 2020) to generate a sequence logo showing the highly conserved amino acids [
37]. Furthermore, the Human HsCAMTA (HsCAMTA1, Q9Y6Y1) and the
Drosophila dmCAMTA (ABI94369) amino acid sequences were obtained from the NCBI database, and their CG-1 domains were compared to that of the AtCAMTA3 sequence. Based on the results of these alignments, some of the amino acids are highly conserved across all plants, human, and insects, and these were selected as candidates for mutational studies.
2.2. Generation of CAMTA3 CG-1 Domain Mutants
Total RNA was isolated from Arabidopsis using the Plant RNAeasy Kit (Qiagen, USA). About 2 µg of DNAse-treated total RNA was used for the first strand cDNA synthesis using Superscript III (Invitrogen, Waltham, MA, USA) as per manufacturer’s instructions. The full-length CAMTA3 cDNA was PCR amplified using the HS Primestar DNA polymerase (Takara, San Jose, CA, USA) with forward and reverse primers bearing BamHI and XhoI, respectively. The amplified product was gel extracted, digested with BamHI and XhoI enzymes and cloned in pET28a(+) digested with the same enzymes. After sequence verification, the full-length coding sequence clone of CAMTA3 in pET28a(+) was used as a template for the generation of the mutants.
Mutants in the CG-1 domain were created using the Q5 site-directed mutagenesis kit (New England Biolabs, Ipswich, MA, USA) as per manufacturer instructions. The primers used for creating the mutations were designed using the online design tool, NEB Base changer web portal (
https://nebasechanger.neb.com, accessed on 16 January 2020). The sequence of primers used for mutagenesis are shown in
Table S1. The generated plasmids carrying the mutations in the CG-1 domain were subsequently transformed into high-efficiency
E. coli competent cells. The mutants were then sequenced using different primers to cover the entire length of the
CAMTA3 coding sequence.
2.3. Expression and Purification of the CG-1 Domain in E. coli and DNA-Binding Assays
2.3.1. Protein Expression and Purification
To prepare CG-1 domain (153 aa) expression constructs, the CAMTA3 CG-1 domain (1–459 bps) was PCR amplified from the pET-28a(+) expression vector bearing WT or individual six-point mutations (M1 to M6) using HS Primestar DNA polymerase (Takara, San Jose, CA, USA) with forward and reverse primers bearing BamHI and XhoI, respectively. The amplified product was gel extracted, digested with BamHI and XhoI enzymes and cloned in pET28a(+) digested with the same enzymes. After verifying the sequence, the pET-28a(+) plasmid bearing WT CG-1 domain or its six-point mutants (M1 to M6) were transformed into BL21-Codon Plus (DE3)-RIL (Stratagene, San Diego, CA, USA) cells. A single transformed colony was inoculated into 5 mL of LB medium with kanamycin (50 mg/L). The culture was then incubated at 37 °C in a shaker maintained at 200 rpm for 15 h. About 500 µL of overnight culture was added to 200 mL of liquid LB medium supplemented with 50 mg/L of kanamycin and incubated at 37 °C with vigorous shaking until the O.D. reached 0.5 to 0.6 and was then induced with 0.1 mM IPTG for 4 h at 37 °C. The cells were harvested by centrifugation at 8000 rpm for 15 min at 4 °C. The pelleted cells were quickly washed with lysis buffer prior to the preparation of the extracts. The pelleted cells were resuspended in 20 mL of lysis buffer (20 mM HEPES-KOH pH 8.0, 1 M NaCl, 2 mM ß-ME, one tablet of EDTA-free complete Protease Inhibitor Cocktail, 100 mM PMSF, 1 mM imidazole). The resuspended cells were sonicated four times with a pulse interval of 10 s followed by 2 min incubation on ice. The cell extract was clarified by centrifugation (11,000 rpm, at 4 °C for 60 min). This clarified extract was used for the purification of the His-tagged proteins using Ni-NTA agarose resin (Qiagen, Germantown, MD, USA). The agarose resin was washed twice by resuspending in lysis buffer followed by centrifugation at 3000 rpm and removal of the supernatant. The clarified