
Molecular Mechanisms Associated with Antifungal Resistance in Pathogenic Candida Species

 ,
,
Abstract
:1. Introduction
1.1. Candidiasis
1.1.1. Candida Species of Interest
1.1.2. Candida auris
1.2. Primary/Intrinsic Resistance vs. Secondary/Acquired Resistance
1.3. Standardized Measures of Susceptibility Testing
1.4. Geographic Influence on Rates of Antifungal Resistance
2. Antifungal Classes and Frequency of Resistance
2.1. Azoles
2.2. Polyenes
2.3. Echinocandins
2.4. 5FC
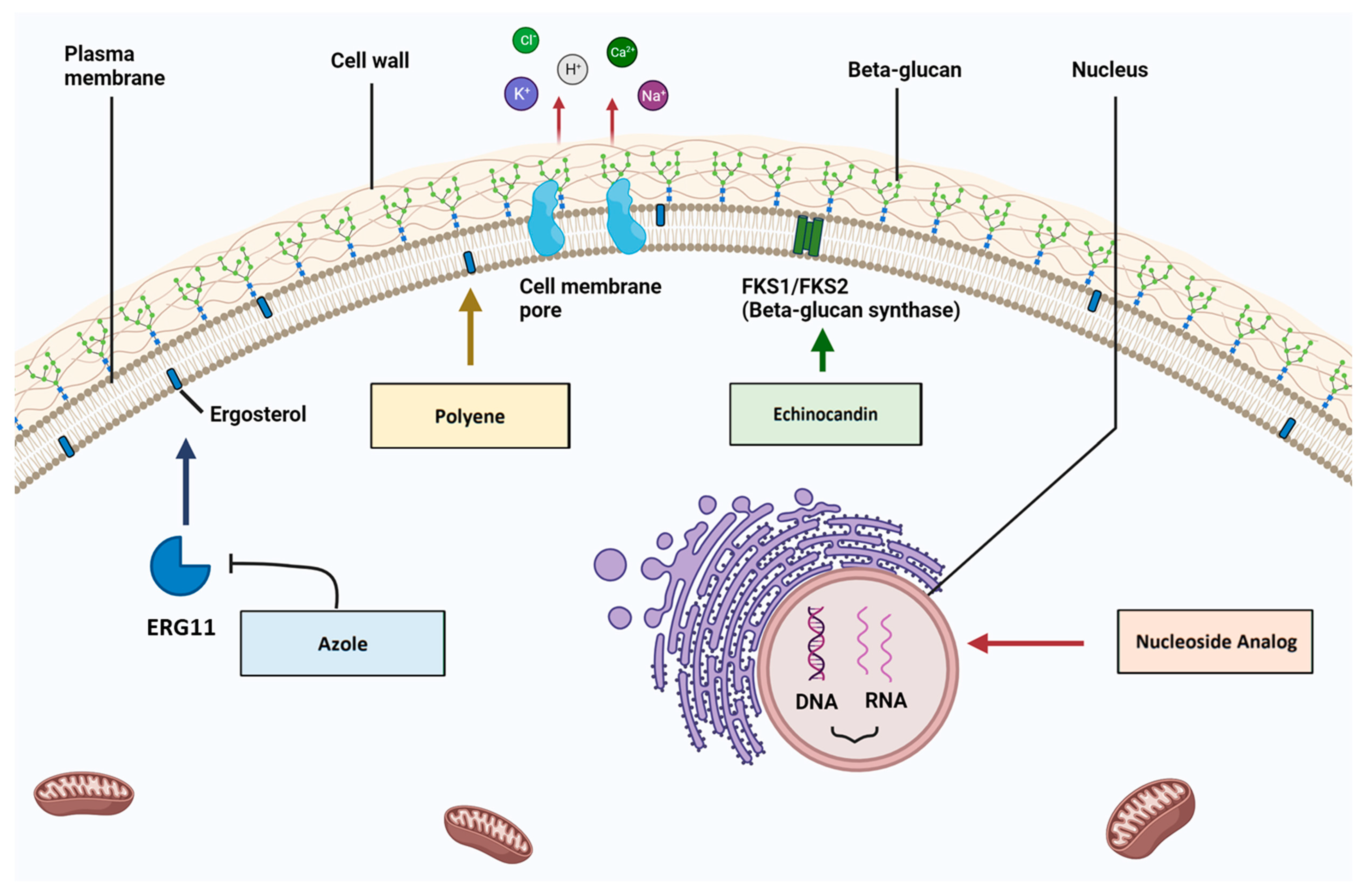
3. The Ergosterol Biosynthesis Pathway and Antifungal Resistance
3.1. ERG11
3.2. Mutations in Transcriptional Regulators
3.3. Other ERG Genes and Toxic Diol Formation
4. Cell Membrane Proteins and Antifungal Resistance
4.1. Drug Efflux Pump/Transporter Genes and Resistant Mutations
4.2. Transcriptional Regulators of Transporter Genes
4.3. Post-Translational Regulation of Transporter Genes
5. The Cell Wall Biosynthesis Pathway and Antifungal Resistance
5.1. FKS1 and FKS2 Sequence Mutations
5.2. Transcriptional Regulators of fks Genes
5.3. Protein Analysis Associated with Echinocandin Resistance
6. The Nucleic Acid Biosynthesis Pathway and Antifungal Resistance
7. Biofilm Formation and Antifungal Resistance
7.1. Biofilm Formation during Antifungal Treatment
7.2. The Roles of β-1,3 Glucan and Biofilm-Associated Antifungal Resistance
7.3. Relevant Antifungal Resistance Genes in Biofilm-Associated Candida Infections
8. Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Bhattacharya, S.; Sae-Tia, S.; Fries, B.C. Candidiasis and Mechanisms of Antifungal Resistance. Antibiotics 2020, 9, 312. [Google Scholar]
- Vázquez-González, D.; Perusquía-Ortiz, A.M.; Hundeiker, M.; Bonifaz, A. Opportunistic Yeast Infections: Candidiasis, Cryptococcosis, Trichosporonosis and Geotrichosis. JDDG J. Ger. Soc. Dermatol. 2013, 11, 381–394. [Google Scholar] [CrossRef]
- Fisher, M.C.; Gow, N.A.R.; Gurr, S.J. Tackling Emerging Fungal Threats to Animal Health, Food Security and Ecosystem Resilience. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20160332. [Google Scholar] [CrossRef] [PubMed]
- Kourkoumpetis, T.K.; Velmahos, G.C.; Ziakas, P.D.; Tampakakis, E.; Manolakaki, D.; Coleman, J.J.; Mylonakis, E. The Effect of Cumulative Length of Hospital Stay on the Antifungal Resistance of Candida Strains Isolated from Critically Ill Surgical Patients. Mycopathologia 2011, 171, 85–91. [Google Scholar] [CrossRef]
- Dąbrowska, M.; Sienkiewicz, M.; Kwiatkowski, P.; Dąbrowski, M. Diagnosis and Treatment of Mucosa Candida spp. Infections—A Review Article. Ann. Univ. Mariae Curie Sklodowska Sect. C Biol. 2019, 73, 61–68. [Google Scholar] [CrossRef]
- Gonsalves, W.C.; Wrightson, A.S.; Henry, R.G. Common Oral Conditions in Older Persons. Am. Fam. Physician 2008, 78, 845–852. [Google Scholar] [PubMed]
- Lalla, R.V.; Latortue, M.C.; Hong, C.H.; Ariyawardana, A.; D’Amato-Palumbo, S.; Fischer, D.J.; Martof, A.; Nicolatou-Galitis, O.; Patton, L.L.; Elting, L.S.; et al. A Systematic Review of Oral Fungal Infections in Patients Receiving Cancer Therapy. Support. Care Cancer 2010, 18, 985–992. [Google Scholar] [PubMed]
- Rohr, Y.; Adams, J.; Young, L. Oral Discomfort in Palliative Care: Results of an Exploratory Study of the Experiences of Terminally Ill Patients. Int. J. Palliat. Nurs. 2010, 16, 439–444. [Google Scholar] [CrossRef]
- Pfaller, M.; Neofytos, D.; Diekema, D.; Azie, N.; Meier-Kriesche, H.U.; Quan, S.P.; Horn, D. Epidemiology and Outcomes of Candidemia in 3648 Patients: Data from the Prospective Antifungal Therapy (PATH Alliance®) Registry, 2004–2008. Diagn. Microbiol. Infect. Dis. 2012, 74, 323–331. [Google Scholar] [CrossRef]
- Fisher, M.C.; Alastruey-Izquierdo, A.; Berman, J.; Bicanic, T.; Bignell, E.M.; Bowyer, P.; Bromley, M.; Brüggemann, R.; Garber, G.; Cornely, O.A.; et al. Tackling the Emerging Threat of Antifungal Resistance to Human Health. Nat. Rev. Microbiol. 2022, 20, 557–571. [Google Scholar]
- Kumar, R.; Srivastava, V. Application of Anti-Fungal Vaccines as a Tool against Emerging Anti-Fungal Resistance. Front. Fungal Biol. 2023, 4, 1241539. [Google Scholar] [CrossRef]
- Benedict, K.; Jackson, B.R.; Chiller, T.; Beer, K.D. Estimation of Direct Healthcare Costs of Fungal Diseases in the United States. Clin. Infect. Dis. 2019, 68, 1791–1797. [Google Scholar] [CrossRef]
- Al-Baqsami, Z.F.; Ahmad, S.; Khan, Z. Antifungal Drug Susceptibility, Molecular Basis of Resistance to Echinocandins and Molecular Epidemiology of Fluconazole Resistance among Clinical Candida Glabrata Isolates in Kuwait. Sci. Rep. 2020, 10, 6238. [Google Scholar] [CrossRef]
- Gupta, P.; Gupta, S.; Sharma, M.; Kumar, N.; Pruthi, V.; Poluri, K.M. Effectiveness of Phytoactive Molecules on Transcriptional Expression, Biofilm Matrix, and Cell Wall Components of Candida Glabrata and Its Clinical Isolates. ACS Omega 2018, 3, 12201–12214. [Google Scholar] [CrossRef] [PubMed]
- Jabeen, G.; Naz, S.A.; Rangel, D.E.N.; Jabeen, N.; Shafique, M.; Yasmeen, K. In-Vitro Evaluation of Virulence Markers and Antifungal Resistance of Clinical Candida Albicans Strains Isolated from Karachi, Pakistan. Fungal Biol. 2023, 127, 1241–1249. [Google Scholar] [CrossRef] [PubMed]
- Yapar, N. Epidemiology and Risk Factors for Invasive Candidiasis. Ther. Clin. Risk Manag. 2014, 10, 95–105. [Google Scholar] [CrossRef]
- World Health Organization. WHO Fungal Priority Pathogens List to Guide Research, Development and Public Health Action; World Health Organization: Geneva, Switzerland, 2022; Volume 1. [Google Scholar]
- Gabaldón, T.; Naranjo-Ortíz, M.A.; Marcet-Houben, M. Evolutionary Genomics of Yeast Pathogens in the Saccharomycotina. FEMS Yeast Res. 2016, 16, fow064. [Google Scholar] [CrossRef]
- Defosse, T.A.; Le Govic, Y.; Courdavault, V.; Clastre, M.; Vandeputte, P.; Chabasse, D.; Bouchara, J.P.; Giglioli-Guivarc’h, N.; Papon, N. Yeasts from the CTG Clade (Candida Clade): Biology, Impact in Human Health, and Biotechnological Applications. J. Mycol. Med. 2018, 28, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.A.S.; Gomes, A.C.; Santos, M.C.; Carreto, L.C.; Moura, G.R. The Genetic Code of the Fungal CTG Clade. C R. Biol. 2011, 334, 607–611. [Google Scholar] [CrossRef] [PubMed]
- Borman, A.M.; Johnson, E.M. Name Changes for Fungi of Medical Importance, 2018 to 2019. J. Clin. Microbiol. 2021, 59. [Google Scholar] [CrossRef]
- Fidel, P.L.; Vazquez, J.A.; Sobel, J.D. Candida Glabrata: Review of Epidemiology, Pathogenesis, and Clinical Disease with Comparison to C. Albicans. Clin. Microbiol. Rev. 1999, 12, 80–96. [Google Scholar] [CrossRef]
- Healey, K.R.; Ortigosa, C.J.; Shor, E.; Perlin, D.S. Genetic Drivers of Multidrug Resistance in Candida Glabrata. Front. Microbiol. 2016, 7, 1995. [Google Scholar] [CrossRef]
- CDC Antibiotic Resistance Threats in the United States; U.S. Department of Health and Human Services: Atlanta, GA, USA, 2019.
