

Advances of Wnt Signalling Pathway in Colorectal Cancer
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
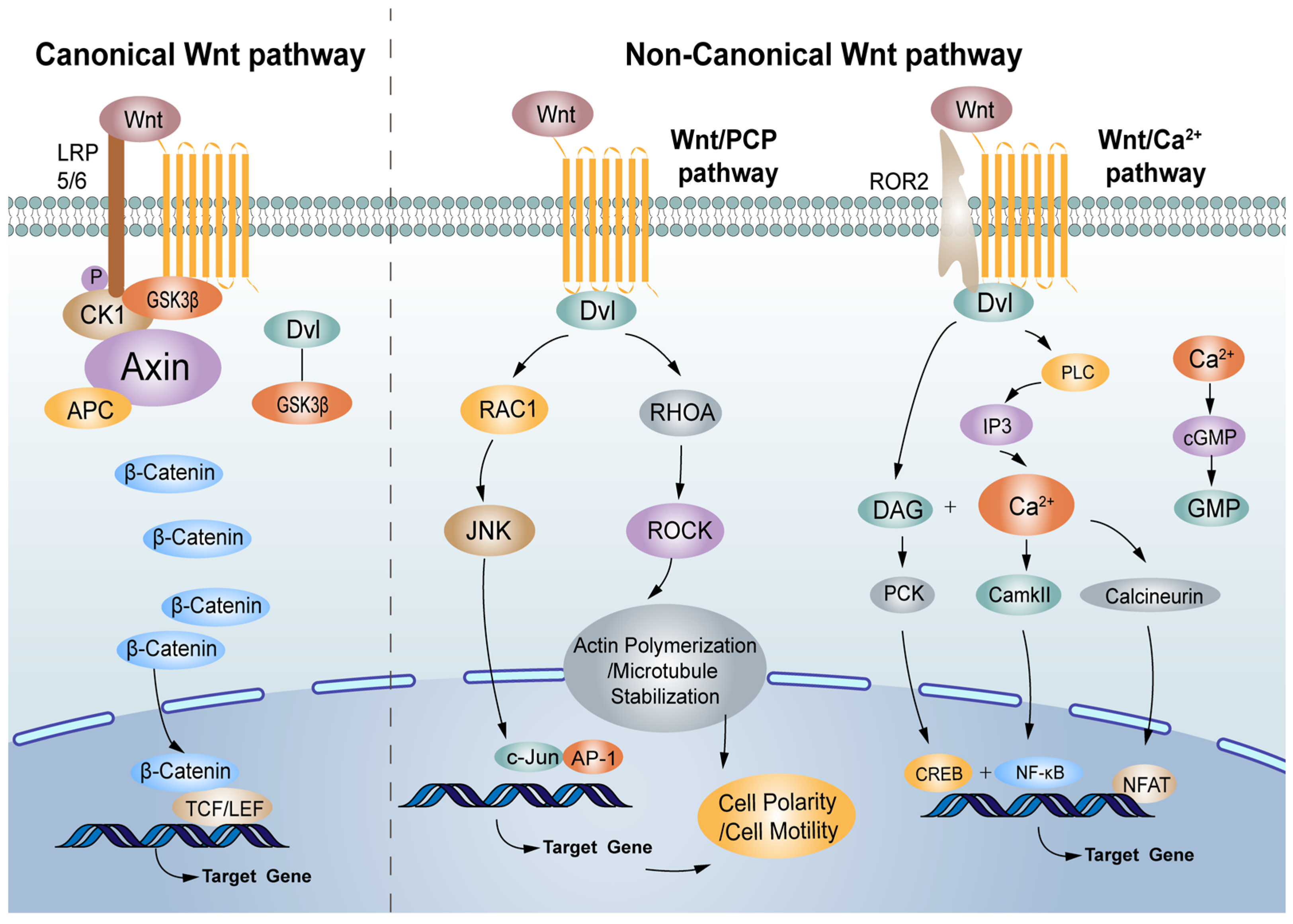
2. The Canonical Wnt/β-Catenin Pathway
3. The Non-Canonical Wnt Signalling Pathway
3.1. The Wnt/PCP Signalling Pathway
3.2. The Wnt/Ca2+ Signalling Pathway
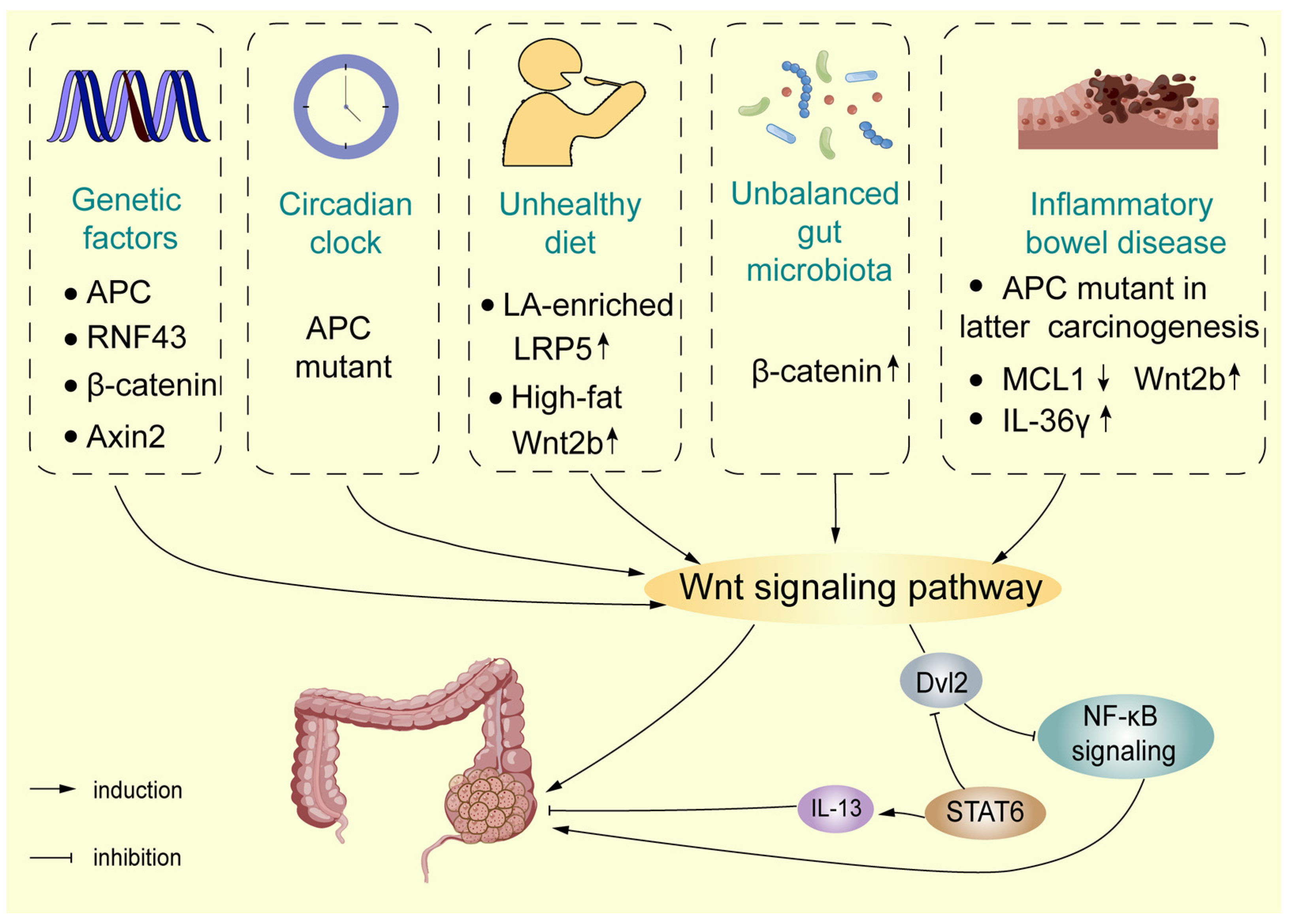
4. The Wnt Signalling Pathway in Occurrence of Colorectal Cancer (CRC)
4.1. The Wnt Signalling Pathway and Environmental Exposure
4.1.1. Lifestyle
4.1.2. Pro-Tumorigenic Bacteria
4.1.3. Gut Microbiota
4.2. The Wnt Signalling Pathway in Colitis-Associated Carcinoma
5. The Wnt Signalling Pathway in the Development of CRC
5.1. The Wnt Signalling Pathway in Cancer Stem Cells (CSCs)
5.2. The Wnt Signalling Pathway in the Tumour Microenvironment
5.3. The Wnt Signalling Pathway in Angiogenesis
6. The Wnt Signalling Pathway in the Invasion and Metastasis of CRC
7. The Wnt Signalling Pathway in CRC Prognosis
7.1. The Wnt Signalling Pathway in Chemotherapy Drug Resistance
7.2. The Wnt Signalling Pathway in Immunotherapy Drug Resistance
7.3. The Wnt Signalling Pathway as a Prognostic Indicator of CRC
8. Challenges and Prospects
9. Conclusions
Funding
Conflicts of Interest
References
- Steinhart, Z.; Angers, S. Wnt Signaling in Development and Tissue Homeostasis. Development 2018, 145, dev146589. [Google Scholar] [CrossRef]