cell lysate was incubated with washed Ni-NTA agarose resin on a rotatory shaker for 1 h at 4 °C. After incubation, proteins bound to the resin were collected via centrifugation at 4 °C, 500× g for 5 min. The resin was repeatedly washed by suspending it in wash buffer 1 (lysis buffer +10 mM imidazole) followed by incubation on a rotatory shaker for 5 min. This step was repeated till the Abs280 of the supernatant reached zero. This was followed by a final wash with wash buffer II (lysis buffer + 20 mM imidazole). For elution of the His-Tagged proteins, the washed resin was resuspended in wash buffer II, transferred to the column and allowed to settle down. His-tagged proteins were eluted using elution buffer (lysis buffer + 250 mM imidazole). Depending upon the bead volume, multiple fractions of fixed volume were collected. His-tagged proteins in the fractions were detected using Bradford assay. The fractions resolved on polyacrylamide gel and were transblotted on to the PVDF membrane. The membrane was then probed with an anti-6X His Tag monoclonal antibody (Abcam; Waltham, MA, USA; ab49746) conjugated to alkaline phosphatase at 1:2000 dilution. The fractions that exhibited single bands of the expected size were pooled and dialyzed overnight at 4 °C against dialysis buffer (20 mM HEPES-KOH pH 8.0, 200 mM NaCl, 20% glycerol, 1 mM DTT) in a dialysis membrane bag (3500 kDa cut-off) (Spectrum, New Brunswick, NJ, USA). The dialyzed His-tagged proteins were aliquoted and then flash-frozen in liquid nitrogen and stored at −80 °C.
The flash-frozen purified proteins were thawed on ice, and the protein concentration was estimated using Bradford reagent (Bio-Rad, Hercules, CA, USA). Then, 10 µg of the purified protein was boiled in 1X sample buffer, resolved on 12% polyacrylamide gel with 10% SDS and electro-blotted onto a PVDF (Bio-Rad, Hercules, CA, USA) membrane. The blot was blocked overnight with 5% nonfat milk in TBST buffer (50 mM Tris-HCl, pH 7.5, 150 mM NaCl, 0.05% Tween-20). The membrane was then probed with an anti-6X His Tag monoclonal antibody (Abcam; ab49746) conjugated to alkaline phosphatase at 1:2000 dilution and detected with an alkaline phosphatase detection system.
2.3.2. EMSA Assays
Electrophoretic mobility shift assay (EMSA) for studying DNA protein interaction was performed with purified CG-1 (DNA-binding domain) variants using a non-isotopic method. For this purpose, the LightShift Chemiluminescent EMSA kit (Thermo Fischer Scientific, Waltham, MA, USA) was used as per the manufacturer’s instructions. The DNA probe was made by combining two oligos with one being 3′-end biotin labeled containing the binding site of CAMTA3. The binding studies were conducted by incubating the 3′-end-labeled DNA probe (20 fmol) with the purified protein in a final reaction volume of 20 µL consisting of 1X binding buffer, 50 ng/µl Poly (dI. dC) with or without 4 pmol of unlabeled DNA at room temperature for 30 min. The reaction was terminated using 1X loading dye, resolved on 5% native polyacrylamide gel (in 0.5X TBE) electrophoresis and electro-blotted onto a nylon membrane. Subsequently, the membrane was cross-linked at 120 mJ/cm2 using a UV cross-linker with 254 nm bulbs for 1 min. The membrane was developed using a chemiluminescent detection module (Thermo Fischer Scientific, Waltham, MA, USA) according to the manufacturer’s instructions. Briefly, the membrane was blocked with blocking buffer for 15 min, transferred to a blocking solution supplemented with stabilized Streptavidin-Horseradish Peroxidase Conjugate and incubated on a shaker for an additional 15 min. Subsequently, the membrane was washed four times with 1X wash buffer and incubated in substrate buffer for 5 min. The chemiluminescent substrate solution was poured onto the membrane and incubated for 5 min in dark. The membrane was wrapped in saran wrap and exposed to a gel documentation system equipped with a CCD camera (BioRad, Hercules, CA, USA) to capture the signal.