- Brandt, M.E.; Lockhart, S.R. Recent Taxonomic Developments with Candida and Other Opportunistic Yeasts. Curr. Fungal Infect. Rep. 2012, 6, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Diekema, D.J.; Gibbs, D.L.; Newell, V.A.; Ellis, D.; Tullio, V.; Rodloff, A.; Fu, W.; Ling, T.A. Results from the Artemis Disk Global Antifungal Surveillance Study, 1997 to 2007: A 10.5-Year Analysis of Susceptibilities of Candida Species to Fluconazole and Voriconazole as Determined by CLSI Standardized Disk Diffusion. J. Clin. Microbiol. 2010, 48, 1366–1377. [Google Scholar] [CrossRef] [PubMed]
- Walsh, T.J.; Groll, A.; Hiemenz, J.; Fleming, R.; Roilides, E.; Anaissie, E. Infections Due to Emerging and Uncommon Medically Important Fungal Pathogens. Clin. Microbiol. Infect. 2004, 10, 48–66. [Google Scholar] [CrossRef]
- Lockhart, S.R.; Etienne, K.A.; Vallabhaneni, S.; Farooqi, J.; Chowdhary, A.; Govender, N.P.; Colombo, A.L.; Calvo, B.; Cuomo, C.A.; Desjardins, C.A.; et al. Simultaneous Emergence of Multidrug-Resistant Candida Auris on 3 Continents Confirmed by Whole-Genome Sequencing and Epidemiological Analyses. Clin. Infect. Dis. 2017, 64, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Infection Prevention and Control. Candida auris. Public Health Ontario. Available online: https://www.publichealthontario.ca/en/Diseases-and-Conditions/Health-Care-Associated-Infections/Candida-auris (accessed on 10 October 2023).
- Centers for Disease Control and Prevention. Antifungal Susceptibility Testing and Interpretation. Available online: https://www.cdc.gov/fungal/candida-auris/c-auris-antifungal.html#print (accessed on 10 October 2023).
- Frías-De-león, M.G.; Hernández-Castro, R.; Vite-Garín, T.; Arenas, R.; Bonifaz, A.; Castañón-Olivares, L.; Acosta-Altamirano, G.; Martínez-Herrera, E. Antifungal Resistance in Candida Auris: Molecular Determinants. Antibiotics 2020, 9, 568. [Google Scholar] [CrossRef]
- CLSI. CLSI Performance Standards for Antifungal Susceptibility Testing of Yeasts, 3rd ed.; CLSI Supplement M27M44S; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2022; Volume 40. [Google Scholar]
- Wiederhold, N.P. Antifungal Susceptibility Testing: A Primer for Clinicians. Open Forum Infect. Dis. 2021, 8, ofab444. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Effron, G. Molecular Markers of Antifungal Resistance: Potential Uses in Routine Practice and Future Perspectives. J. Fungi 2021, 7, 197. [Google Scholar] [CrossRef]
- Castanheira, M.; Deshpande, L.M.; Davis, A.P.; Carvalhaes, C.G.; Pfaller, M.A. Azole Resistance in Candida Glabrata Clinical Isolates from Global Surveillance Is Associated with Efflux Overexpression. J. Glob. Antimicrob. Resist. 2022, 29, 371–377. [Google Scholar] [CrossRef]
- White, T.C.; Holleman, S.; Dy, F.; Mirels, L.F.; Stevens, D.A. Resistance Mechanisms in Clinical Isolates of Candida Albicans. Antimicrob. Agents Chemother. 2002, 46, 1704–1713. [Google Scholar] [CrossRef]
- Shields, R.K.; Nguyen, M.H.; Press, E.G.; Kwa, A.L.; Cheng, S.; Du, C.; Clancy, C.J. The Presence of an FKS Mutation Rather than MIC Is an Independent Risk Factor for Failure of Echinocandin Therapy among Patients with Invasive Candidiasis Due to Candida Glabrata. Antimicrob. Agents Chemother. 2012, 56, 4862–4869. [Google Scholar] [CrossRef]
- Bienvenu, A.L.; Leboucher, G.; Picot, S. Comparison of Fks Gene Mutations and Minimum Inhibitory Concentrations for the Detection of Candida Glabrata Resistance to Micafungin: A Systematic Review and Meta-Analysis. Mycoses 2019, 62, 835–846. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Andes, D.; Diekema, D.J.; Espinel-Ingroff, A.; Sheehan, D. Wild-Type MIC Distributions, Epidemiological Cutoff Values and Species-Specific Clinical Breakpoints for Fluconazole and Candida: Time for Harmonization of CLSI and EUCAST Broth Microdilution Methods. Drug Resist. Updates 2010, 13, 180–195. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Andes, D.; Arendrup, M.C.; Diekema, D.J.; Espinel-Ingroff, A.; Alexander, B.D.; Brown, S.D.; Chaturvedi, V.; Fowler, C.L.; Ghannoum, M.A.; et al. Clinical Breakpoints for Voriconazole and Candida Spp. Revisited: Review of Microbiologic, Molecular, Pharmacodynamic, and Clinical Data as They Pertain to the Development of Species-Specific Interpretive Criteria. Diagn. Microbiol. Infect. Dis. 2011, 70, 330–343. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Diekema, D.J.; Andes, D.; Arendrup, M.C.; Brown, S.D.; Lockhart, S.R.; Motyl, M.; Perlin, D.S. Clinical Breakpoints for the Echinocandins and Candida Revisited: Integration of Molecular, Clinical, and Microbiological Data to Arrive at Species-Specific Interpretive Criteria. Drug Resist. Updates 2011, 14, 164–176. [Google Scholar] [CrossRef] [PubMed]
- EUCAST. The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 13.0. 2023. Available online: http://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_5.0_Breakpoint_Table_01.pdf (accessed on 10 October 2023).
- Maria, S.; Barnwal, G.; Kumar, A.; Mohan, K.; Vinod, V.; Varghese, A.; Biswas, R. Species Distribution and Antifungal Susceptibility among Clinical Isolates of Candida Parapsilosis Complex from India. Rev. Iberoam. Micol. 2018, 35, 147–150. [Google Scholar] [CrossRef]
- Borman, A.M.; Muller, J.; Walsh-Quantick, J.; Szekely, A.; Patterson, Z.; Palmer, M.D.; Fraser, M.; Johnson, E.M. Fluconazole Resistance in Isolates of Uncommon Pathogenic Yeast Species from the United Kingdom. Antimicrob. Agents Chemother. 2019, 63, e00211-19. [Google Scholar] [CrossRef]
- Vigezzi, C.; Icely, P.A.; Dudiuk, C.; Rodríguez, E.; Miró, M.S.; Castillo, G.D.V.; Azcurra, A.I.; Abiega, C.; Caeiro, J.P.; Riera, F.O.; et al. Frequency, Virulence Factors and Antifungal Susceptibility of Candida Parapsilosis Species Complex Isolated from Patients with Candidemia in the Central Region of Argentina. J. Mycol. Med. 2019, 29, 285–291. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Epidemiological Cutoff Values for Antifungal Susceptibility Testing, 2nd ed.; CLSI Supplement M59; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Pfaller, M.A.; Diekema, D.J.; Turnidge, J.D.; Castanheira, M.; Jones, R.N. Twenty Years of the SENTRY Antifungal Surveillance Program: Results for Candida Species from 1997-2016. Open Forum Infect. Dis. 2019, 6, S79–S94. [Google Scholar] [CrossRef]
- Wiederhold, N.P. Antifungal Resistance: Current Trends and Future Strategies to Combat. Infect. Drug Resist. 2017, 10, 249–259. [Google Scholar] [CrossRef]
- Yesudhason, B.L.; Mohanram, K. Candida Tropicalis as a Predominant Isolate from Clinical Specimens and Its Antifungal Susceptibility Pattern in a Tertiary Care Hospital in Southern India. J. Clin. Diagn. Res. 2015, 9, DC14-6. [Google Scholar] [CrossRef]
- Kermani, F.; Taghizadeh-Armaki, M.; Hosseini, S.A.; Amirrajab, N.; Javidnia, J.; Zaghrami, M.F.; Shokohi, T. Antifungal Resistance of Clinical Candida Albicans Isolates in Iran: A Systematic Review and Meta-Analysis. Iran. J. Public Health 2023, 52, 290–305. [Google Scholar] [CrossRef] [PubMed]
- Szweda, P.; Gucwa, K.; Romanowska, E.; Dzierżanowska-Fangrat, K.; Naumiuk, Ł.; Brillowska-Dąbrowska, A.; Wojciechowska-Koszko, I.; Milewski, S. Mechanisms of Azole Resistance among Clinical Isolates of Candida Glabrata in Poland. J. Med. Microbiol. 2015, 64, 610–619. [Google Scholar] [CrossRef] [PubMed]
- Tan, T.Y.; Hsu, L.Y.; Alejandria, M.M.; Chaiwarith, R.; Chinniah, T.; Chayakulkeeree, M.; Choudhury, S.; Chen, Y.H.; Shin, J.H.; Kiratisin, P.; et al. Antifungal Susceptibility of Invasive Candida Bloodstream Isolates from the Asia-Pacific Region. Med. Mycol. 2016, 54, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Amanloo, S.; Shams-Ghahfarokhi, M.; Ghahri, M.; Razzaghi-Abyaneh, M. Drug Susceptibility Profile of Candida Glabrata Clinical Isolates from Iran and Genetic Resistant Mechanisms to Caspofungin. Rev. Iberoam. Micol. 2018, 35, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Herrera, E.; Frías-De-león, M.G.; Hernández-Castro, R.; García-Salazar, E.; Arenas, R.; Ocharan-Hernández, E.; Rodríguez-Cerdeira, C. Antifungal Resistance in Clinical Isolates of Candida Glabrata in Ibero-America. J. Fungi 2022, 8, 14. [Google Scholar] [CrossRef]
- de Tilly, A.N.; Tharmalingam, S. Review of Treatments for Oropharyngeal Fungal Infections in HIV/AIDS Patients. Microbiol. Res. 2022, 13, 219–234. [Google Scholar] [CrossRef]
- Shalini, K.; Kumar, N.; Drabu, S.; Sharma, P.K. Advances in Synthetic Approach to and Antifungal Activity of Triazoles. Beilstein J. Org. Chem. 2011, 7, 668–677. [Google Scholar] [CrossRef]
- Robbins, N.; Caplan, T.; Cowen, L.E. Molecular Evolution of Antifungal Drug Resistance. Annu. Rev. Microbiol. 2017, 71, 753–775. [Google Scholar] [CrossRef]
- Robbins, N.; Wright, G.D.; Cowen, L.E. Antifungal Drugs: The Current Armamentarium and Development of New Agents. Microbiol. Spectr. 2016, 4, 903–922. [Google Scholar] [CrossRef] [PubMed]
- Sanglard, D.; Coste, A.; Ferrari, S. Antifungal Drug Resistance Mechanisms in Fungal Pathogens from the Perspective of Transcriptional Gene Regulation. FEMS Yeast Res. 2009, 9, 1029–1050. [Google Scholar] [CrossRef]
- Azanza, J.R.; García-Quetglas, E.; Sádaba, B. Pharmacology of Azoles. Rev. Iberoam. Micol. 2007, 24, 223–227. [Google Scholar]
- Berkow, E.L.; Lockhart, S.R. Fluconazole Resistance in Candida Species: A Current Perspective. Infect. Drug Resist. 2017, 10, 237–245. [Google Scholar] [CrossRef]
- Sanguinetti, M.; Posteraro, B.; Fiori, B.; Ranno, S.; Torelli, R.; Fadda, G. Mechanisms of Azole Resistance in Clinical Isolates of Candida Glabrata Collected during a Hospital Survey of Antifungal Resistance. Antimicrob. Agents Chemother. 2005, 49, 668–679. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Diekema, D.J.; Sheehan, D.J. Interpretive Breakpoints for Fluconazole and Candida Revisited: A Blueprint for the Future of Antifungal Susceptibility Testing. Clin. Microbiol. Rev. 2006, 19, 435–447. [Google Scholar] [CrossRef]
