
Axon-Autonomous Effects of the Amyloid Precursor Protein Intracellular Domain (AICD) on Kinase Signaling and Fast Axonal Transport

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
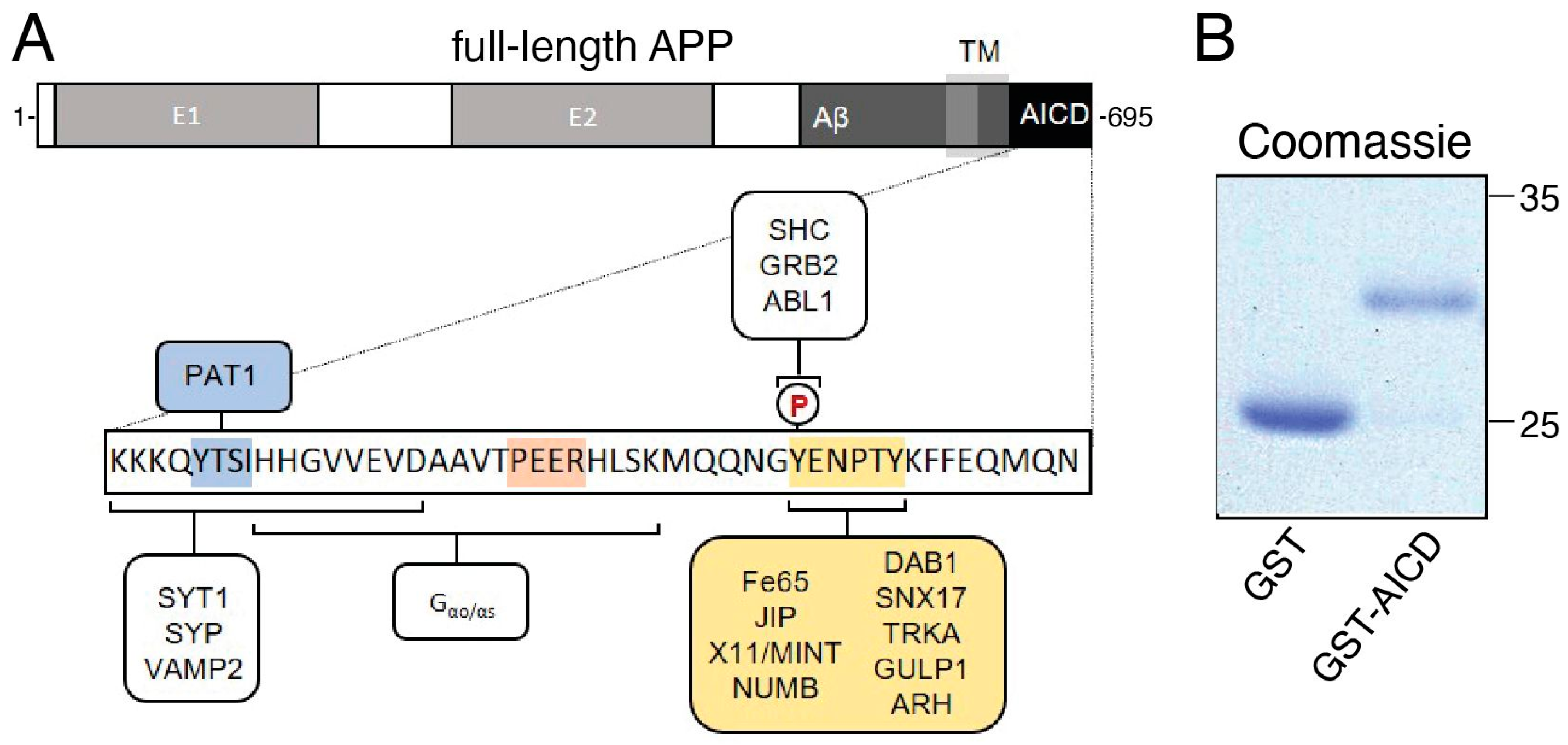
2.1. Recombinant GST–AICD Proteins
2.2. Recombinant Fe65 PTB2-6xHis Protein
2.3. Squid Axoplasm Vesicle Motility Assay
2.4. Immunoprecipitation of P32-Radiolabeled Conventional Kinesin
2.5. Immunoblot-Based Analysis of Kinase Activity
2.6. Statistics
3. Results
3.1. AICD Inhibits Anterograde and Retrograde FAT, Increasing also Phosphorylation of the Motor Protein Conventional Kinesin
3.2. AICD Activates Selected Axonal Kinases
3.3. Inhibitory Effects of AICD on FAT Involve Activation of Specific MAPKs and CK2

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Figures Featuring Data | Experimental Comparisons (vs) | p Value | ||
|---|---|---|---|---|
| Anterograde | Retrograde | |||
| Figure 2 | Buffer X/2 (control) (n = 7) | AICD (n = 7) | <0.0001 | <0.0001 |
| Figure 4 | AICD (n = 7) | AICD + SB203580 (n = 3) | <0.0001 | <0.0001 |
| AICD (n = 7) | AICD + SP600125 (n = 3) | <0.0001 | <0.0001 | |
| AICD (n = 7) | AICD + ING35 (n = 3) | 0.0036 | <0.0001 | |
| AICD (n = 7) | AICD + TBCA (n = 3) | <0.0001 | 0.0009 | |
| Buffer (n = 7) | AICD + SB203580 (n = 3) | <0.0001 | n.s. (0.7361) | |
| Buffer (n = 7) | AICD + SP600125 (n = 3) | <0.0001 | 0.0095 | |
| Buffer (n = 7) | AICD + ING35 (n = 3) | <0.0001 | 0.0163 | |
| Buffer (n = 7) | AICD + TBCA (n = 3) | <0.0001 | 0.0011 | |
| Figure 5 | AICD (n = 7) | AICDΔYTSI (n = 3) | <0.0001 | <0.0001 |
| AICD (n = 7) | AICDΔPEER (n = 2) | 0.0005 | <0.0001 | |
| AICD (n = 7) | AICDΔNPTY (n = 6) | <0.0001 | <0.0001 | |
| Buffer (n = 7) | AICDΔYTSI (n = 3) | <0.0001 | <0.0001 | |
| Buffer (n = 7) | AICDΔPEER (n = 2) | <0.0001 | <0.0001 | |
| Buffer (n = 7) | AICDΔNPTY (n = 6) | <0.0001 | n.s. (0.0026) | |
| Figure 6 | AICD (n = 7) | AICD + PTB2-His (n = 5) | <0.0001 | <0.0001 |
| Buffer (n = 7) | AICD + PTB2-His (n = 5) | <0.0001 | n.s. (0.2272) | |
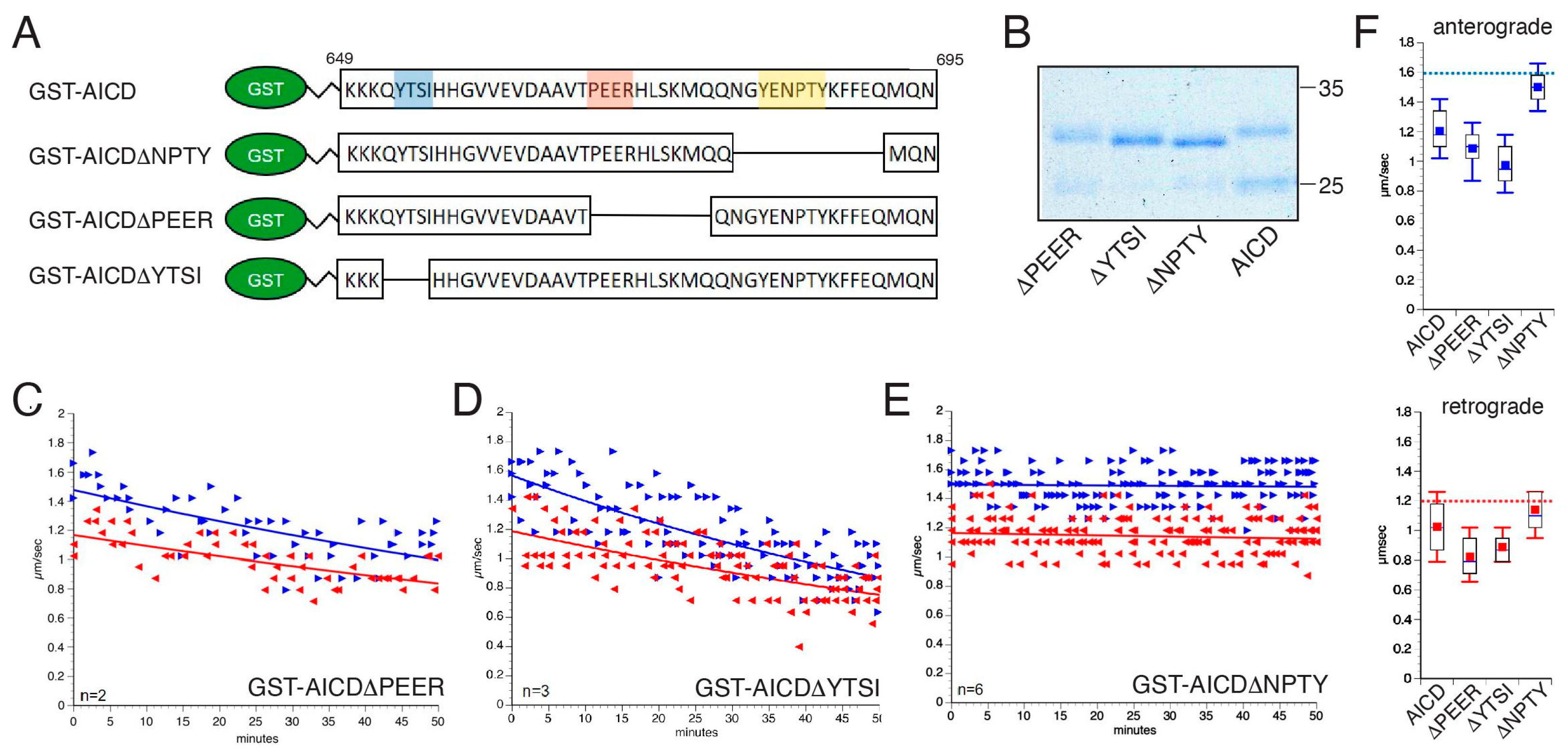
3.4. The Inhibitory Effect of AICD on FAT Depends on a Peptide Sequence Encompassing the NPTY Motif
3.5. Inhibitory Effects of AICD on FAT Involve NPTY Motif Interactions with Endogenous PTB-Containing Squid Proteins
4. Discussion
4.1. AICD Impacts FAT via Activation of Axonal Kinases
4.2. AICD’s Potential Involvement in AD-Linked Axonopathy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Black, M.M. Axonal transport: The orderly motion of axonal structures. Methods Cell Biol. 2016, 131, 1–19. [Google Scholar] [CrossRef]