To determine the minimum amount of protein required for binding to the probe, EMSA assays were performed using different protein concentrations in the presence of a constant amount of the labeled DNA. The chemiluminescence signal for each band was quantified using gel documentation software (Image Lab, BioRad, Hercules, CA, USA), and the fraction bound was plotted as a function of protein concentration.
2.4. Generation of Effector and Reporter Constructs
2.4.1. Effector Constructs
The WT as well as the mutated versions of
CAMTA3 CG-1 mutants (
M1–
M6) were PCR amplified using the primers indicated in
Table S1. The fragments were then cloned into
pFGC5941 binary vector (downstream to
CaMV35S promoter) between the
Asc1 and
BamH1 sites using the restriction sites for
AscI and
BamHI that were added to the forward and reverse primers, respectively. All the generated constructs were sequence verified, and then each construct was transformed into
Agrobacterium tumefaciens strain GV3101 and subsequently used for the generation of transgenic lines and transient assay experiment.
2.4.2. Reporter Constructs
For making the reporter constructs for the transient assay experiments, a previously reported
4X RSRE sequence together with the upstream sequence of a minimal NOS promoter (−101 to +4) [
34] (
4XRSRE NOS) was synthesized. A mutated version (
4XmRSRE NOS) was also synthesized by changing the core binding motifs (
Supplementary Figure S4). The
4XRSRE NOS sequence was amplified with
PstI and
StuI restriction sites at the 5′ end and
SalI and
NcoI at the 3′ end to facilitate cloning upstream of the luciferase reporter gene. The synthesized DNA fragments were amplified using high-fidelity PrimeStar HS DNA polymerase with the end primers (
Table S1) that contained the above-mentioned restriction sites. The amplicons were gel purified and digested with
PstI and
SalI, and then used for replacing the
CaMV35S promoter upstream of the luciferase gene in the
pUC18 vector, resulting in pUC18
4XRSRE NOS::LUC or
4XmRSRE NOS::LUC expression constructs (
Supplementary Figure S5a). The resulting constructs were then digested with the
SacI, and the ends were blunted with the Fast End Repair Kit (Thermofisher, Waltham, MA, USA) followed by another digestion with
StuI to obtain blunt-end fragments consisting of
4XRSRE NOS::LUC or
4XmRSRE NOS::LUC. These fragments were then inserted into binary vector
pFGC5941, which was previously digested with
StuI and
SmaI. The resulting plasmids were
pFGC5941 4XRSRE NOS::LUC-OCS and
pFGC5941 4XmRSRE NOS::LUC-OCS (
Supplementary Figure S5b).
2.5. Generation of the Transgenic Lines
Each of the generated
pFGC5941 constructs with WT or CG-1 mutants of CAMTA3 was transformed into
Agrobacterium tumefaciens and subsequently used to transform
camta3 mutant plants using the floral dipping method. The transgenic plants were then selected on MS plates containing Glufosinate ammonium (Crescent Chemical Co, Islandia, NY, USA) (10 µg ml
−1) and genotyped using RT-PCR using the primers listed in
Table S1. The selected plants were then selfed to obtain homozygous lines, and three independent lines were chosen for each construct.
2.6. RNA Extraction and Gene Expression Analysis
For RNA isolation, leaf tissue was collected from 4-week-old plants grown in soil and flash-frozen in liquid nitrogen. Total RNA was isolated using TRIzol reagent (Invitrogen, Waltham, MA, USA) and treated with an RNase-free DNase (Promega, Madison, WI, USA) to remove any genomic DNA contamination. Then, 2 µg of the DNAse-treated RNA was reverse transcribed into cDNA using oligo dT primer and Superscript II reverse transcriptase (Invitrogen, Waltham, MA, USA) according to the manufacturer’s instructions. The cDNA was diluted with 80 µL sterile nuclease-free water. Expression analysis was performed using RT-qPCR in a Roche LC480 machine (Roche Diagnostics, Indianapolis, IN, USA) using the preprogrammed “SYBR green-I 96 well program”. For every qPCR reaction, 10 µL of 2X LightCycler 480 SYBR Green I Master mix (Roche Diagnostics, Indianapolis, IN, USA) was used along with 1 µL of 5 µM of each primer and 2.5 µL cDNA template in a final reaction volume of 20 µL.