- Castanheira, M.; Deshpande, L.M.; Davis, A.P.; Rhomberg, P.R.; Pfaller, M.A. Monitoring Antifungal Resistance in a Global Collection of Invasive Yeasts and Molds: Application of CLSI Epidemiological Cutoff Values and Whole-Genome Sequencing Analysis for Detection of Azole Resistance in Candida Albicans. Antimicrob. Agents Chemother. 2017, 61, e00906-17. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Messer, S.A.; Boyken, L.; Tendolkar, S.; Hollis, R.J.; Diekema, D.J. Geographic Variation in the Susceptibilities of Invasive Isolates of Candida Glabrata to Seven Systemically Active Antifungal Agents: A Global Assessment from the ARTEMIS Antifungal Surveillance Program Conducted in 2001 and 2002. J. Clin. Microbiol. 2004, 42, 3142–3146. [Google Scholar] [CrossRef]
- Bohner, F.; Papp, C.; Gácser, A. The Effect of Antifungal Resistance Development on the Virulence of Candida Species. FEMS Yeast Res. 2022, 22, foac019. [Google Scholar] [CrossRef]
- Efimova, S.S.; Schagina, L.V.; Ostroumova, O.S. Investigation of Channel-Forming Activity of Polyene Macrolide Antibiotics in Planar Lipid Bilayers in the Presence of Dipole Modifiers. Acta Naturae 2014, 6, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Birch, M.; Sibley, G. Antifungal Chemistry Review. In Comprehensive Medicinal Chemistry III; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Waller, D.G.; Sampson, A.P. Chemotherapy of Infections. In Medical Pharmacology and Therapeutics; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Espinel-Ingroff, A.; Arendrup, M.; Canton, E.; Cordob, S.; Dannaoui, E.; Garcia-Rodriguez, J.; Gonzalez, G.M.; Govender, N.P.; Martin-Mazuelos, E.; Lackner, M.; et al. Multicenter Study of Method-Dependent Epidemiological Cutoff Values for Detection of Resistance in Candida Spp. and Aspergillus Spp. to Amphotericin B and Echinocandins for the Etest Agar Diffusion Method. Antimicrob. Agents Chemother. 2017, 61, e01792-16. [Google Scholar] [CrossRef] [PubMed]
- Kanafani, Z.A.; Perfect, J.R. Resistance to Antifungal Agents: Mechanisms and Clinical Impact. Clin. Infect. Dis. 2008, 46, 120–128. [Google Scholar] [CrossRef]
- Vitiello, A.; Ferrara, F.; Boccellino, M.; Ponzo, A.; Cimmino, C.; Comberiati, E.; Zovi, A.; Clemente, S.; Sabbatucci, M. Antifungal Drug Resistance: An Emergent Health Threat. Biomedicines 2023, 11, 1063. [Google Scholar] [CrossRef]
- Popolo, L.; Gualtieri, T.; Ragni, E. The Yeast Cell-Wall Salvage Pathway. Med. Mycol. Suppl. 2001, 39, 111–121. [Google Scholar] [CrossRef]
- Walker, L.A.; Gow, N.A.R.; Munro, C.A. Fungal Echinocandin Resistance. Fungal Genet. Biol. 2010, 47, 117–126. [Google Scholar] [CrossRef]
- Douglas, C.M.; D’Ippolito, J.A.; Shei, G.J.; Meinz, M.; Onishi, J.; Marrinan, J.A.; Li, W.; Abruzzo, G.K.; Flattery, A.; Bartizal, K.; et al. Identification of the FKS1 Gene of Candida Albicans as the Essential Target of 1,3-β-D-Glucan Synthase Inhibitors. Antimicrob. Agents Chemother. 1997, 41, 2471–2479. [Google Scholar] [CrossRef]
- Grover, N. Echinocandins: A Ray of Hope in Antifungal Drug Therapy. Indian. J. Pharmacol. 2010, 42, 9–11. [Google Scholar] [CrossRef] [PubMed]
- Barchiesi, F.; Spreghini, E.; Tomassetti, S.; Della Vittoria, A.; Arzeni, D.; Manso, E.; Scalise, G. Effects of Caspofungin against Candida Guilliermondii and Candida Parapsilosis. Antimicrob. Agents Chemother. 2006, 50, 2719–2727. [Google Scholar] [CrossRef] [PubMed]
- Cantón, E.; Pemán, J.; Sastre, M.; Romero, M.; Espinel-Ingroff, A. Killing Kinetics of Caspofungin, Micafungin, and Amphotericin B against Candida Guilliermondii. Antimicrob. Agents Chemother. 2006, 50, 2829–2832. [Google Scholar] [CrossRef] [PubMed]
- Arastehfar, A.; Lass-Flörl, C.; Garcia-Rubio, R.; Daneshnia, F.; Ilkit, M.; Boekhout, T.; Gabaldon, T.; Perlin, D.S. The Quiet and Underappreciated Rise of Drug-Resistant Invasive Fungal Pathogens. J. Fungi 2020, 6, 138. [Google Scholar] [CrossRef] [PubMed]
- Vermes, A.; Guchelaar, H.J.; Dankert, J. Flucytosine: A Review of Its Pharmacology, Clinical Indications, Pharmacokinetics, Toxicity and Drug Interactions. J. Antimicrob. Chemother. 2000, 46, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Mourad, A.; Perfect, J.R. Present and Future Therapy of Cryptococcus Infections. J. Fungi 2018, 4, 79. [Google Scholar] [CrossRef]
- Perfect, J.R.; Dismukes, W.E.; Dromer, F.; Goldman, D.L.; Graybill, J.R.; Hamill, R.J.; Harrison, T.S.; Larsen, R.A.; Lortholary, O.; Nguyen, M.H.; et al. Clinical Practice Guidelines for the Management of Cryptococcal Disease: 2010 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2010, 50, 291–322. [Google Scholar] [CrossRef]
- Delma, F.Z.; Al-Hatmi, A.M.S.; Brüggemann, R.J.M.; Melchers, W.J.G.; de Hoog, S.; Verweij, P.E.; Buil, J.B. Molecular Mechanisms of 5-Fluorocytosine Resistance in Yeasts and Filamentous Fungi. J. Fungi 2021, 7, 909. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Messer, S.A.; Boyken, L.; Huynh, H.; Hollis, R.J.; Diekema, D.J. In Vitro Activities of 5-Fluorocytosine against 8803 Clinical Isolates of Candida spp.: Global Assessment of Primary Resistance Using National Committee for Clinical Laboratory Standards Susceptibility Testing Methods. Antimicrob. Agents Chemother. 2002, 46, 3518–3521. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Esquivel, B.D.; White, T.C. Overexpression or Deletion of Ergosterol Biosynthesis Genes Alters Doubling Time, Response to Stress Agents, and Drug Susceptibility in Saccharomyces Cerevisiae. mBio 2018, 9, e01291-18. [Google Scholar] [CrossRef] [PubMed]
- Xiang, M.J.; Liu, J.Y.; Ni, P.H.; Wang, S.; Shi, C.; Wei, B.; Ni, Y.X.; Ge, H.L. Erg11 Mutations Associated with Azole Resistance in Clinical Isolates of Candida Albicans. FEMS Yeast Res. 2013, 13, 386–393. [Google Scholar] [CrossRef]
- Manastir, L.; Ergon, M.C.; Yücesoy, M. Investigation of Mutations in Erg11 Gene of Fluconazole Resistant Candida Albicans Isolates from Turkish Hospitals. Mycoses 2011, 54, 99–104. [Google Scholar] [CrossRef]
- Morio, F.; Loge, C.; Besse, B.; Hennequin, C.; Le Pape, P. Screening for Amino Acid Substitutions in the Candida Albicans Erg11 Protein of Azole-Susceptible and Azole-Resistant Clinical Isolates: New Substitutions and a Review of the Literature. Diagn. Microbiol. Infect. Dis. 2010, 66, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.J.; Wan, Z.; Wang, X.H.; Li, R.Y.; Liu, W. Relationship between Antifungal Resistance of Fluconazole Resistant Candida Albicans and Mutations in ERG11 Gene. Chin. Med. J. 2010, 123, 544–548. [Google Scholar]
- Xu, Y.; Chen, L.; Li, C. Susceptibility of Clinical Isolates of Candida Species to Fluconazole and Detection of Candida Albicans ERG11 Mutations. J. Antimicrob. Chemother. 2008, 61, 798–804. [Google Scholar] [CrossRef]
- Chen, S.H.; Sheng, C.Q.; Xu, X.H.; Jiang, Y.Y.; Zhang, W.N.; He, C. Identification of Y118 Amino Acid Residue in Candida Albicans Sterol 14α-Demethylase Associated with the Enzyme Activity and Selective Antifungal Activity of Azole Analogues. Biol. Pharm. Bull. 2007, 30, 1246–1253. [Google Scholar] [CrossRef]
- Lamb, D.C.; Kelly, D.E.; Schunck, W.H.; Shyadehi, A.Z.; Akhtar, M.; Lowe, D.J.; Baldwin, B.C.; Kelly, S.L. The Mutation T315A in Candida Albicans Sterol 14α-Demethylase Causes Reduced Enzyme Activity and Fluconazole Resistance through Reduced Affinity. J. Biol. Chem. 1997, 272, 5682–5688. [Google Scholar] [CrossRef]
- Li, X.; Brown, N.; Chau, A.S.; López-Ribot, J.L.; Ruesga, M.T.; Quindos, G.; Mendrick, C.A.; Hare, R.S.; Loebenberg, D.; DiDomenico, B.; et al. Changes in Susceptibility to Posaconazole in Clinical Isolates of Candida Albicans. J. Antimicrob. Chemother. 2004, 53, 74–80. [Google Scholar] [CrossRef]
- dos Santos Silva, D.B.; Carbonera Rodrigues, L.M.; De Almeida, A.A.; de Oliveira, K.M.P.; Grisolia, A.B. Novel Point Mutations in the ERG11 Gene in Clinical Isolates of Azole Resistant Candida Species. Mem. Inst. Oswaldo Cruz 2016, 111, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Berila, N.; Subik, J. Molecular Analysis of Candida Glabrata Clinical Isolates. Mycopathologia 2010, 170, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Berila, N.; Borecka, S.; Dzugasova, V.; Bojnansky, J.; Subik, J. Mutations in the CgPDR1 and CgERG11 Genes in Azole-Resistant Candida Glabrata Clinical Isolates from Slovakia. Int. J. Antimicrob. Agents 2009, 33, 574–578. [Google Scholar] [CrossRef]
- Vandeputte, P.; Larcher, G.; Bergès, T.; Renier, G.; Chabasse, D.; Bouchara, J.P. Mechanisms of Azole Resistance in a Clinical Isolate of Candida Tropicalis. Antimicrob. Agents Chemother. 2005, 49, 4608–4615. [Google Scholar] [CrossRef] [PubMed]
- Xisto, M.I.D.S.; Caramalho, R.D.F.; Rocha, D.A.S.; Ferreira-Pereira, A.; Sartori, B.; Barreto-Bergter, E.; Junqueira, M.L.; Lass-Flörl, C.; Lackner, M. Pan-Azole-Resistant Candida Tropicalis Carrying Homozygous Erg11 Mutations at Position K143R: A New Emerging Superbug? J. Antimicrob. Chemother. 2017, 72, 988–992. [Google Scholar] [CrossRef]
- Yoo, J.I.; Choi, C.W.; Lee, K.M.; Lee, Y.S. Gene Expression and Identification Related to Fluconazole Resistance of Candida Glabrata Strains. Osong Public. Health Res. Perspect. 2010, 1, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Flowers, S.A.; Barker, K.S.; Berkow, E.L.; Toner, G.; Chadwick, S.G.; Gygax, S.E.; Morschhäuser, J.; David Rogers, P. Gain-of-Function Mutations in UPC2 Are a Frequent Cause of ERG11 Upregulation in Azole-Resistant Clinical Isolates of Candida Albicans. Eukaryot. Cell 2012, 11, 1289–1299. [Google Scholar] [CrossRef] [PubMed]
- Dunkel, N.; Liu, T.T.; Barker, K.S.; Homayouni, R.; Morschhäuser, J.; Rogers, P.D. A Gain-of-Function Mutation in the Transcription Factor Upc2p Causes Upregulation of Ergosterol Biosynthesis Genes and Increased Fluconazole Resistance in a Clinical Candida Albicans Isolate. Eukaryot. Cell 2008, 7, 1180–1190. [Google Scholar] [CrossRef] [PubMed]
- Chau, A.S.; Mendrick, C.A.; Sabatelli, F.J.; Loebenberg, D.; McNicholas, P.M. Application of Real-Time Quantitative PCR to Molecular Analysis of Candida Albicans Strains Exhibiting Reduced Susceptibility to Azoles. Antimicrob. Agents Chemother. 2004, 48, 2124–2131. [Google Scholar] [CrossRef] [PubMed]