- Morfini, G.; Schmidt, N.; Weissmann, C.; Pigino, G.; Kins, S. Conventional kinesin: Biochemical heterogeneity and functional implications in health and disease. Brain Res. Bull. 2016, 126, 347–353. [Google Scholar] [CrossRef]
- Brady, S.T.; Morfini, G.A. Regulation of motor proteins, axonal transport deficits and adult-onset neurodegenerative diseases. Neurobiol. Dis. 2017, 105, 273–282. [Google Scholar] [CrossRef]
- Kanaan, N.M.; Pigino, G.F.; Brady, S.T.; Lazarov, O.; Binder, L.I.; Morfini, G.A. Axonal degeneration in Alzheimer’s disease: When signaling abnormalities meet the axonal transport system. Exp. Neurol. 2013, 246, 44–53. [Google Scholar] [CrossRef]
- Morfini, G.A.; You, Y.-M.; Pollema, S.L.; Kaminska, A.; Liu, K.; Yoshioka, K.; Björkblom, B.; Coffey, E.T.; Bagnato, C.; Han, D.; et al. Pathogenic huntingtin inhibits fast axonal transport by activating JNK3 and phosphorylating kinesin. Nat. Neurosci. 2009, 12, 864–871. [Google Scholar] [CrossRef]
- Morfini, G.A.; Bosco, D.A.; Brown, H.; Gatto, R.; Kaminska, A.; Song, Y.; Molla, L.; Baker, L.; Marangoni, M.N.; Berth, S.; et al. Inhibition of fast axonal transport by pathogenic SOD1 involves activation of p38 MAP kinase. PLoS ONE 2013, 8, e65235. [Google Scholar] [CrossRef]
- Bosco, D.A.; Morfini, G.; Karabacak, N.M.; Song, Y.; Gros-Louis, F.; Pasinelli, P.; Goolsby, H.; Fontaine, B.A.; Lemay, N.; McKenna-Yasek, D.; et al. Wild-type and mutant SOD1 share an aberrant conformation and a common pathogenic pathway in ALS. Nat. Neurosci. 2010, 13, 1396–1403. [Google Scholar] [CrossRef]
- Sama, R.R.K.; Fallini, C.; Gatto, R.; McKeon, J.E.; Song, Y.; Rotunno, M.S.; Penaranda, S.; Abdurakhmanov, I.; Landers, J.E.; Morfini, G.; et al. ALS-linked FUS exerts a gain of toxic function involving aberrant p38 MAPK activation. Sci. Rep. 2017, 7, 115. [Google Scholar] [CrossRef]
- Pigino, G.; Morfini, G.; Atagi, Y.; Deshpande, A.; Yu, C.; Jungbauer, L.; LaDu, M.; Busciglio, J.; Brady, S. Disruption of fast axonal transport is a pathogenic mechanism for intraneuronal amyloid beta. Proc. Natl. Acad. Sci. USA 2009, 106, 5907–5912. [Google Scholar] [CrossRef]
- Müller, U.C.; Deller, T.; Korte, M. Not just amyloid: Physiological functions of the amyloid precursor protein family. Nat. Rev. Neurosci. 2017, 18, 281–298. [Google Scholar] [CrossRef]
- Müller, T.; Meyer, H.E.; Egensperger, R.; Marcus, K. The amyloid precursor protein intracellular domain (AICD) as modulator of gene expression, apoptosis, and cytoskeletal dynamics-relevance for Alzheimer’s disease. Prog. Neurobiol. 2008, 85, 393–406. [Google Scholar] [CrossRef] [PubMed]