ACTIN2 was used as a reference gene as this gene does not exhibit any difference in its expression level among the various genotypes under different conditions [
25]. Fold change in expression was calculated and plotted with respect to control treatments. Three biological replicates were used for each experiment. Primers (see
Table S1) for Real-time qPCR (RT-qPCR) were designed using the Primer Quest web tool (
http://www.idtdna.com/Primerquest/Home/Index, accessed on 10 November 2017) from Integrated DNA Technologies (IDT, Boulder, CO, USA).
2.7. Transient Expression Assay in Nicotiana Benthamiana
For the transient expression assay, we used
Agrobacterium tumefaciens cells containing
CaMV35S::CAMTA3-OCS or
CaMV 35S::CAMTA3-OCS mutants (
M1–
M6) as effector constructs and
pFGC5941 4XRSRE NOS::LUC:OCS or pFGC5941 4XmRSRE NOS::LUC-OCS as reporter constructs. The Agrobacterium cultures were grown for 36 h to OD600 ~1.0–1.2. Individual cultures were then centrifuged at 3000 rpm for 20 min. Supernatants were then removed, and the pellets were re-suspended in 10 mL infiltration medium (2 mM Na
3PO
4, 50 mM MES, 0.5% glucose, 100 μM Acetosyringone) followed by incubation at room temperature for 3 h. Each culture was then adjusted to OD600 = 1.0, and the transgenes to be co-expressed were mixed in a 1:1 ratio. Each combination was then mixed with p19 culture in a 1:1 ratio before infiltration to suppress post-transcriptional gene silencing (PTGS) and enhance transient expression [
38]. Specifically, the
RSRE NOS::LUC construct or its dysfunctional mutant versions,
mRSRE NOS::LUC, were infiltrated independently in
Nicotiana benthamiana leaves, alone or together with
CaMV35S::CAMTA3 or one of its mutant versions (
M1–
M6). The combined cultures were spot-infiltrated into six-week-old Nicotiana benthamiana leaves. Three plants (with three leaves per plant) were infiltrated with each combination. After 3 days, infiltrated leaves were collected and then flash-frozen in liquid nitrogen for protein extraction and quantification of LUC activity.
2.8. Protein Extraction and Quantification of LUC Activity
Infiltrated leaves were flash-frozen, ground to a fine powder in TissueLyser and suspended at room temperature in 1X CCLR (25 mM trisphosphate (pH 7.8), 2 mM DTT, 2 mM 1,2-diaminocyclohexane-N,N,N’,N´-tetraacetic acid, 10% glycerol, 1% Triton® X-100) (Promega, Madison, WI, USA) with further homogenization on a rocker. After cell lysis, the extract was clarified by centrifugation at 4°C for 10 min at 13,500 rpm using standard assay conditions. The LUC activity was assayed in the supernatant using the Luciferase Assay System (Promega, Madison, WI, USA). Briefly, 20 μL of cell lysate was added to 100 μL of the Luciferase Assay Reagent into a tube and vortexed for 10 s prior to placing it in a cuvette holder. The generated luminescence was measured using a luminometer (Turner Designs, San Jose, CA, USA) programmed to perform a 2 s measurement delay followed by a 10 s measurement read. Protein concentration was determined using the Bradford reagent (Bio-Rad, Hercules, CA, USA).
2.9. Protein Extraction and Immunoblot Analysis
For extraction of total protein, the leaves of the 30-day-old plants were flash-frozen, ground to fine powder in TissueLyser and dissolved in 500 µL protein extraction buffer (40 mM K2HPO4, 10 mM KH2PO4, 1 mg mL−1 ascorbate, 0.05% β-mercaptoethanol (v/v) 0.1% TritonX-100, 1 mM PMSF) containing 1% protease inhibitor cocktail (Sigma-Aldrich, St. Louis, MO, USA). The extract was centrifuged for 10 min at 13,500 rpm at 4 °C. Protein concentration was determined using the Bradford reagent (Bio-Rad Hercules, CA, USA). An amount of 40 µg of total protein from each sample was resolved in 12% SDS gel and blotted onto a PVDF membrane (Millipore, Burlington, MA, USA). The blot was blocked with 5% non-fat milk in TBST buffer (50 mM Tris–HCl, pH 7.5, 150 mM NaCl, 0.05% Tween-20). Then, the membrane was probed with an anti-AtCAMTA3 polyclonal antibody at a 1:500 dilution and detected with an antirabbit secondary antibody conjugated with horseradish peroxidase using the chemiluminescence detection system (Pierce, ThermoFischer Scientific, Waltham, MA, USA).