- Sanglard, D.; Ischer, F.; Calabrese, D.; Micheli, M.d.; Bille, J. Multiple Resistance Mechanisms to Azole Antifungals in Yeast Clinical Isolates. Drug Resist. Updates 1998, 1, 255–265. [Google Scholar] [CrossRef]
- Sanglard, D.; Ischer, F.; Koymans, L.; Bille, J. Amino Acid Substitutions in the Cytochrome P-450 Lanosterol 14α- Demethylase (CYP51A1) from Azole-Resistant Candida Albicans Clinical Isolates Contribute to Resistance to Azole Antifungal Agents. Antimicrob. Agents Chemother. 1998, 42, 241–253. [Google Scholar] [CrossRef]
- Marichal, P.; Koymans, L.; Willemsens, S.; Bellens, D.; Verhasselt, P.; Luyten, W.; Borgers, M.; Ramaekers, F.C.S.; Odds, F.C.; Bossche, H. Vanden Contribution of Mutations in the Cytochrome P450 14α-Demethylase (Erg11p, Cyp51p) to Azole Resistance in Candida Albicans. Microbiol. 1999, 145, 2701–2713. [Google Scholar] [CrossRef]
- Akins, R.A. An Update on Antifungal Targets and Mechanisms of Resistance in Candida Albicans. Med. Mycol. 2005, 43, 285–318. [Google Scholar] [CrossRef] [PubMed]
- Hoot, S.J.; Smith, A.R.; Brown, R.P.; White, T.C. An A643V Amino Acid Substitution in Upc2p Contributes to Azole Resistance in Well-Characterized Clinical Isolates of Candida Albicans. Antimicrob. Agents Chemother. 2011, 55, 940–942. [Google Scholar] [CrossRef]
- Heilmann, C.J.; Schneider, S.; Barker, K.S.; Rogers, P.D.; Morschhäuser, J. An A643T Mutation in the Transcription Factor Upc2p Causes Constitutive ERG11 Upregulation and Increased Fluconazole Resistance in Candida Albicans. Antimicrob. Agents Chemother. 2010, 54, 353–359. [Google Scholar] [CrossRef]
- Löffler, J.; Kelly, S.L.; Hebart, H.; Schumacher, U.; Lass-Flörl, C.; Einsele, H. Molecular Analysis of Cyp51 from Fluconazole-Resistant Candida Albicans Strains. FEMS Microbiol. Lett. 1997, 151, 263–268. [Google Scholar] [CrossRef]
- Perea, S.; López-Ribot, J.L.; Kirkpatrick, W.R.; McAtee, R.K.; Santillán, R.A.; Martínez, M.; Calabrese, D.; Sanglard, D.; Patterson, T.F. Prevalence of Molecular Mechanisms of Resistance to Azole Antifungal Agents in Candida Albicans Strains Displaying High-Level Fluconazole Resistance Isolated from Human Immunodeficiency Virus-Infected Patients. Antimicrob. Agents Chemother. 2001, 45, 2676–2684. [Google Scholar] [CrossRef] [PubMed]
- Znaidi, S.; Weber, S.; Al-Abdin, O.Z.; Bomme, P.; Saidane, S.; Drouin, S.; Lemieux, S.; De Deken, X.; Robert, F.; Raymond, M. Genomewide Location Analysis of Candida Albicans Upc2p, a Regulator of Sterol Metabolism and Azole Drug Resistance. Eukaryot. Cell 2008, 7, 836–847. [Google Scholar] [CrossRef]
- Barker, K.S.; Crisp, S.; Wiederhold, N.; Lewis, R.E.; Bareither, B.; Eckstein, J.; Barbuch, R.; Bard, M.; Rogers, P.D. Genome-Wide Expression Profiling Reveals Genes Associated with Amphotericin B and Fluconazole Resistance in Experimentally Induced Antifungal Resistant Isolates of Candida Albicans. J. Antimicrob. Chemother. 2004, 54, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Kelly, S.L.; Lamb, D.C.; Corran, A.J.; Baldwin, B.C.; Kelly, D.E. Mode of Action and Resistance to Azole Antifungals Associated with the Formation of 14α-Methylergosta-8,24(28)-Dien-3β,6α-Diol. Biochem. Biophys. Res. Commun. 1995, 207, 910–915. [Google Scholar] [CrossRef]
- Watson, P.F.; Rose, M.E.; Ellis, S.W.; England, H.; Kelly, S.L. Defective Sterol C5-6 Desaturation and Azole Resistance: A New Hypothesis for the Mode of Action of Azole Antifungals. Biochem. Biophys. Res. Commun. 1989, 164, 1170–1175. [Google Scholar] [CrossRef] [PubMed]
- Sanglard, D.; Ischer, F.; Parkinson, T.; Falconer, D.; Bille, J. Candida Albicans Mutations in the Ergosterol Biosynthetic Pathway and Resistance to Several Antifungal Agents. Antimicrob. Agents Chemother. 2003, 47, 2404–2412. [Google Scholar] [CrossRef] [PubMed]
- Whaley, S.G.; Berkow, E.L.; Rybak, J.M.; Nishimoto, A.T.; Barker, K.S.; Rogers, P.D. Azole Antifungal Resistance in Candida Albicans and Emerging Non-Albicans Candida Species. Front. Microbiol. 2017, 7, 2173. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.B.; Sirjusingh, C.; Parsons, A.B.; Boone, C.; Wickens, C.; Cowen, L.E.; Kohn, L.M. Mode of Selection and Experimental Evolution of Antifungal Drug Resistance in Saccharomyces Cerevisiae. Genetics 2003, 163, 1287–1298. [Google Scholar] [CrossRef]
- Xu, D.; Jiang, B.; Ketela, T.; Lemieux, S.; Veillette, K.; Martel, N.; Davison, J.; Sillaots, S.; Trosok, S.; Bachewich, C.; et al. Genome-Wide Fitness Test and Mechanism-of-Action Studies of Inhibitory Compounds in Candida Albicans. PLoS Pathog. 2007, 3, e92. [Google Scholar] [CrossRef]
- Lewis, R.E.; Viale, P.; Kontoyiannis, D.P. The Potential Impact of Antifungal Drug Resistance Mechanisms on the Host Immune Response to Candida. Virulence 2012, 3, 368–376. [Google Scholar] [CrossRef]
- Jia, Y.; Tang, R.J.; Wang, L.; Zhang, X.; Wang, Y.; Jia, X.M.; Jiang, Y.Y. Calcium-Activated-Calcineurin Reduces the In Vitro and In Vivo Sensitivity of Fluconazole to Candida Albicans via Rta2p. PLoS ONE 2012, 7, e48369. [Google Scholar] [CrossRef] [PubMed]
- Prasad, R.; Nair, R.; Banerjee, A. Multidrug Transporters of Candida Species in Clinical Azole Resistance. Fungal Genet. Biol. 2019, 132, 103252. [Google Scholar] [CrossRef]
- Sanglard, D.; Ischer, F.; Monod, M.; Bille, J. Susceptibilities of Candida Albicans Multidrug Transporter Mutants to Various Antifungal Agents and Other Metabolic Inhibitors. Antimicrob. Agents Chemother. 1996, 40, 2300–2305. [Google Scholar] [CrossRef] [PubMed]
- Khandelwal, N.K.; Wasi, M.; Nair, R.; Gupta, M.; Kumar, M.; Mondal, A.K.; Gaur, N.A.; Prasad, R. Vacuolar Sequestration of Azoles, a Novel Strategy of Azole Antifungal Resistance Conserved across Pathogenic and Nonpathogenic Yeast. Antimicrob. Agents Chemother. 2019, 63, e01347-18. [Google Scholar] [CrossRef]
- Perepnikhatka, V.; Fischer, F.J.; Niimi, M.; Baker, R.A.; Cannon, R.D.; Wang, Y.K.; Sherman, F.; Rustchenko, E. Specific Chromosome Alterations in Fluconazole-Resistant Mutants of Candida Albicans. J. Bacteriol. 1999, 181, 4041–4049. [Google Scholar] [CrossRef] [PubMed]
- Coste, A.; Turner, V.; Ischer, F.; Morschhäuser, J.; Forche, A.; Selmecki, A.; Berman, J.; Bille, J.; Sanglard, D. A Mutation in Tac1p, a Transcription Factor Regulating CDR1 and CDR2, Is Coupled with Loss of Heterozygosity at Chromosome 5 to Mediate Antifungal Resistance in Candida Albicans. Genetics 2006, 172, 2139–2156. [Google Scholar] [CrossRef]
- Rybak, J.M.; Muñoz, J.F.; Barker, K.S.; Parker, J.E.; Esquivel, B.D.; Berkow, E.L.; Lockhart, S.R.; Gade, L.; Palmer, G.E.; White, T.C.; et al. Mutations in TAC1B: A Novel Genetic Determinant of Clinical Fluconazole Resistance in Candida Auris. mBio 2020, 11, e00365-20. [Google Scholar] [CrossRef]
- Kalkandelen, K.T.; Doluca Dereli, M. Investigation of Mutations in Transcription Factors of Efflux Pump Genes in Fluconazole-Resistant Candida Albicans Strains Overexpressing the Efflux Pumps. Mikrobiyol. Bul. 2015, 49, 609–618. [Google Scholar] [CrossRef]
- Schubert, S.; Rogers, P.D.; Morschhäuser, J. Gain-of-Function Mutations in the Transcription Factor MRR1 Are Responsible for Overexpression of the MDR1 Efflux Pump in Fluconazole-Resistant Candida Dubliniensis Strains. Antimicrob. Agents Chemother. 2008, 52, 4274–4280. [Google Scholar] [CrossRef]
- Moran, G.P.; Sullivan, D.J.; Henman, M.C.; McCreary, C.E.; Harrington, B.J.; Shanley, D.B.; Coleman, D.C. Antifungal Drug Susceptibilities of Oral Candida Dubliniensis Isolates from Human Immunodeficiency Virus (HIV)-Infected and Non-HIV-Infected Subjects and Generation of Stable Fluconazole-Resistant Derivatives in Vitro. Antimicrob. Agents Chemother. 1997, 41, 617–623. [Google Scholar] [CrossRef]
- Moran, G.P.; Sanglard, D.; Donnelly, S.M.; Shanley, D.B.; Sullivan, D.J.; Coleman, D.C. Identification and Expression of Multidrug Transporters Responsible for Fluconazole Resistance in Candida Dubliniensis. Antimicrob. Agents Chemother. 1998, 42, 1819–1830. [Google Scholar] [CrossRef] [PubMed]
- Bencova, A.; Goffa, E.; Morvova, M.; Valachovic, M.; Griač, P.; Toth Hervay, N.; Gbelska, Y. The Absence of PDR16 Gene Restricts the Overexpression of CaSNQ2 Gene in the Presence of Fluconazole in Candida Albicans. Mycopathologia 2020, 185, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Ischer, F.; Calabrese, D.; Posteraro, B.; Sanguinetti, M.; Fadda, G.; Rohde, B.; Bauser, C.; Bader, O.; Sanglard, D. Gain of Function Mutations in CgPDR1 of Candida Glabrata Not Only Mediate Antifungal Resistance but Also Enhance Virulence. PLoS Pathog. 2009, 5, e1000268. [Google Scholar] [CrossRef]
- Znaidi, S.; De Deken, X.; Weber, S.; Rigby, T.; Nantel, A.; Raymond, M. The Zinc Cluster Transcription Factor Tac1p Regulates PDR16 Expression in Candida Albicans. Mol. Microbiol. 2007, 66, 440–452. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, H.; Miyazaki, Y.; Geber, A.; Parkinson, T.; Hitchcock, C.; Falconer, D.J.; Ward, D.J.; Marsden, K.; Bennett, J.E. Fluconazole Resistance Associated with Drug Efflux and Increased Transcription of a Drug Transporter Gene, PDH1, in Candida Glabrata. Antimicrob. Agents Chemother. 1998, 42, 1695–1701. [Google Scholar] [CrossRef]
- Whaley, S.G.; Zhang, Q.; Caudle, K.E.; Rogers, P.D. Relative Contribution of the ABC Transporters Cdr1, Pdh1, and Snq2 to Azole Resistance in Candida Glabrata. Antimicrob. Agents Chemother. 2018, 62, e01070-18. [Google Scholar] [CrossRef] [PubMed]
- Costa, C.; Dias, P.J.; Sá-Correia, I.; Teixeira, M.C. MFS Multidrug Transporters in Pathogenic Fungi: Do They Have Real Clinical Impact? Front. Physiol. 2014, 5, 197. [Google Scholar] [CrossRef]
- Costa, C.; Ribeiro, J.; Miranda, I.M.; Silva-Dias, A.; Cavalheiro, M.; Costa-de-Oliveira, S.; Rodrigues, A.G.; Teixeira, M.C. Clotrimazole Drug Resistance in Candida Glabrata Clinical Isolates Correlates with Increased Expression of the Drug: H+ Antiporters CgAqr1, CgTpo1_1, CgTpo3, and CgQdr2. Front. Microbiol. 2016, 7, 526. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Friesa, B.C. Enhanced Efflux Pump Activity in Old Candida Glabrata Cells. Antimicrob. Agents Chemother. 2018, 62, e02227-17. [Google Scholar] [CrossRef]