- Taciak, B.; Pruszynska, I.; Kiraga, L.; Bialasek, M.; Krol, M. Wnt Signaling Pathway in Development and Cancer. J. Physiol. Pharmacol. 2018, 69, 185–195. [Google Scholar] [CrossRef]
- Nie, X.; Liu, H.; Liu, L.; Wang, Y.-D.; Chen, W.-D. Emerging Roles of Wnt Ligands in Human Colorectal Cancer. Front. Oncol. 2020, 10, 1341. [Google Scholar] [CrossRef] [PubMed]
- Parichha, A.; Suresh, V.; Chatterjee, M.; Kshirsagar, A.; Ben-Reuven, L.; Olender, T.; Taketo, M.M.; Radosevic, V.; Bobic-Rasonja, M.; Trnski, S.; et al. Constitutive Activation of Canonical Wnt Signaling Disrupts Choroid Plexus Epithelial Fate. Nat. Commun. 2022, 13, 633. [Google Scholar] [CrossRef] [PubMed]
- Morgan, E.; Arnold, M.; Gini, A.; Lorenzoni, V.; Cabasag, C.J.; Laversanne, M.; Vignat, J.; Ferlay, J.; Murphy, N.; Bray, F. Global Burden of Colorectal Cancer in 2020 and 2040: Incidence and Mortality Estimates from GLOBOCAN. Gut 2022, 72, 338–344. [Google Scholar] [CrossRef]
- Xi, Y.; Xu, P. Global Colorectal Cancer Burden in 2020 and Projections to 2040. Transl. Oncol. 2021, 14, 101174. [Google Scholar] [CrossRef]
- Ochs-Balcom, H.M.; Kanth, P.; Cannon-Albright, L.A. Early-Onset Colorectal Cancer Risk Extends to Second- and Third-Degree Relatives. Cancer Epidemiol. 2021, 73, 101973. [Google Scholar] [CrossRef]
- Gausman, V.; Dornblaser, D.; Anand, S.; Hayes, R.B.; O’Connell, K.; Du, M.; Liang, P.S. Risk Factors Associated with Early-Onset Colorectal Cancer. Clin. Gastroenterol. Hepatol. 2020, 18, 2752–2759. [Google Scholar] [CrossRef]