- Flammang, B.; Pardossi-Piquard, R.; Sevalle, J.; Debayle, D.; Dabert-Gay, A.-S.; Thévenet, A.; Lauritzen, I.; Checler, F. Evidence that the amyloid-β protein precursor intracellular domain, AICD, derives from β-secretase-generated C-terminal fragment. J. Alzheimers. Dis. 2012, 30, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Bignante, E.A.; Heredia, F.; Morfini, G.; Lorenzo, A. Amyloid β precursor protein as a molecular target for amyloid β-induced neuronal degeneration in Alzheimer’s disease. Neurobiol. Aging 2013, 34, 2525–2537. [Google Scholar] [CrossRef] [PubMed]
- Konietzko, U. AICD nuclear signaling and its possible contribution to Alzheimer’s disease. Curr. Alzheimer Res. 2012, 9, 200–216. [Google Scholar] [CrossRef]
- Pousinha, P.A.; Mouska, X.; Bianchi, D.; Temido-Ferreira, M.; Rajão-Saraiva, J.; Gomes, R.; Fernandez, S.P.; Salgueiro-Pereira, A.R.; Gandin, C.; Raymond, E.F.; et al. The Amyloid Precursor Protein C-Terminal Domain Alters CA1 Neuron Firing, Modifying Hippocampus Oscillations and Impairing Spatial Memory Encoding. Cell Rep. 2019, 29, 317–331.e5. [Google Scholar] [CrossRef]
- Rajão-Saraiva, J.; Dunot, J.; Ribera, A.; Temido-Ferreira, M.; Coelho, J.E.; König, S.; Moreno, S.; Enguita, F.J.; Willem, M.; Kins, S.; et al. Age-dependent NMDA receptor function is regulated by the amyloid precursor protein. Aging Cell 2023, 22, e13778. [Google Scholar] [CrossRef]
- Ryan, K.A.; Pimplikar, S.W. Activation of GSK-3 and phosphorylation of CRMP2 in transgenic mice expressing APP intracellular domain. J. Cell Biol. 2005, 171, 327–335. [Google Scholar] [CrossRef]
- Kim, H.-S.; Kim, E.-M.; Lee, J.-P.; Park, C.H.; Kim, S.; Seo, J.-H.; Chang, K.-A.; Yu, E.; Jeong, S.-J.; Chong, Y.H.; et al. C-terminal fragments of amyloid precursor protein exert neurotoxicity by inducing glycogen synthase kinase-3beta expression. FASEB J. 2003, 17, 1951–1953. [Google Scholar] [CrossRef]
- Margevicius, D.R.; Bastian, C.; Fan, Q.; Davis, R.J.; Pimplikar, S.W. JNK-interacting protein 1 mediates Alzheimer’s-like pathological features in AICD-transgenic mice. Neurobiol. Aging 2015, 36, 2370–2379. [Google Scholar] [CrossRef]
- Gibbs, K.L.; Greensmith, L.; Schiavo, G. Regulation of Axonal Transport by Protein Kinases. Trends Biochem. Sci. 2015, 40, 597–610. [Google Scholar] [CrossRef]
- Dai, J.; Buijs, R.M.; Kamphorst, W.; Swaab, D.F. Impaired axonal transport of cortical neurons in Alzheimer’s disease is associated with neuropathological changes. Brain Res. 2002, 948, 138–144. [Google Scholar] [CrossRef]
- Wang, Z.-X.; Tan, L.; Yu, J.-T. Axonal transport defects in Alzheimer’s disease. Mol. Neurobiol. 2015, 51, 1309–1321. [Google Scholar] [CrossRef] [PubMed]