- Culakova, H.; Dzugasova, V.; Valencikova, R.; Gbelska, Y.; Subik, J. Stress Response and Expression of Fluconazole Resistance Associated Genes in the Pathogenic Yeast Candida Glabrata Deleted in the CgPDR16 Gene. Microbiol. Res. 2015, 174, 17–23. [Google Scholar] [CrossRef]
- Hampe, I.A.I.; Friedman, J.; Edgerton, M.; Morschhäuser, J. An Acquired Mechanism of Antifungal Drug Resistance Simultaneously Enables Candida Albicans to Escape from Intrinsic Host Defenses. PLoS Pathog. 2017, 13, e1006655. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xiao, M.; Watts, M.R.; Wang, H.; Fan, X.; Kong, F.; Xu, Y.C. Development of Fluconazole Resistance in a Series of Candida Parapsilosis Isolates from a Persistent Candidemia Patient with Prolonged Antifungal Therapy. BMC Infect. Dis. 2015, 15, 340. [Google Scholar] [CrossRef] [PubMed]
- Pinjon, E.; Moran, G.P.; Coleman, D.C.; Sullivan, D.J. Azole Susceptibility and Resistance in Candida Dubliniensis. Biochem. Soc. Trans. 2005, 33, 1210–1214. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Sobel, J.D.; White, T.C. A Combination Fluorescence Assay Demonstrates Increased Efflux Pump Activity as a Resistance Mechanism in Azole-Resistant Vaginal Candida Albicans Isolates. Antimicrob. Agents Chemother. 2016, 60, 5858–5866. [Google Scholar] [CrossRef]
- Morschhäuser, J.; Barker, K.S.; Liu, T.T.; Blaß-Warmuth, J.; Homayouni, R.; Rogers, P.D. The Transcription Factor Mrr1p Controls Expression of the MDR1 Efflux Pump and Mediates Multidrug Resistance in Candida Albicans. PLoS Pathog. 2007, 3, e164. [Google Scholar] [CrossRef]
- Schubert, S.; Barker, K.S.; Znaidi, S.; Schneider, S.; Dierolf, F.; Dunkel, N.; Aïd, M.; Boucher, G.; Rogers, P.D.; Raymond, M.; et al. Regulation of Efflux Pump Expression and Drug Resistance by the Transcription Factors Mrr1, Upc2, and Cap1 in Candida Albicans. Antimicrob. Agents Chemother. 2011, 55, 2212–2223. [Google Scholar] [CrossRef]
- Dunkel, N.; Blaß, J.; Rogers, P.D.; Morschhäuser, J. Mutations in the Multi-Drug Resistance Regulator MRR1, Followed by Loss of Heterozygosity, Are the Main Cause of MDR1 Overexpression in Fluconazole-Resistant Candida Albicans Strains. Mol. Microbiol. 2008, 69, 827–840. [Google Scholar] [CrossRef]
- Shukla, S.; Yadav, V.; Mukhopadhyay, G.; Prasad, R. Ncb2 Is Involved in Activated Transcription of CDR1 in Azole-Resistant Clinical Isolates of Candida Albicans ∇. Eukaryot. Cell 2011, 10, 1357–1366. [Google Scholar] [CrossRef]
- Mogavero, S.; Tavanti, A.; Senesi, S.; Rogers, P.D.; Morschhäuser, J. Differential Requirement of the Transcription Factor Mcm1 for Activation of the Candida Albicans Multidrug Efflux Pump MDR1 by Its Regulators Mrr1 and Cap1. Antimicrob. Agents Chemother. 2011, 55, 2061–2066. [Google Scholar] [CrossRef]
- Alarco, A.M.; Raymond, M. The BZip Transcription Factor Cap1p Is Involved in Multidrug Resistance and Oxidative Stress Response in Candida Albicans. J. Bacteriol. 1999, 181, 700–708. [Google Scholar] [CrossRef]
- Ni, Q.; Wang, C.; Tian, Y.; Dong, D.; Jiang, C.; Mao, E.; Peng, Y. CgPDR1 Gain-of-Function Mutations Lead to Azole-Resistance and Increased Adhesion in Clinical Candida Glabrata Strains. Mycoses 2018, 61, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Vermitsky, J.P.; Earhart, K.D.; Smith, W.L.; Homayouni, R.; Edlind, T.D.; Rogers, P.D. Pdr1 Regulates Multidrug Resistance in Candida Glabrata: Gene Disruption and Genome-Wide Expression Studies. Mol. Microbiol. 2006, 61, 704–722. [Google Scholar] [CrossRef]
- Vermitsky, J.P.; Edlind, T.D. Azole Resistance in Candida Glabrata: Coordinate Upregulation of Multidrug Transporters and Evidence for a Pdr1-like Transcription Factor. Antimicrob. Agents Chemother. 2004, 48, 3773–3781. [Google Scholar] [CrossRef]
- Thomas, E.; Roman, E.; Claypool, S.; Manzoor, N.; Pla, J.; Panwar, S.L. Mitochondria Influence CDR1 Efflux Pump Activity, Hog1-Mediated Oxidative Stress Pathway, Iron Homeostasis, and Ergosterol Levels in Candida Albicans. Antimicrob. Agents Chemother. 2013, 57, 5580–5599. [Google Scholar] [CrossRef]
- Manoharlal, R.; Gorantala, J.; Sharma, M.; Sanglard, D.; Prasad, R. PAP1 [Poly(A) Polymerase 1] Homozygosity and Hyperadenylation Are Major Determinants of Increased MRNA Stability of CDR1 in Azole-Resistant Clinical Isolates of Candida Albicans. Microbiol. 2010, 156, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Latgé, J.P. The Cell Wall: A Carbohydrate Armour for the Fungal Cell. Mol. Microbiol. 2007, 66, 279–290. [Google Scholar] [CrossRef]
- Alexander, B.D.; Johnson, M.D.; Pfeiffer, C.D.; Jiménez-Ortigosa, C.; Catania, J.; Booker, R.; Castanheira, M.; Messer, S.A.; Perlin, D.S.; Pfaller, M.A. Increasing Echinocandin Resistance in Candida Glabrata: Clinical Failure Correlates with Presence of FKS Mutations and Elevated Minimum Inhibitory Concentrations. Clin. Infect. Dis. 2013, 56, 1724–1732. [Google Scholar] [CrossRef] [PubMed]
- Cleary, J.D.; Garcia-Effron, G.; Chapman, S.W.; Perlin, D.S. Reduced Candida Glabrata Susceptibility Secondary to an FKS1 Mutation Developed during Candidemia Treatment. Antimicrob. Agents Chemother. 2008, 52, 2263–2265. [Google Scholar] [CrossRef]
- Garcia-Effron, G.; Lee, S.; Park, S.; Cleary, J.D.; Perlin, D.S. Effect of Candida Glabrata FKS1 and FKS2 Mutations on Echinocandin Sensitivity and Kinetics of 1,3-β-D-Glucan Synthase: Implication for the Existing Susceptibility Breakpoint. Antimicrob. Agents Chemother. 2009, 53, 3690–3699. [Google Scholar] [CrossRef]
- Garcia-Effron, G.; Chua, D.J.; Tomada, J.R.; DiPersio, J.; Perlin, D.S.; Ghannoum, M.; Bonilla, H. Novel FKS Mutations Associated with Echinocandin Resistance in Candida Species. Antimicrob. Agents Chemother. 2010, 54, 2225–2227. [Google Scholar] [CrossRef]
- Thompson, G.R.; Wiederhold, N.P.; Vallor, A.C.; Villareal, N.C.; Lewis, J.S.; Patterson, T.F. Development of Caspofungin Resistance Following Prolonged Therapy for Invasive Candidiasis Secondary to Candida Glabrata Infection. Antimicrob. Agents Chemother. 2008, 52, 3783–3785. [Google Scholar] [CrossRef]
- Garcia-Effron, G.; Katiyar, S.K.; Park, S.; Edlind, T.D.; Perlin, D.S. A Naturally Occurring Proline-to-Alanine Amino Acid Change in Fks1p in Candida Parapsilosis, Candida Orthopsilosis, and Candida Metapsilosis Accounts for Reduced Echinocandin Susceptibility. Antimicrob. Agents Chemother. 2008, 52, 2305–2312. [Google Scholar] [CrossRef] [PubMed]
- Munro, C.A. Fungal Echinocandin Resistance. F1000 Biol. Rep. 2010, 2, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kelly, R.; Kahn, J.N.; Robles, J.; Hsu, M.J.; Register, E.; Li, W.; Vyas, V.; Fan, H.; Abruzzo, G.; et al. Specific Substitutions in the Echinocandin Target Fks1p Account for Reduced Susceptibility of Rare Laboratory and Clinical Candida sp. Isolates. Antimicrob. Agents Chemother. 2005, 49, 3264–3273. [Google Scholar] [CrossRef] [PubMed]
- Perlin, D.S. Resistance to Echinocandin-Class Antifungal Drugs. Drug Resist. Updates 2007, 10, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Hakki, M.; Staab, J.F.; Marr, K.A. Emergence of a Candida Krusei Isolate with Reduced Susceptibility to Caspofungin during Therapy. Antimicrob. Agents Chemother. 2006, 50, 2522–2524. [Google Scholar] [CrossRef]
- Kahn, J.N.; Garcia-Effron, G.; Hsu, M.J.; Park, S.; Marr, K.A.; Perlin, D.S. Acquired Echinocandin Resistance in a Candida Krusei Isolate Due to Modification of Glucan Synthase. Antimicrob. Agents Chemother. 2007, 51, 1876–1878. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Effron, G.; Park, S.; Perlin, D.S. Correlating Echinocandin MIC and Kinetic Inhibition of Fks1 Mutant Glucan Synthases for Candida Albicans: Implications for Interpretive Breakpoints. Antimicrob. Agents Chemother. 2009, 53, 112–122. [Google Scholar] [CrossRef]
- Sharma, D.; Paul, R.A.; Rudramurthy, S.M.; Kashyap, N.; Bhattacharya, S.; Soman, R.; Shankarnarayan, S.A.; Chavan, D.; Singh, S.; Das, P.; et al. Impact of FKS1 Genotype on Echinocandin In Vitro Susceptibility in Candida Auris and In Vivo Response in a Murine Model of Infection. Antimicrob. Agents Chemother. 2022, 66, e0165221. [Google Scholar] [CrossRef]
- Balashov, S.V.; Park, S.; Perlin, D.S. Assessing Resistance to the Echinocandin Antifungal Drug Caspofungin in Candida Albicans by Profiling Mutations in FKS1. Antimicrob. Agents Chemother. 2006, 50, 2058–2063. [Google Scholar] [CrossRef]
- Ben-Ami, R.; Garcia-Effron, G.; Lewis, R.E.; Gamarra, S.; Leventakos, K.; Perlin, D.S.; Kontoyiannis, D.P. Fitness and Virulence Costs of Candida Albicans FKS1 Hot Spot Mutations Associated with Echinocandin Resistance. J. Infect. Dis. 2011, 204, 626–635. [Google Scholar] [CrossRef]
- Shields, R.K.; Kline, E.G.; Healey, K.R.; Kordalewska, M.; Perlin, D.S.; Hong Nguyen, M.; Clancy, C.J. Spontaneous Mutational Frequency and Fks Mutation Rates Vary by Echinocandin Agent against Candida Glabrata. Antimicrob. Agents Chemother. 2019, 63, e01692-18. [Google Scholar] [CrossRef]
- Katiyar, S.K.; Alastruey-Izquierdo, A.; Healey, K.R.; Johnson, M.E.; Perlin, D.S.; Edlind, T.D. Fks1 and Fks2 Are Functionally Redundant but Differentially Regulated in Candida Glabrata: Implications for Echinocandin Resistance. Antimicrob. Agents Chemother. 2012, 56, 6304–6309. [Google Scholar] [CrossRef]
- Katiyar, S.; Pfaller, M.; Edlind, T. Candida Albicans and Candida Glabrata Clinical Isolates Exhibiting Reduced Echinocandin Susceptibility. Antimicrob. Agents Chemother. 2006, 50, 2892–2894. [Google Scholar] [CrossRef] [PubMed]
- Pham, C.D.; Bolden, C.B.; Kuykendall, R.J.; Lockhart, S.R. Development of a Luminex-Based Multiplex Assay for Detection of Mutations Conferring Resistance to Echinocandins in Candida Glabrata. J. Clin. Microbiol. 2014, 52, 790–795. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Diekema, D.J.; Jones, R.N.; Castanheira, M. Use of Anidulafungin as a Surrogate Marker to Predict Susceptibility and Resistance to Caspofungin among 4290 Clinical Isolates of Candida by Using CLSI Methods and Interpretive Criteria. J. Clin. Microbiol. 2014, 52, 3223–3229. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Messer, S.A.; Diekema, D.J.; Jones, R.N.; Castanheira, M. Use of Micafungin as a Surrogate Marker to Predict Susceptibility and Resistance to Caspofungin among 3764 Clinical Isolates of Candida by Use of Clsi Methods and Interpretive Criteria. J. Clin. Microbiol. 2014, 52, 108–114. [Google Scholar] [CrossRef]