4. Discussion
Several genetic and biochemical studies have shown that the transcriptional regulatory activity of CAMTAs is associated with their binding to the (
A/C/G)
CGCG(
T/C/G), (
A/C)
CGTGT or
CG(
C/T)
G cis-elements in the promoter regions of their target genes, primarily through the CG-1 domain [
8,
10,
11,
17,
41]. However, more recently, Lolle et al. [
35] suggested that CAMTA3 acts as a guardee for NLR immune receptors in modulating plant immunity rather than having a direct transcriptional activity role in regulating plant immunity. They have identified two TIR-type NLR proteins, DSC1 and DSC2, to be guards for the host effector target (guardee) CAMTA3. They suggested that the autoimmunity in
camta3 plants results from the ectopic activation of the two NLRs, DSC1 and DSC2, rather than the loss of the CAMTA3 transcriptional repressor function per se [
35]. The
camta1 or
camta2 null alleles in
camta3 mutants enhance the latter’s autoimmune phenotype, whereas neither
camta1 nor
camta2 shows an autoimmune phenotype [
24]. Interestingly, the expression of
DSC1-DN or
DSC2-DN versions in the
camta1 camta3 double mutant did not rescue the autoimmune phenotype [
35]. Therefore, if CAMTA3 works only as a guardee for DSC1, the expression of
DSC1-DN or
DSC2-DN into the
camta1 camta3 background should be able to repress the autoimmune phenotype. In another study, the expression of a truncated version of CAMTA3
1–344 that includes only the CAMTA3 CG-1 domain suppressed the autoimmune phenotype of
camta2 camta3 [
18]. Moreover, the expression of the
DSC1 orthologue of
Gossypium hirsutum (
GhDSC1) as well as
CAMTA3 was found to be induced in response to the
Verticillium wilt and JA treatment in
Arabidopsis, suggesting a coordinated
DSC1 and
CAMTA3 response in
Arabidopsis to
Verticillium wilt [
42]. CAMTA3 has also been reported as a negative regulator for plant immunity by direct binding to the promoter regions of the SA positive regulator
CBP60g as well as the TNL- and CNL-mediated defense pathways regulators,
EDS1 and
NDR1, respectively, and suppressing their expression [
14,
17,
27]. Additionally, Doherty et al. [
26] demonstrated that the CG-1 domains of CAMTA1, 2, 3 and 5 bind to the
CM2 motif in the promoter region of
CBF2. They also provided evidence that CAMTA3 acts as a positive regulator for cold-induced
CBFs gene expression in Arabidopsis [
30].