- Morfini, G.A.; Burns, M.; Binder, L.I.; Kanaan, N.M.; LaPointe, N.; Bosco, D.A.; Brown, R.H.; Brown, H.; Tiwari, A.; Hayward, L.; et al. Axonal transport defects in neurodegenerative diseases. J. Neurosci. 2009, 29, 12776–12786. [Google Scholar] [CrossRef] [PubMed]
- Lazarov, O.; Morfini, G.A.; Lee, E.B.; Farah, M.H.; Szodorai, A.; DeBoer, S.R.; Koliatsos, V.E.; Kins, S.; Lee, V.M.-Y.; Wong, P.C.; et al. Axonal transport, amyloid precursor protein, kinesin-1, and the processing apparatus: Revisited. J. Neurosci. 2005, 25, 2386–2395. [Google Scholar] [CrossRef] [PubMed]
- Radzimanowski, J.; Beyreuther, K.; Sinning, I.; Wild, K. Overproduction, purification, crystallization and preliminary X-ray analysis of human Fe65-PTB2 in complex with the amyloid precursor protein intracellular domain. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2008, 64, 409–412. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Kang, M.; Morfini, G.; Brady, S.T. Fast axonal transport in isolated axoplasm from the squid giant axon. Methods Cell Biol. 2016, 131, 331–348. [Google Scholar] [CrossRef]
- Kozikowski, A.P.; Gaisina, I.N.; Yuan, H.; Petukhov, P.A.; Blond, S.Y.; Fedolak, A.; Caldarone, B.; McGonigle, P. Structure-based design leads to the identification of lithium mimetics that block mania-like effects in rodents. possible new GSK-3beta therapies for bipolar disorders. J. Am. Chem. Soc. 2007, 129, 8328–8332. [Google Scholar] [CrossRef] [PubMed]
- LaPointe, N.E.; Morfini, G.; Pigino, G.; Gaisina, I.N.; Kozikowski, A.P.; Binder, L.I.; Brady, S.T. The amino terminus of tau inhibits kinesin-dependent axonal transport: Implications for filament toxicity. J. Neurosci. Res. 2009, 87, 440–451. [Google Scholar] [CrossRef]
- Kang, M.; Baker, L.; Song, Y.; Brady, S.T.; Morfini, G. Biochemical analysis of axon-specific phosphorylation events using isolated squid axoplasms. Methods Cell Biol. 2016, 131, 199–216. [Google Scholar] [CrossRef]
- Pfister, K.K.; Wagner, M.C.; Stenoien, D.L.; Brady, S.T.; Bloom, G.S. Monoclonal antibodies to kinesin heavy and light chains stain vesicle-like structures, but not microtubules, in cultured cells. J. Cell Biol. 1989, 108, 1453–1463. [Google Scholar] [CrossRef]
- Grabinski, T.; Kanaan, N.M. Novel Non-phosphorylated Serine 9/21 GSK3β/α Antibodies: Expanding the Tools for Studying GSK3 Regulation. Front. Mol. Neurosci. 2016, 9, 123. [Google Scholar] [CrossRef] [PubMed]
- Salvadores, N.; Gerónimo-Olvera, C.; Court, F.A. Axonal Degeneration in AD: The Contribution of Aβ and Tau. Front. Aging Neurosci. 2020, 12, 581767. [Google Scholar] [CrossRef] [PubMed]
- Brunholz, S.; Sisodia, S.; Lorenzo, A.; Deyts, C.; Kins, S.; Morfini, G. Axonal transport of APP and the spatial regulation of APP cleavage and function in neuronal cells. Exp. Brain Res. 2012, 217, 353–364. [Google Scholar] [CrossRef]