- Vu, B.G.; Stamnes, M.A.; Li, Y.; David Rogers, P.; Scott Moye-Rowley, W. The Candida Glabrata Upc2A Transcription Factor Is a Global Regulator of Antifungal Drug Resistance Pathways. PLoS Genet. 2021, 17, e1009582. [Google Scholar] [CrossRef]
- Pardini, G.; De Groot, P.W.J.; Coste, A.T.; Karababa, M.; Klis, F.M.; De Koster, C.G.; Sanglard, D. The CRH Family Coding for Cell Wall Glycosylphosphatidylinositol Proteins with a Predicted Transglycosidase Domain Affects Cell Wall Organization and Virulence of Candida Albicans. J. Biol. Chem. 2006, 281, 40399–40411. [Google Scholar] [CrossRef] [PubMed]
- Fonzi, W.A. PHR1 and PHR2 of Candida Albicans Encode Putative Glycosidases Required for Proper Cross-Linking of β-1,3- and β-1,6-Glucans. J. Bacteriol. 1999, 181, 7070–7079. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.W.; Quinn, P.J. Solvation Effects of Dimethyl Sulphoxide on the Structure of Phospholipid Bilayers. Biophys. Chem. 1998, 70, 35–39. [Google Scholar] [CrossRef]
- De Cesare, G.B.; Hafez, A.; Stead, D.; Llorens, C.; Munro, C.A. Biomarkers of Caspofungin Resistance in Candida Albicans Isolates: A Proteomic Approach. Virulence 2022, 13, 1005–1018. [Google Scholar] [CrossRef] [PubMed]
- Caplan, T.; Polvi, E.J.; Xie, J.L.; Buckhalter, S.; Leach, M.D.; Robbins, N.; Cowen, L.E. Functional Genomic Screening Reveals Core Modulators of Echinocandin Stress Responses in Candida Albicans. Cell Rep. 2018, 23, 2292–2298. [Google Scholar] [CrossRef]
- Kelly, J.; Kavanagh, K. Proteomic Analysis of Proteins Released from Growth-Arrested Candida Albicans Following Exposure to Caspofungin. Med. Mycol. 2010, 48, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Lafayette, S.L.; Collins, C.; Zaas, A.K.; Schell, W.A.; Betancourt-Quiroz, M.; Leslie Gunatilaka, A.A.; Perfect, J.R.; Cowen, L.E. PKC Signaling Regulates Drug Resistance of the Fungal Pathogen Candida Albicans via Circuitry Comprised of Mkc1, Calcineurin, and Hsp90. PLoS Pathog. 2010, 6, e1001069. [Google Scholar] [CrossRef] [PubMed]
- Cowen, L.E.; Steinbach, W.J. Stress, Drugs, and Evolution: The Role of Cellular Signaling in Fungal Drug Resistance. Eukaryot. Cell 2008, 7, 747–764. [Google Scholar] [CrossRef]
- Chamilos, G.; Lewis, R.E.; Albert, N.; Kontoyiannis, D.P. Paradoxical Effect of Echinocandins across Candida Species in Vitro: Evidence for Echinocandin-Specific and Candida Species-Related Differences. Antimicrob. Agents Chemother. 2007, 51, 2257–2259. [Google Scholar] [CrossRef]
- Munro, C.A.; Selvaggini, S.; De Bruijn, I.; Walker, L.; Lenardon, M.D.; Gerssen, B.; Milne, S.; Brown, A.J.P.; Gow, N.A.R. The PKC, HOG and Ca2+ Signalling Pathways Co-Ordinately Regulate Chitin Synthesis in Candida Albicans. Mol. Microbiol. 2007, 63, 1399–1413. [Google Scholar] [CrossRef]
- Plaine, A.; Walker, L.; Da Costa, G.; Mora-Montes, H.M.; McKinnon, A.; Gow, N.A.R.; Gaillardin, C.; Munro, C.A.; Richard, M.L. Functional Analysis of Candida Albicans GPI-Anchored Proteins: Roles in Cell Wall Integrity and Caspofungin Sensitivity. Fungal Genet. Biol. 2008, 45, 1404–1414. [Google Scholar] [CrossRef]
- Erbs, P.; Exinger, F.; Jund, R. Characterization of the Saccaromyces Cerevisiae FCY1 Gene Encoding Cytosine Deaminase and Its Homologue FCA1 of Candida Albicans. Curr. Genet. 1997, 31, 1–6. [Google Scholar] [CrossRef]
- McManus, B.A.; Moran, G.P.; Higgins, J.A.; Sullivan, D.J.; Coleman, D.C. A Ser29Leu Substitution in the Cytosine Deaminase Fca1p Is Responsible for Clade-Specific Flucytosine Resistance in Candida Dubliniensis. Antimicrob. Agents Chemother. 2009, 53, 4678–4685. [Google Scholar] [CrossRef]
- Lestrade, P.P.; Bentvelsen, R.G.; Schauwvlieghe, A.F.A.D.; Schalekamp, S.; Van Der Velden, W.J.F.M.; Kuiper, E.J.; Van Paassen, J.; Van Der Hoven, B.; Van Der Lee, H.A.; Melchers, W.J.G.; et al. Voriconazole Resistance and Mortality in Invasive Aspergillosis: A Multicenter Retrospective Cohort Study. Clin. Infect. Dis. 2019, 68, 1463–1471. [Google Scholar] [CrossRef] [PubMed]
- Jund, R.; Lacroute, F. Genetic and Physiological Aspects of Resistance to 5-Fluoropyrimidines in Saccharomyces Cerevisiae. J. Bacteriol. 1970, 102, 607–615. [Google Scholar] [CrossRef]
- Chevallier, M.R.; Jund, R.; Lacroute, F. Characterization of Cytosine Permeation in Saccharomyces Cerevisiae. J. Bacteriol. 1975, 122, 629–641. [Google Scholar] [CrossRef]
- Kern, L.; de Montigny, J.; Lacroute, F.; Jund, R. Regulation of the Pyrimidine Salvage Pathway by the FUR1 Gene Product of Saccharomyces Cerevisiae. Curr. Genet. 1991, 19, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Chapeland-Leclerc, F.; Hennequin, C.; Papon, N.; Noël, T.; Girard, A.; Socié, G.; Ribaud, P.; Lacroix, C. Acquisition of Flucytosine, Azole, and Caspofungin Resistance in Candida Glabrata Bloodstream Isolates Serially Obtained from a Hematopoietic Stem Cell Transplant Recipient. Antimicrob. Agents Chemother. 2010, 54, 1360–1362. [Google Scholar] [CrossRef] [PubMed]
- Dodgson, A.R.; Dodgson, K.J.; Pujol, C.; Pfaller, M.A.; Soll, D.R. Clade-Specific Flucytosine Resistance Is Due to a Single Nucleotide Change in the FUR1 Gene of Candida Albicans. Antimicrob. Agents Chemother. 2004, 48, 2223–2227. [Google Scholar] [CrossRef]
- Hope, W.W.; Tabernero, L.; Denning, D.W.; Anderson, M.J. Molecular Mechanisms of Primary Resistance to Flucytosine in Candida Albicans. Antimicrob. Agents Chemother. 2004, 48, 4377–4386. [Google Scholar] [CrossRef]
- Florent, M.; Noël, T.; Ruprich-Robert, G.; Da Silva, B.; Fitton-Ouhabi, V.; Chastin, C.; Papon, N.; Chapeland-Leclerc, F. Nonsense and Missense Mutations in FCY2 and FCY1 Genes Are Responsible for Flucytosine Resistance and Flucytosine-Fluconazole Cross-Resistance in Clinical Isolates of Candida Lusitaniae. Antimicrob. Agents Chemother. 2009, 53, 2982–2990. [Google Scholar] [CrossRef]
- Kannan, A.; Asner, S.A.; Trachsel, E.; Kelly, S.; Parker, J.; Sanglard, D. Comparative Genomics for the Elucidation of Multidrug Resistance in Candida Lusitaniae. mBio 2019, 10, e02512-19. [Google Scholar] [CrossRef]
- Chen, Y.N.; Lo, H.J.; Wu, C.C.; Ko, H.C.; Chang, T.P.; Yang, Y.L. Loss of Heterozygosity of FCY2 Leading to the Development of Flucytosine Resistance in Candida Tropicalis. Antimicrob. Agents Chemother. 2011, 55, 2506–2514. [Google Scholar] [CrossRef]
- Edlind, T.D.; Katiyar, S.K. Mutational Analysis of Flucytosine Resistance in Candida Glabrata. Antimicrob. Agents Chemother. 2010, 54, 4733–4738. [Google Scholar] [CrossRef]
- Vandeputte, P.; Pineau, L.; Larcher, G.; Noel, T.; Brèthes, D.; Chabasse, D.; Bouchara, J.P. Molecular Mechanisms of Resistance to 5-Fluorocytosine in Laboratory Mutants of Candida Glabrata. Mycopathologia 2011, 171, 11–21. [Google Scholar] [CrossRef]
- Ramage, G.; Mowat, E.; Jones, B.; Williams, C.; Lopez-Ribot, J. Our Current Understanding of Fungal Biofilms Fungal Biofilms Gordon Ramage et Al. Crit. Rev. Microbiol. 2009, 35, 340–355. [Google Scholar] [CrossRef]
- Wu, S.; Wang, Y.; Liu, N.; Dong, G.; Sheng, C. Tackling Fungal Resistance by Biofilm Inhibitors. J. Med. Chem. 2017, 60, 2193–2211. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Thompson, A.; Sobue, T.; Kashleva, H.; Xu, H.; Vasilakos, J.; Dongari-Bagtzoglou, A. Candida Albicans Biofilms Do Not Trigger Reactive Oxygen Species and Evade Neutrophil Killing. J. Infect. Dis. 2012, 206, 1936–1945. [Google Scholar] [CrossRef] [PubMed]
- Atriwal, T.; Azeem, K.; Husain, F.M.; Hussain, A.; Khan, M.N.; Alajmi, M.F.; Abid, M. Mechanistic Understanding of Candida Albicans Biofilm Formation and Approaches for Its Inhibition. Front. Microbiol. 2021, 12, 638609. [Google Scholar] [CrossRef] [PubMed]
- Nett, J.E.; Andes, D.R. Contributions of the Biofilm Matrix to Candida Pathogenesis. J. Fungi 2020, 6, 21. [Google Scholar] [CrossRef]
- Fan, F.M.; Liu, Y.; Liu, Y.Q.; Lv, R.X.; Sun, W.; Ding, W.J.; Cai, Y.X.; Li, W.W.; Liu, X.; Qu, W. Candida Albicans Biofilms: Antifungal Resistance, Immune Evasion, and Emerging Therapeutic Strategies. Int. J. Antimicrob. Agents 2022, 60, 106673. [Google Scholar] [CrossRef] [PubMed]
- Chandra, J.; Kuhn, D.M.; Mukherjee, P.K.; Hoyer, L.L.; McCormick, T.; Ghannoum, M.A. Biofilm Formation by the Fungal Pathogen Candida Albicans: Development, Architecture, and Drug Resistance. J. Bacteriol. 2001, 183, 5385–5394. [Google Scholar] [CrossRef]
- de Beer, D.; Stoodley, P.; Lewandowski, Z. Liquid Flow in Heterogeneous Biofilms. Biotechnol. Bioeng. 1994, 44, 636–641. [Google Scholar] [CrossRef]
- Lawrence, J.R.; Korber, D.R.; Hoyle, B.D.; Costerton, J.W.; Caldwell, D.E. Optical Sectioning of Microbial Biofilms. J. Bacteriol. 1991, 173, 6558–6567. [Google Scholar] [CrossRef]
- Tumbarello, M.; Posteraro, B.; Trecarichi, E.M.; Fiori, B.; Rossi, M.; Porta, R.; Donati, K.D.G.; La Sorda, M.; Spanu, T.; Fadda, G.; et al. Biofilm Production by Candida Species and Inadequate Antifungal Therapy as Predictors of Mortality for Patients with Candidemia. J. Clin. Microbiol. 2007, 45, 1843–1850. [Google Scholar] [CrossRef]
- Shin, J.H.; Kee, S.J.; Shin, M.G.; Kim, S.H.; Shin, D.H.; Lee, S.K.; Suh, S.P.; Ryang, D.W. Biofilm Production by Isolates of Candida Species Recovered from Nonneutropenic Patients: Comparison of Bloodstream Isolates with Isolates from Other Sources. J. Clin. Microbiol. 2002, 40, 1244–1248. [Google Scholar] [CrossRef] [PubMed]
- Ramage, G.; Martínez, J.P.; López-Ribot, J.L. Candida Biofilms on Implanted Biomaterials: A Clinically Significant Problem. FEMS Yeast Res. 2006, 6, 979–986. [Google Scholar] [CrossRef]