- Aran, V.; Victorino, A.P.; Thuler, L.C.; Ferreira, C.G. Colorectal Cancer: Epidemiology, Disease Mechanisms and Interventions to Reduce Onset and Mortality. Clin. Colorectal Cancer 2016, 15, 195–203. [Google Scholar] [CrossRef]
- Koni, M.; Pinnaro, V.; Brizzi, M.F. The Wnt Signalling Pathway: A Tailored Target in Cancer. Int. J. Mol. Sci. 2020, 21, 7697. [Google Scholar] [CrossRef]
- Clevers, H.; Nusse, R. Wnt/Beta-Catenin Signaling and Disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Xu, X.; Chen, D.; Zhao, F.; Wang, W. Therapeutic Potential of Targeting the Wnt/Beta-Catenin Signaling Pathway in Colorectal Cancer. Biomed. Pharmacother. 2019, 110, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Li, V.S.W.; Ng, S.S.; Boersema, P.J.; Low, T.Y.; Karthaus, W.R.; Gerlach, J.P.; Mohammed, S.; Heck, A.J.R.; Maurice, M.M.; Mahmoudi, T.; et al. Wnt Signaling through Inhibition of Beta-Catenin Degradation in an Intact Axin1 Complex. Cell 2012, 149, 1245–1256. [Google Scholar] [CrossRef]
- Mantilla, C.; Suárez Mellado, I.; Duque Jaramillo, A.; Navas, M.C. β-catenin signaling mechanisms and its role in carcinogenesis. CES Med. 2015, 29, 109–127. [Google Scholar]
- Kim, N.-G.; Xu, C.; Gumbiner, B.M. Identification of Targets of the Wnt Pathway Destruction Complex in Addition to Beta-Catenin. Proc. Natl. Acad. Sci. USA 2009, 106, 5165–5170. [Google Scholar] [CrossRef] [PubMed]