- Aydin, D.; Weyer, S.W.; Müller, U.C. Functions of the APP gene family in the nervous system: Insights from mouse models. Exp. Brain Res. 2012, 217, 423–434. [Google Scholar] [CrossRef]
- Kanaan, N.M.; Morfini, G.A.; LaPointe, N.E.; Pigino, G.F.; Patterson, K.R.; Song, Y.; Andreadis, A.; Fu, Y.; Brady, S.T.; Binder, L.I. Pathogenic forms of tau inhibit kinesin-dependent axonal transport through a mechanism involving activation of axonal phosphotransferases. J. Neurosci. 2011, 31, 9858–9868. [Google Scholar] [CrossRef]
- Morfini, G.; Pigino, G.; Szebenyi, G.; You, Y.; Pollema, S.; Brady, S.T. JNK mediates pathogenic effects of polyglutamine-expanded androgen receptor on fast axonal transport. Nat. Neurosci. 2006, 9, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Brady, S.T.; Pfister, K.K.; Bloom, G.S. A monoclonal antibody against kinesin inhibits both anterograde and retrograde fast axonal transport in squid axoplasm. Proc. Natl. Acad. Sci. USA 1990, 87, 1061–1065. [Google Scholar] [CrossRef] [PubMed]
- Leo, L.; Weissmann, C.; Burns, M.; Kang, M.; Song, Y.; Qiang, L.; Brady, S.T.; Baas, P.W.; Morfini, G. Mutant spastin proteins promote deficits in axonal transport through an isoform-specific mechanism involving casein kinase 2 activation. Hum. Mol. Genet. 2017, 26, 2321–2334. [Google Scholar] [CrossRef]
- King, G.D.; Scott Turner, R. Adaptor protein interactions: Modulators of amyloid precursor protein metabolism and Alzheimer’s disease risk? Exp. Neurol. 2004, 185, 208–219. [Google Scholar] [CrossRef]
- Ghosal, K.; Vogt, D.L.; Liang, M.; Shen, Y.; Lamb, B.T.; Pimplikar, S.W. Alzheimer’s disease-like pathological features in transgenic mice expressing the APP intracellular domain. Proc. Natl. Acad. Sci. USA 2009, 106, 18367–18372. [Google Scholar] [CrossRef]
- Morfini, G.; Szebenyi, G.; Richards, B.; Brady, S.T. Regulation of kinesin: Implications for neuronal development. Dev. Neurosci. 2001, 23, 364–376. [Google Scholar] [CrossRef]
- Ghosal, K.; Fan, Q.; Dawson, H.N.; Pimplikar, S.W. Tau Protein Mediates APP Intracellular Domain (AICD)-Induced Alzheimer’s-Like Pathological Features in Mice. PLoS ONE 2016, 11, e0159435. [Google Scholar] [CrossRef]
- Morris, S.L.; Brady, S.T. Tau phosphorylation and PAD exposure in regulation of axonal growth. Front. Cell Dev. Biol. 2022, 10, 1023418. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.L.; Tsai, M.-Y.; Aloe, S.; Bechberger, K.; König, S.; Morfini, G.; Brady, S.T. Defined Tau Phosphospecies Differentially Inhibit Fast Axonal Transport Through Activation of Two Independent Signaling Pathways. Front. Mol. Neurosci. 2020, 13, 610037. [Google Scholar] [CrossRef] [PubMed]