- Rajendran, R.; Sherry, L.; Nile, C.J.; Sherriff, A.; Johnson, E.M.; Hanson, M.F.; Williams, C.; Munro, C.A.; Jones, B.J.; Ramage, G. Biofilm Formation Is a Risk Factor for Mortality in Patients with Candida Albicans Bloodstream Infection-Scotland, 2012–2013. Clin. Microbiol. Infect. 2016, 22, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Horton, M.V.; Nett, J.E. Candida Auris Infection and Biofilm Formation: Going Beyond the Surface. Curr. Clin. Microbiol. Rep. 2020, 7, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Ramage, G.; Vande Walle, K.; Wickes, B.L.; López-Ribot, J.L. Standardized Method for in Vitro Antifungal Susceptibility Testing of Candida Albicans Biofilms. Antimicrob. Agents Chemother. 2001, 45, 2475–2479. [Google Scholar] [CrossRef]
- Knot, P.D.; Suci, P.A.; Miller, R.L.; Nelson, R.D.; Tyler, B.J. A Small Subpopulation of Blastospores in Candida Albicans Biofilms Exhibit Resistance to Amphotericin B Associated with Differential Regulation of Ergosterol and β-1,6-Glucan Pathway Genes. Antimicrob. Agents Chemother. 2006, 50, 3708–3716. [Google Scholar] [CrossRef]
- Ramage, G.; Rajendran, R.; Sherry, L.; Williams, C. Fungal Biofilm Resistance. Int. J. Microbiol. 2012, 2012, 1–14. [Google Scholar] [CrossRef]
- Perumal, P.; Mekala, S.; Chaffin, W.L.J. Role for Cell Density in Antifungal Drug Resistance in Candida Albicans Biofilms. Antimicrob. Agents Chemother. 2007, 51, 2454–2463. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, T.B.; Givskov, M. Quorum Sensing Inhibitors: A Bargain of Effects. Microbiology 2006, 152, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Deng, K.; Jiang, W.; Jiang, Y.; Deng, Q.; Cao, J.; Yang, W.; Zhao, X. ALS3 Expression as an Indicator for Candida Albicans Biofilm Formation and Drug Resistance. Front. Microbiol. 2021, 12, 655242. [Google Scholar] [CrossRef] [PubMed]
- Nett, J.; Lincoln, L.; Marchillo, K.; Massey, R.; Holoyda, K.; Hoff, B.; VanHandel, M.; Andes, D. Putative Role of β-1,3 Glucans in Candida Albicans Biofilm Resistance. Antimicrob. Agents Chemother. 2007, 51, 510–520. [Google Scholar] [CrossRef]
- Nett, J.E.; Sanchez, H.; Cain, M.T.; Andes, D.R. Genetic Basis of Candida Biofilm Resistance Due to Drug-Sequestering Matrix Glucan. J. Infect. Dis. 2010, 202, 171–175. [Google Scholar] [CrossRef]
- Tan, Y.; Ma, S.; Leonhard, M.; Moser, D.; Schneider-Stickler, B. β-1,3-Glucanase Disrupts Biofilm Formation and Increases Antifungal Susceptibility of Candida Albicans DAY185. Int. J. Biol. Macromol. 2018, 108, 942–946. [Google Scholar] [CrossRef]
- Al-Fattani, M.A.; Douglas, L.J. Biofilm Matrix of Candida Albicans and Candida Tropicalis: Chemical Composition and Role in Drug Resistance. J. Med. Microbiol. 2006, 55, 999–1008. [Google Scholar] [CrossRef]
- Nett, J.E.; Crawford, K.; Marchillo, K.; Andes, D.R. Role of Fks1p and Matrix Glucan in Candida Albicans Biofilm Resistance to an Echinocandin, Pyrimidine, and Polyene. Antimicrob. Agents Chemother. 2010, 54, 3505–3508. [Google Scholar] [CrossRef]
- Kaur, J.; Nobile, C.J. Antifungal Drug-Resistance Mechanisms in Candida Biofilms. Curr. Opin. Microbiol. 2023, 71, 102237. [Google Scholar] [CrossRef]
- Taff, H.T.; Nett, J.E.; Zarnowski, R.; Ross, K.M.; Sanchez, H.; Cain, M.T.; Hamaker, J.; Mitchell, A.P.; Andes, D.R. A Candida Biofilm-Induced Pathway for Matrix Glucan Delivery: Implications for Drug Resistance. PLoS Pathog. 2012, 8, e1002848. [Google Scholar] [CrossRef]
- Nailis, H.; Vandenbosch, D.; Deforce, D.; Nelis, H.J.; Coenye, T. Transcriptional Response to Fluconazole and Amphotericin B in Candida Albicans Biofilms. Res. Microbiol. 2010, 161, 284–292. [Google Scholar] [CrossRef]
- Borecká-Melkusová, S.; Moran, G.P.; Sullivan, D.J.; Kucharíková, S.; Chorvát, D.; Bujdáková, H. The Expression of Genes Involved in the Ergosterol Biosynthesis Pathway in Candida Albicans and Candida Dubliniensis Biofilms Exposed to Fluconazole. Mycoses 2009, 52, 118–128. [Google Scholar] [CrossRef]
- Katragkou, A.; Chatzimoschou, A.; Simitsopoulou, M.; Dalakiouridou, M.; Diza-Mataftsi, E.; Tsantali, C.; Roilides, E. Differential Activities of Newer Antifungal Agents against Candida Albicans and Candida Parapsilosis Biofilms. Antimicrob. Agents Chemother. 2008, 52, 357–360. [Google Scholar] [CrossRef]
- Rossignol, T.; Ding, C.; Guida, A.; D’Enfert, C.; Higgins, D.G.; Butler, G. Correlation between Biofilm Formation and the Hypoxic Response in Candida Parapsilosis. Eukaryot. Cell 2009, 8, 550–559. [Google Scholar] [CrossRef]
- Yu, L.H.; Wei, X.; Ma, M.; Chen, X.J.; Xu, S.B. Possible Inhibitory Molecular Mechanism of Farnesol on the Development of Fluconazole Resistance in Candida Albicans Biofilm. Antimicrob. Agents Chemother. 2012, 56, 770–775. [Google Scholar] [CrossRef] [PubMed]
- Lepak, A.; Nett, J.; Lincoln, L.; Marchillo, K.; Andes, D. Time Course of Microbiologic Outcome and Gene Expression in Candida Albicans during and Following in Vitro and in Vivo Exposure to Fluconazole. Antimicrob. Agents Chemother. 2006, 50, 1311–1319. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.K.; Chandra, J.; Kuhn, D.M.; Ghannoum, M.A. Mechanism of Fluconazole Resistance in Candida Albicans Biofilms: Phase-Specific Role of Efflux Pumps and Membrane Sterols. Infect. Immun. 2003, 71, 4333–4340. [Google Scholar] [CrossRef]
- Ramage, G.; Bachmann, S.; Patterson, T.F.; Wickes, B.L.; López-Ribot, J.L. Investigation of Multidrug Efflux Pumps in Relation to Fluconazole Resistance in Candida Albicans Biofilms. J. Antimicrob. Chemother. 2002, 49, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Mateus, C.; Crow, S.A.; Ahearn, D.G. Adherence of Candida Albicans to Silicone Induces Immediate Enhanced Tolerance to Fluconazole. Antimicrob. Agents Chemother. 2004, 48, 3358–3366. [Google Scholar] [CrossRef]
- Bizerra, F.C.; Nakamura, C.V.; De Poersch, C.; Estivalet Svidzinski, T.I.; Borsato Quesada, R.M.; Goldenberg, S.; Krieger, M.A.; Yamada-Ogatta, S.F. Characteristics of Biofilm Formation by Candida Tropicalis and Antifungal Resistance. FEMS Yeast Res. 2008, 8, 442–450. [Google Scholar] [CrossRef]
- Song, J.W.; Shin, J.H.; Kee, S.J.; Kim, S.H.; Shin, M.G.; Suh, S.P.; Ryang, D.W. Expression of CgCDR1, CgCDR2, and CgERG11 in Candida Glabrata Biofilms Formed by Bloodstream Isolates. Med. Mycol. 2009, 47, 545–548. [Google Scholar] [CrossRef]
- Andes, D.; Nett, J.; Oschel, P.; Albrecht, R.; Marchillo, K.; Pitula, A. Development and Characterization of an in Vivo Central Venous Catheter Candida Albicans Biofilm Model. Infect. Immun. 2004, 72, 6023–6031. [Google Scholar] [CrossRef]
- Kuhn, D.M.; George, T.; Chandra, J.; Mukherjee, P.K.; Ghannoum, M.A. Antifungal Susceptibility of Candida Biofilms: Unique Efficacy of Amphotericin B Lipid Formulations and Echinocandins. Antimicrob. Agents Chemother. 2002, 46, 1773–1780. [Google Scholar] [CrossRef] [PubMed]
- Toulet, D.; Debarre, C.; Imbert, C. Could Liposomal Amphotericin B (L-AMB) Lock Solutions Be Useful to Inhibit Candida Spp. Biofilms on Silicone Biomaterials? J. Antimicrob. Chemother. 2012, 67, 430–432. [Google Scholar] [CrossRef]
- Moen, M.D.; Lyseng-Williamson, K.A.; Scott, L.J. Liposomal Amphotericin B: A Review of Its Use as Empirical Therapy in Febrile Neutropenia and in the Treatment of Invasive Fungal Infections. Drugs 2009, 69, 361–392. [Google Scholar] [CrossRef] [PubMed]
- Gursu, B.Y.; Dag, İ.; Dikmen, G. Antifungal and Antibiofilm Efficacy of Cinnamaldehyde-Loaded Poly(DL-Lactide-Co-Glycolide) (PLGA) Nanoparticles against Candida Albicans. Int. Microbiol. 2022, 25, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Shariati, A.; Didehdar, M.; Razavi, S.; Heidary, M.; Soroush, F.; Chegini, Z. Natural Compounds: A Hopeful Promise as an Antibiofilm Agent Against Candida Species. Front. Pharmacol. 2022, 13, 917787. [Google Scholar] [CrossRef] [PubMed]
- Hadley, S.; Martinez, J.A.; McDermott, L.; Rapino, B.; Snydman, D.R. Real-Time Antifungal Susceptibility Screening Aids Management of Invasive Yeasts Infections in Immunocompromised Patients. J. Antimicrob. Chemother. 2002, 49, 415–419. [Google Scholar] [CrossRef]
- Xu, Z.; Zhang, L.X.; Zhang, J.D.; Cao, Y.B.; Yu, Y.Y.; Wang, D.J.; Ying, K.; Chen, W.S.; Jiang, Y.Y. CDNA Microarray Analysis of Differential Gene Expression and Regulation in Clinically Drug-Resistant Isolates of Candida Albicans from Bone Marrow Transplanted Patients. Int. J. Med. Microbiol. 2006, 296, 421–434. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, R.P.; Thakku, S.G.; Hung, D.T. Harnessing CRISPR Effectors for Infectious Disease Diagnostics. ACS Infect. Dis. 2018, 4, 1278–1282. [Google Scholar] [CrossRef]
- Matthijs, G.; Souche, E.; Alders, M.; Corveleyn, A.; Eck, S.; Feenstra, I.; Race, V.; Sistermans, E.; Sturm, M.; Weiss, M.; et al. Guidelines for Diagnostic Next-Generation Sequencing. Eur. J. Hum. Genet. 2016, 24, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Scheler, O.; Glynn, B.; Kurg, A. Nucleic Acid Detection Technologies and Marker Molecules in Bacterial Diagnostics. Expert. Rev. Mol. Diagn. 2014, 14, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chen, F.; Li, Q.; Wang, L.; Fan, C. Isothermal Amplification of Nucleic Acids. Chem. Rev. 2015, 115, 12491–12545. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Antifungal Class | Drug Name | Fungal Species | CLSI MIC Breakpoints (µg/mL) | EUCAST MIC Breakpoints (µg/mL) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| S | I | SSD | R | S | I | SSD | R | |||
| Azole | Fluconazole (FLZ) | C. albicans | ≤2 | - | 4 | ≥8 | ≤2 | 4 | - | >4 |
| C. dubliniensis | - | - | - | - | ≤2 | 4 | - | >4 | ||
| N. glabrata | - | - | ≤32 | ≥64 | ≤0.001 | ≤16 | - | >16 | ||
| P. kudriavzevii | - | - | - | - | - | - | - | - | ||
| C. parapsilosis | ≤2 | 4 | ≥8 | ≤2 | 4 | - | >4 | |||
| C. tropicalis | ≤2 | 4 | ≥8 | ≤2 | 4 | - | >4 | |||
| Voriconazole (VOR) | C. albicans | ≤0.12 | 0.25–0.5 | - | ≥1 | ≤0.06 | 0.125–0.25 | - | >0.25 | |
| C. dubliniensis | ≤0.06 | 0.125–0.25 | - | >0.25 | ||||||
| N. glabrata | - | - | - | - | - | - | - | - | ||
| P. kudriavzevii | ≤0.5 | 1 | - | ≥2 | - | - | - | - | ||
| C. parapsilosis | ≤0.12 | 0.25–0.5 | - | ≥1 | ≤0.125 | 0.25 | - | >0.25 | ||
| C. tropicalis | ≤0.12 | 0.25–0.5 | - | ≥1 | ≤0.125 | 0.25 | - | >0.25 | ||