One way to address whether DNA binding and transcriptional activities of CAMTA3 are necessary for regulating plant immunity is to generate
CAMTA3 mutants that are unable to exhibit DNA binding and thus abolish their transcriptional activity. Irrespective of the plant species, virtually all CAMTAs have a CG-1 domain in the N-terminal region of the protein, indicating that the position of this domain is evolutionarily conserved [
8,
9,
14]. The CG-1 domain is also present in human and animal CAMTAs [
8] (
Figure 1). Our analysis of the CG-1 domain revealed a high conservation of aromatic amino acids such as W, H, Y and F in the CG-1 domain indicating their likely role in DNA binding [
43]. Similarly, a high occurrence of these W and Y amino acids was also noted in the DBD of WRKY, ERF, MYB and MYB-related TFs that are also involved in plant immune response [
44]. Since
camta3 mutants exhibit temperature-dependent constitutive autoimmune response, we hypothesized that the necessity of CAMTA3 transcriptional activity can be effectively scored by expressing WT and
CAMTA3 CG-1 mutant variants in the
camta3 background and see if it will rescue the mutant phenotype. Remarkably, the majority of the CG-1 mutant variants (M1–M4 and M6) that exhibited no DNA-binding activity also failed to rescue the mutant phenotype, while WT and M5 with DNA-binding activity did complement the phenotype (
Figure 5). Together, these results suggest that the transcriptional activity of CAMTA3 is required for its function in plant immunity. We further validated this by analyzing the expression of several CAMTA3-regulated genes that were previously reported to be involved in plant immunity and cold response [
17,
18,
25,
26,
27]. The expression levels of all the immune and cold response genes in the
M1 to
M4 and
M6 variants were similar to that in the
camta3 mutant, whereas their expression in the
M5 mutant was like WT plants (
Figure 6). Furthermore, to exclude the possibility that some of these results were due to the in vivo destabilization of CAMTA3 proteins as a result of the mutation, we analyzed the proteins levels and compared them with that of WT. Our results revealed similar levels of protein in all the CG-1 mutants and WT plants, implying that these CG-1 mutations did not alter the stability of the CAMTA3 protein. Collectively, our results provide strong biochemical and genetic evidence that the transcriptional activity of CAMTA3 is essential for its function. The requirement of transcriptional activity of CAMTA3 for its repressor and activator function was further supported by the function exhibited by the NRM module (CAMTA
1–334), a truncated version of CAMTA3 protein that includes the N terminal CG-1 DNA-binding domain and lacks other domains [
18,
45]. The expression of NRM in the