- Tiernan, C.T.; Combs, B.; Cox, K.; Morfini, G.; Brady, S.T.; Counts, S.E.; Kanaan, N.M. Pseudophosphorylation of tau at S422 enhances SDS-stable dimer formation and impairs both anterograde and retrograde fast axonal transport. Exp. Neurol. 2016, 283, 318–329. [Google Scholar] [CrossRef]
- Deyts, C.; Vetrivel, K.S.; Das, S.; Shepherd, Y.M.; Dupré, D.J.; Thinakaran, G.; Parent, A.T. Novel GαS-protein signaling associated with membrane-tethered amyloid precursor protein intracellular domain. J. Neurosci. 2012, 32, 1714–1729. [Google Scholar] [CrossRef]
- Murphy, J.A.; Stein, I.S.; Lau, C.G.; Peixoto, R.T.; Aman, T.K.; Kaneko, N.; Aromolaran, K.; Saulnier, J.L.; Popescu, G.K.; Sabatini, B.L.; et al. Phosphorylation of Ser1166 on GluN2B by PKA is critical to synaptic NMDA receptor function and Ca2+ signaling in spines. J. Neurosci. 2014, 34, 869–879. [Google Scholar] [CrossRef]
- Pousinha, P.A.; Mouska, X.; Raymond, E.F.; Gwizdek, C.; Dhib, G.; Poupon, G.; Zaragosi, L.-E.; Giudici, C.; Bethus, I.; Pacary, E.; et al. Physiological and pathophysiological control of synaptic GluN2B-NMDA receptors by the C-terminal domain of amyloid precursor protein. Elife 2017, 6, e25659. [Google Scholar] [CrossRef]
- Grimm, M.O.W.; Mett, J.; Stahlmann, C.P.; Haupenthal, V.J.; Zimmer, V.C.; Hartmann, T. Neprilysin and Aβ Clearance: Impact of the APP Intracellular Domain in NEP Regulation and Implications in Alzheimer’s Disease. Front. Aging Neurosci. 2013, 5, 98. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Südhof, T.C. A transcriptionally correction of transcriptively active complex of APP with Fe65 and histone acetyltransferase Tip60. Science 2001, 293, 115–120. [Google Scholar] [CrossRef]
- Margolis, B.; Borg, J.P.; Straight, S.; Meyer, D. The function of PTB domain proteins. Kidney Int. 1999, 56, 1230–1237. [Google Scholar] [CrossRef]
- Borg, J.P.; Ooi, J.; Levy, E.; Margolis, B. The phosphotyrosine interaction domains of X11 and FE65 bind to distinct sites on the YENPTY motif of amyloid precursor protein. Mol. Cell. Biol. 1996, 16, 6229–6241. [Google Scholar] [CrossRef]
- Götz, J.; Ittner, L.M.; Kins, S. Do axonal defects in tau and amyloid precursor protein transgenic animals model axonopathy in Alzheimer’s disease? J. Neurochem. 2006, 98, 993–1006. [Google Scholar] [CrossRef]
- Whitmarsh, A.J.; Cavanagh, J.; Tournier, C.; Yasuda, J.; Davis, R.J. A mammalian scaffold complex that selectively mediates MAP kinase activation. Science 1998, 281, 1671–1674. [Google Scholar] [CrossRef] [PubMed]