| Posaconazole | C. albicans | - | - | - | - | ≤0.06 | - | - | >0.06 | |
| C. dubliniensis | - | - | - | - | ≤0.06 | - | - | >0.06 | ||
| C. parapsilosis | - | - | - | - | ≤0.06 | - | - | >0.06 | ||
| C. tropicalis | - | - | - | - | ≤0.06 | - | - | >0.06 | ||
| Itraconazole | C. albicans | - | - | - | - | ≤0.06 | - | - | >0.06 | |
| C. dubliniensis | - | - | - | - | ≤0.06 | - | - | >0.06 | ||
| C. parapsilosis | - | - | - | - | ≤0.125 | - | - | >0.125 | ||
| C. tropicalis | - | - | - | - | ≤0.125 | - | - | >0.125 | ||
| Echinocandin | Caspofungin | C. albicans | ≤0.25 | 0.5 | - | ≥1 | - | - | - | - |
| N. glabrata | ≤0.12 | 0.25 | - | ≥0.5 | - | - | - | - | ||
| M. guilliermondii | ≤2 | 4 | - | ≥8 | - | - | - | - | ||
| P. kudriavzevii | ≤0.25 | 0.5 | - | ≥1 | - | - | - | - | ||
| C. parapsilosis | ≤2 | 4 | - | ≥8 | - | - | - | - | ||
| C. tropicalis | ≤0.25 | 0.5 | - | ≥1 | - | - | - | - | ||
| Anidulafungin | C. albicans | ≤0.25 | 0.5 | - | ≥1 | ≤0.03 | - | - | >0.03 | |
| N. glabrata | ≤0.12 | 0.25 | ≥0.5 | ≤0.06 | - | - | >0.06 | |||
| M. guilliermondii | ≤2 | 4 | ≥8 | - | - | - | - | |||
| P. kudriavzevii | ≤0.25 | 0.5 | ≥1 | ≤0.06 | - | - | >0.06 | |||
| C. parapsilosis | ≤2 | 4 | ≥8 | ≤4 | - | - | >4 | |||
| C. tropicalis | ≤0.25 | 0.5 | ≥1 | ≤0.06 | - | - | >0.06 | |||
| Micafungin | C. albicans | ≤0.25 | 0.5 | - | ≥1 | ≤0.016 | - | - | >0.016 | |
| N. glabrata | ≤0.06 | 0.12 | - | ≥0.25 | ≤0.03 | - | - | >0.03 | ||
| M. guilliermondii | ≤2 | 4 | - | ≥8 | - | - | - | - | ||
| P. kudriavzevii | ≤0.25 | 0.5 | - | ≥1 | - | - | - | - | ||
| C. parapsilosis | ≤2 | 4 | - | ≥8 | ≤2 | - | - | >2 | ||
| C. tropicalis | ≤0.25 | 0.5 | - | ≥1 | - | - | - | - | ||
| Polyene | Amphotericin B | C. albicans | ≤1 | - | - | >1 | ||||
| C. dubliniensis | ≤1 | - | - | >1 | ||||||
| N. glabrata | ≤1 | - | - | >1 | ||||||
| P. kudriavzevii | ≤1 | - | - | >1 | ||||||
| C. parapsilosis | ≤1 | - | - | >1 | ||||||
| C.tropicalis | ≤1 | - | - | >1 | ||||||
| C. auris | Tentative breakpoints based on a mouse model reported by the CDC (2020): S (≤1), R (≥2) | |||||||||
| Nystatin | Candida | CLSI and EUCAST MIC breakpoints unavailable.Broth microdilution estimates based on Brito et al., 2011: S (≤4), I (8–32), R (≥64) | ||||||||
| Gene | Candida Species | Mutation | Type of Mutation | Antifungal Resistance | Location | Isolate Type | Ref. |
|---|---|---|---|---|---|---|---|
| ERG11 (lanosterol 14a-demethylase) | C. albicans | Hotspot regions: aa105–165, 266–287 and 405–488 | Substitution | Azole | USA | Clinical | [36] |
| A61V, S405F, G448E, F449S, G464S, R467K and I471T | Non-synonymous substitution | Fluconazole | China | Clinical | [86] | ||
| Y132H, Y132F, K143R and K143Q | Non-synonymous substitution | Fluconazole and voriconazole | China | Clinical | [86,89] | ||
| A114S and Y257H | Non-synonymous substitution | Fluconazole and voriconazole | China | Clinical | [86,90] | ||
| T315A, Y118A, Y18F and Y118T | Non-synonymous substitution | Fluconazole | - | Lab-created | [91,92] | ||
| K128T | Non-synonymous substitution | Likely no effect | China | Clinical | [86,93] | ||
| D116E and E266D | Non-synonymous substitution | No effect on protein function or resistance | USA | Clinical | [36] | ||
| C. auris | F126T, Y132F and K143R | Non-synonymous substitution | Fluconazole | South Africa, Venezuela, India | Clinical | [28] | |
| N. glabrata | C108G, C423T and A1581G | Synonymous substitution | No effect | Brazil | Clinical | [94] | |
| T768C, A1023G and T1557A | Synonymous substitution | No effect | Slovakia | Clinical | [95] | ||
| E502V | Non-synonymous substitution | No effect | Slovakia | Clinical | [96] | ||
| P. kudriavzevii | G524R | Non-synonymous substitution | No effect on protein function or resistance | Brazil | Clinical | [94] | |
| Y166S | Non-synonymous substitution | Voriconazole | Brazil | Clinical | [94] | ||
| C. tropicalis | Y132F | Missense | Fluconazole | Brazil | Clinical | [97] | |
| K143R | Non-synonymous substitution | Fluconazole, voriconazole and itraconazole | Brazil | Clinical | [98] | ||
| ERG3 (C5 sterol desaturase) | N. glabrata | Q139A | Non-synonymous substitution | Fluconazole | Korea | Clinical | [99] |
| UPC2 (TF, regulates most ERG genes) | C. albicans | G648D, G648S, A643T, Y642F, A646V and W478C | GOF substitution | Fluconazole | USA | Clinical | [100] |
| A643V | GOF substitution | Fluconazole | USA | Clinical | [100] | ||
| G307S and G448E | GOF substitution | Fluconazole | Germany | Clinical | [101] |
| Gene | Candida Species | Mutation | Type of Mutation | Antifungal Resistance | Location | Isolate Type | Ref. |
|---|---|---|---|---|---|---|---|
| CDR1 + CDR2 (ABC-Ts) | C. albicans | Chr 3 trisomy | Increased cdr1 and cdr2 copy numbers | Azole | - | In vitro | [124] |
| MLT1 (ABC-T) | C. albicans | K710A | Loss of function | Reduced azole resistance | - | In vitro | [123] |
| F765Δ | Loss of function | Reduced azole resistance | - | In vitro | [123] | ||
| TAC1 (TF, regulates CDR1, CDR2 and PDR16) | C. albicans | T225A, V736A, N972D, N977D, G980E and G980W | GOF substitution | Azole | USA | Clinical | [125] |
| C. auris | K143R, F214S, R495G and A640V | Non-synonymous substitution | Fluconazole | USA | Clinical/ in vitro | [126] | |
| MRR1 (TF, regulates MDR1) | C. albicans | P683S and P683H | GOF substitution | Azole | Germany | Clinical | [101,127] |
| C. dubliniensis | T374I, S595Y and C866Y | GOF substitution | Azole | Ireland | Clinical | [128,129,130] | |
| T965∆ and (D987-I998)∆ | Deletion | Azole | Ireland | Clinical | [128] | ||
| PDR16 (phosphatidylinositol transfer protein) | N. glabrata | ∆pdr16 | Gene deletion | Reduced resistance to fluconazole, itraconazole and ketoconazole miconazole | - | In vitro | [131] |
| PDR1 (TF, regulates CDR1, SNQ2, PDH1 and QDR2) | N. glabrata | Hotspot regions: 312–382, 800–1107 and 539–632 | GOF substitution | Azole | Italy, Switzerland, France and Japan | Clinical | [35,132] |
| L328F, R376W, D1082G, T588A, T607S, E1083Q, Y584C, D876Y, L280F, N691D, S316I, D261G, R293I, R592S, G583S, S343F and R376G | GOF substitution | Fluconazole | Italy, Switzerland, France and Japan | Clinical | [132] |
| Gene | Candida Species | Mutation | Type of Mutation | Antifungal Resistance | Location | Isolate Type | Ref. |
|---|---|---|---|---|---|---|---|
| FKS1 (β1–3 glucan synthase) | C. albicans | Hotspot regions: aa 637–654 and 1345–1365 | Non-synonymous substitution | Echinocandin | - | Clinical | [74,162] |
| S645F | Non-synonymous substitution | Echinocandin | USA | Clinical | [170] | ||
| C. auris | F635Y, F635L, S639F and R1354S | Non-synonymous substitution | Echinocandin | India | In vitro/ in vivo | [168] | |
| N. glabrata | F625C and S629P | Non-synonymous substitution | Echinocandin | - | Clinical/ in vitro | [171,172] | |
| F625∆ | Deletion | Echinocandin | - | Clinical/ in vitro | [171,172] | ||
| P. kudriavzevii | F655C | Non-synonymous substitution | Echinocandin | USA | Clinical | [166] | |
| C. parapsilosis | P660A | Non-synonymous substitution | Echinocandin | - | All species members | [161] | |
| FKS2 (β1–3 glucan synthase) | N. glabrata | F659S and F659V | Non-synonymous substitution | Echinocandin | USA | Clinical | [158,159,173] |
| F659∆ | Deletion | Echinocandin | USA | Clinical | [158,159,173] | ||
| S663P and S663F | Non-synonymous substitution | Echinocandin | USA | Clinical | [171,172] | ||
| E655G, E655K, P667H and P667T | Non-synonymous substitution | Echinocandin | USA | Clinical | [171,172] | ||
| R1378S and R1378G | Non-synonymous substitution | Echinocandin | USA | Clinical | [171,172] |
| Gene | Candida Species | Mutation | Type of Mutation | Antifungal Resistance | Location | Isolate Type | Ref. |
|---|---|---|---|---|---|---|---|
| FCA1/FCY1 (cytosine deaminase) | C. albicans | G28D and S29L | LOF substitution | 5FC | UK | Clinical | [197] |
| C. dubliniensis | S29L | Non-synonymous substitution | 5FC | Egypt and Saudi Arabia | Clinical | [190] | |
| N. glabrata | A15D, G11D and W148R | Non-synonymous substitution | 5FC | - | In vitro | [201] | |
| FCY2 (cytosine permease) | C. albicans | A176G | LOF substitution | 5FC | UK | Clinical | [197] |
| C. tropicalis | G145T | Non-synonymous substitution | 5-FC | Taiwan | Clinical | [200] | |
| FUR1 (uracil phosphoribosyltransferase (UPRT)) | C. albicans | C101R | LOF substitution | 5FC | Multiple countries | Clinical | [196,197] |
| N. glabrata | G190D | LOF substitution | 5FC | France | Clinical | [195] | |
| I83K and D193G | LOF substitution | 5FC/5FU | - | In vitro | [201,202] | ||
| ∆G73-V81 | LOF Deletion | 5FC/5FU | - | In vitro | [201,202] | ||
| MSH2 (DNA mismatch repair) | N. glabrata | V239L | Non-synonymous substitution | Fluconazole or echinocandin | Multiple countries | Clinical | [35] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Czajka, K.M.; Venkataraman, K.; Brabant-Kirwan, D.; Santi, S.A.; Verschoor, C.; Appanna, V.D.; Singh, R.; Saunders, D.P.; Tharmalingam, S. Molecular Mechanisms Associated with Antifungal Resistance in Pathogenic Candida Species. Cells 2023, 12, 2655. https://doi.org/10.3390/cells12222655
Czajka KM, Venkataraman K, Brabant-Kirwan D, Santi SA, Verschoor C, Appanna VD, Singh R, Saunders DP, Tharmalingam S. Molecular Mechanisms Associated with Antifungal Resistance in Pathogenic Candida Species. Cells. 2023; 12(22):2655. https://doi.org/10.3390/cells12222655
Chicago/Turabian StyleCzajka, Karolina M., Krishnan Venkataraman, Danielle Brabant-Kirwan, Stacey A. Santi, Chris Verschoor, Vasu D. Appanna, Ravi Singh, Deborah P. Saunders, and Sujeenthar Tharmalingam. 2023. "Molecular Mechanisms Associated with Antifungal Resistance in Pathogenic Candida Species" Cells 12, no. 22: 2655. https://doi.org/10.3390/cells12222655
APA StyleCzajka, K. M., Venkataraman, K., Brabant-Kirwan, D., Santi, S. A., Verschoor, C., Appanna, V. D., Singh, R., Saunders, D. P., & Tharmalingam, S. (2023). Molecular Mechanisms Associated with Antifungal Resistance in Pathogenic Candida Species. Cells, 12(22), 2655. https://doi.org/10.3390/cells12222655