camta2 camta3 double mutant not only suppressed the expression of
ICS1,
CBP60g and
PRI but also rescued the autoimmune phenotype of this mutant [
18]. In another study, NRM (CAMTA3
334) was also reported to be sufficient to rapidly induce the expression of
CBF2 and two other cold-inducible genes,
EXPansin-Like A1 (
EXPL1) and
Nine-Cis-Epoxycarotenoid Dioxygenase 3 (
NCED3), under cold stress [
18,
45]. However, these studies did not identify the amino acids involved in the DNA-binding activity of the NRM. In this context, our study provides evidence for the role of the amino acids involved in the transcriptional activity of CAMTA3.
As CAMTA3 stability and turnover are known to be critical for its function as a repressor for the immune response [
46,
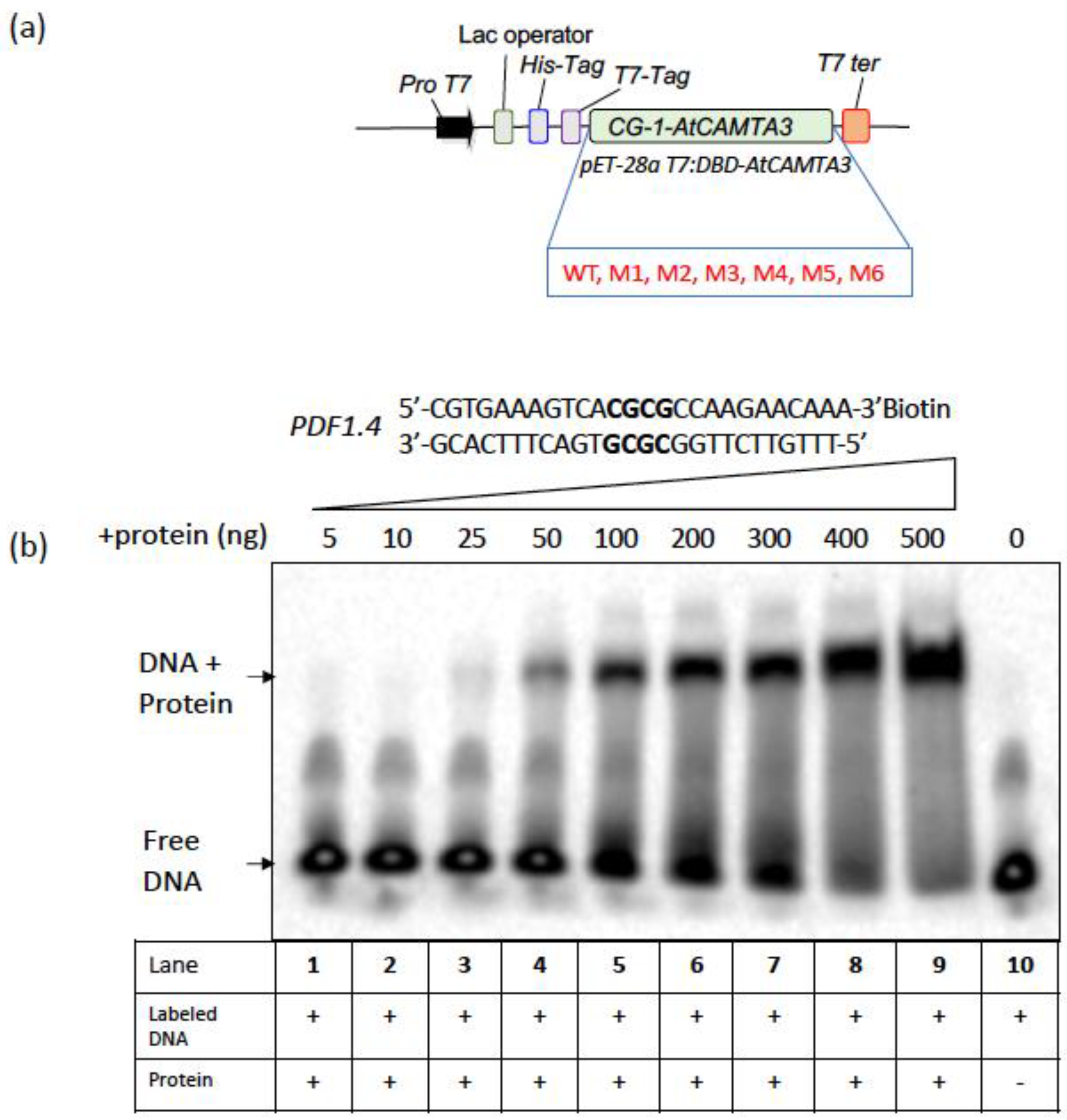
47], we show that the minimum concentration of 25 ng of WT CAMTA3 CG-1 domain to be required for binding to
PDF1.4 (
Figure 2). However, most of the CAMTA3 CG-1 mutants (M1, M2, M3, M4 and M6) failed to bind to the same probe even at a higher protein concentration of 500 ng (
Figure 3), indicating that these amino acids are critical for DNA-binding and CAMTA3 transcriptional activity. Interestingly, the M5 mutant exhibited binding to the probe, albeit at higher concentrations (100 ng), indicating weak binding (
Figure 3). Previously, it has been shown that the expression of the truncated variant of CAMTA3, comprising the CG-1 domain (334 amino acids, NRM), exhibited transcriptional activation and repressor activities in plants [
18]. Interestingly, mutations of K108A and K141E in the CG-1 domain of the human CAMTA2 and CAMTA1, respectively (
Figure 1), completely abolished their DNA-binding activity, indicating a critical role for these amino acids in the DNA-binding activity of these CAMTAs [
48]. Indeed, this amino acid (K) is also conserved across the plant kingdom; however, the degree of conservation is slightly lower than that observed with other aromatic amino acids (
Figure 1). However, the role of K in DNA-binding cannot be ruled out in plant CAMTAs, and additional mutational studies are needed.
Previously, CAMTA3 has been shown to bind to the
RSRE elements and activate downstream genes [
31]. In that study, the expression of full-length WT and CAMTA3 mutant proteins transiently in tobacco, and their ability to activate the expression of luciferase gene driven by
RSRE (
vCGCGb) was tested. In agreement with earlier reports, the binding of WT CAMTA3 is highly specific, and it transcriptionally activates the luciferase gene expression. However, except for the M5 mutant, all other mutants failed to activate the luciferase reporter gene expression (
Figure 4), which concurred with our results in
Figure 2 and
Figure 3, with truncated CAMTA3 proteins.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}