- Scheinfeld, M.H.; Roncarati, R.; Vito, P.; Lopez, P.A.; Abdallah, M.; D’Adamio, L. Jun NH2-terminal kinase (JNK) interacting protein 1 (JIP1) binds the cytoplasmic domain of the Alzheimer’s beta-amyloid precursor protein (APP). J. Biol. Chem. 2002, 277, 3767–3775. [Google Scholar] [CrossRef] [PubMed]
- Rusu, P.; Jansen, A.; Soba, P.; Kirsch, J.; Löwer, A.; Merdes, G.; Kuan, Y.-H.; Jung, A.; Beyreuther, K.; Kjaerulff, O.; et al. Axonal accumulation of synaptic markers in APP transgenic Drosophila depends on the NPTY motif and is paralleled by defects in synaptic plasticity. Eur. J. Neurosci. 2007, 25, 1079–1086. [Google Scholar] [CrossRef] [PubMed]
- Borgo, C.; D’Amore, C.; Sarno, S.; Salvi, M.; Ruzzene, M. Protein kinase CK2: A potential therapeutic target for diverse human diseases. Sig. Transduct. Target. Ther. 2021, 6, 183. [Google Scholar] [CrossRef]
- Sayed, M.; Kim, S.O.; Salh, B.S.; Issinger, O.G.; Pelech, S.L. Stress-induced activation of protein kinase CK2 by direct interaction with p38 mitogen-activated protein kinase. J. Biol. Chem. 2000, 275, 16569–16573. [Google Scholar] [CrossRef]
- Sayed, M.; Pelech, S.; Wong, C.; Marotta, A.; Salh, B. Protein kinase CK2 is involved in G2 arrest and apoptosis following spindle damage in epithelial cells. Oncogene 2001, 20, 6994–7005. [Google Scholar] [CrossRef]
- Ramser, E.M.; Gan, K.J.; Decker, H.; Fan, E.Y.; Suzuki, M.M.; Ferreira, S.T.; Silverman, M.A. Amyloid-β oligomers induce tau-independent disruption of BDNF axonal transport via calcineurin activation in cultured hippocampal neurons. Mol. Biol. Cell 2013, 24, 2494–2505. [Google Scholar] [CrossRef]
- Mueller, R.L.; Combs, B.; Alhadidy, M.M.; Brady, S.T.; Morfini, G.A.; Kanaan, N.M. Tau: A Signaling Hub Protein. Front. Mol. Neurosci. 2021, 14, 647054. [Google Scholar] [CrossRef]
- Lauritzen, I.; Pardossi-Piquard, R.; Bauer, C.; Brigham, E.; Abraham, J.-D.; Ranaldi, S.; Fraser, P.; St-George-Hyslop, P.; Le Thuc, O.; Espin, V.; et al. The β-secretase-derived C-terminal fragment of βAPP, C99, but not Aβ, is a key contributor to early intraneuronal lesions in triple-transgenic mouse hippocampus. J. Neurosci. 2012, 32, 16243–16255. [Google Scholar] [CrossRef] [PubMed]
- Aow, J.; Huang, T.-R.; Goh, Y.T.; Sun, A.X.; Thinakaran, G.; Koo, E.H. Evidence for a clathrin-independent endocytic pathway for APP internalization in the neuronal somatodendritic compartment. Cell Rep. 2023, 42, 112774. [Google Scholar] [CrossRef] [PubMed]
- Cacace, R.; Sleegers, K.; van Broeckhoven, C. Molecular genetics of early-onset Alzheimer’s disease revisited. Alzheimer’s Dement. 2016, 12, 733–748. [Google Scholar] [CrossRef] [PubMed]
- Hooper, C.; Killick, R.; Lovestone, S. The GSK3 hypothesis of Alzheimer’s disease. J. Neurochem. 2008, 104, 1433–1439. [Google Scholar] [CrossRef] [PubMed]
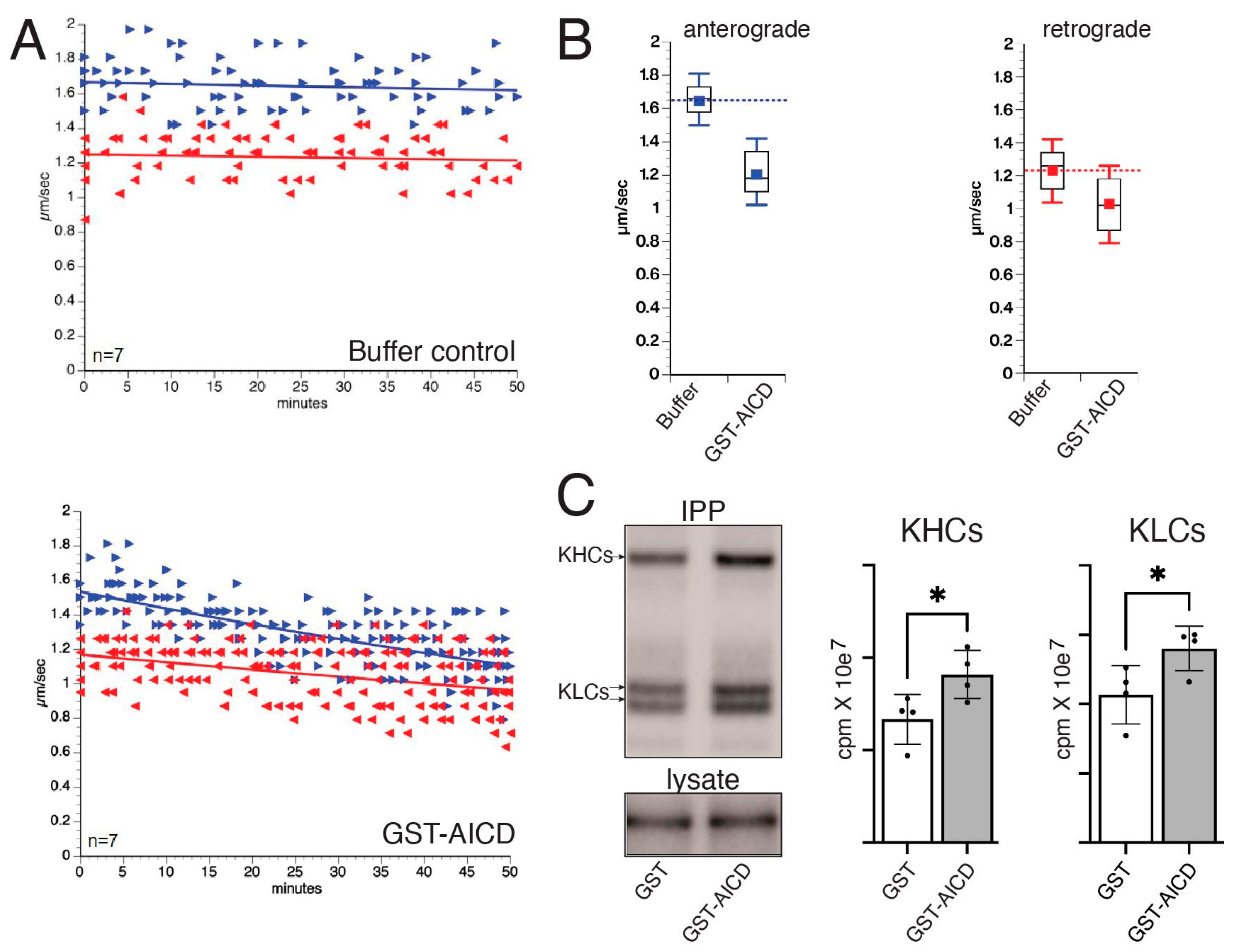



| Experimental Condition | Direction | n | Mean | Variance | Standard Deviation | Standard Error |
|---|---|---|---|---|---|---|
| Buffer X/2 (control) | Anterograde | 24 | 1.64 | 0.01 | 0.118743 | 0.024238 |
| Retrograde | 27 | 1.23 | 0.01 | 0.126022 | 0.024253 | |
| AICD | Anterograde | 81 | 1.20 | 0.03 | 0.158260 | 0.017584 |
| Retrograde | 78 | 1.02 | 0.03 | 0.181814 | 0.020586 | |
| AICD + SB203580 | Anterograde | 33 | 1.50 | 0.01 | 0.073443 | 0.012785 |
| Retrograde | 36 | 1.22 | 0.01 | 0.088644 | 0.014774 | |
| AICD + SP600125 | Anterograde | 52 | 1.51 | 0.01 | 0.107138 | 0.014857 |
| Retrograde | 50 | 1.15 | 0.01 | 0.098633 | 0.013949 | |
| AICD + ING-35 | Anterograde | 26 | 1.28 | 0.01 | 0.107972 | 0.021175 |
| Retrograde | 25 | 1.15 | 0.01 | 0.101575 | 0.020315 | |
| AICD + TBCA | Anterograde | 65 | 1.42 | 0.03 | 0.166072 | 0.020599 |
| Retrograde | 66 | 1.12 | 0.03 | 0.162290 | 0.019977 | |
| AICDΔPEER | Anterograde | 23 | 1.08 | 0.02 | 0.124697 | 0.026001 |
| Retrograde | 22 | 0.89 | 0.01 | 0.091746 | 0.019560 | |
| AICDΔYTSI | Anterograde | 46 | 0.97 | 0.02 | 0.157401 | 0.023208 |
| Retrograde | 46 | 0.82 | 0.02 | 0.146043 | 0.021533 | |
| AICDΔNPTY | Anterograde | 76 | 1.50 | 0.02 | 0.125715 | 0.014421 |
| Retrograde | 76 | 1.14 | 0.01 | 0.122468 | 0.014048 | |
| AICD + PTB2 | Anterograde | 25 | 1.40 | 0.02 | 0.165501 | 0.033100 |
| Retrograde | 27 | 1.19 | 0.02 | 0.123390 | 0.023746 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
König, S.; Schmidt, N.; Bechberger, K.; Morris, S.; Priego, M.; Zaky, H.; Song, Y.; Pielage, J.; Brunholz, S.; Brady, S.T.; et al. Axon-Autonomous Effects of the Amyloid Precursor Protein Intracellular Domain (AICD) on Kinase Signaling and Fast Axonal Transport. Cells 2023, 12, 2403. https://doi.org/10.3390/cells12192403
König S, Schmidt N, Bechberger K, Morris S, Priego M, Zaky H, Song Y, Pielage J, Brunholz S, Brady ST, et al. Axon-Autonomous Effects of the Amyloid Precursor Protein Intracellular Domain (AICD) on Kinase Signaling and Fast Axonal Transport. Cells. 2023; 12(19):2403. https://doi.org/10.3390/cells12192403
Chicago/Turabian StyleKönig, Svenja, Nadine Schmidt, Karin Bechberger, Sarah Morris, Mercedes Priego, Hannah Zaky, Yuyu Song, Jan Pielage, Silke Brunholz, Scott T. Brady, and et al. 2023. "Axon-Autonomous Effects of the Amyloid Precursor Protein Intracellular Domain (AICD) on Kinase Signaling and Fast Axonal Transport" Cells 12, no. 19: 2403. https://doi.org/10.3390/cells12192403