- Latres, E.; Chiaur, D.S.; Pagano, M. The Human F Box Protein Beta-Trcp Associates with the Cul1/Skp1 Complex and Regulates the Stability of Beta-Catenin. Oncogene 1999, 18, 849–854. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Semenov, M.; Tamai, K.; Zeng, X. LDL Receptor-Related Proteins 5 and 6 in Wnt/Beta-Catenin Signaling: Arrows Point the Way. Development 2004, 131, 1663–1677. [Google Scholar] [CrossRef]
- Kishida, S.; Yamamoto, H.; Hino, S.; Ikeda, S.; Kishida, M.; Kikuchi, A. DIX Domains of Dvl and Axin Are Necessary for Protein Interactions and Their Ability to Regulate β-Catenin Stability. Mol. Cell. Biol. 1999, 19, 4414. [Google Scholar] [CrossRef]
- Cadigan, K.M.; Waterman, M.L. TCF/LEFs and Wnt Signaling in the Nucleus. Cold Spring Harb. Perspect. Biol. 2012, 4, a007906. [Google Scholar] [CrossRef]
- Kikuchi, A.; Kishida, S.; Yamamoto, H. Regulation of Wnt Signaling by Protein-Protein Interaction and Post-Translational Modifications. Exp. Mol. Med. 2006, 38, 1–10. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Z.; Tang, Y.; Xiao, Q. The Involvement of Noncanonical Wnt Signaling in Cancers. Biomed. Pharmacother. 2021, 133, 110946. [Google Scholar] [CrossRef] [PubMed]
- Zhan, T.; Rindtorff, N.; Boutros, M. Wnt Signaling in Cancer. Oncogene 2017, 36, 1461–1473. [Google Scholar] [CrossRef] [PubMed]
- Dreyer, C.A.; VanderVorst, K.; Carraway, K.L. Vangl as a Master Scaffold for Wnt/Planar Cell Polarity Signaling in Development and Disease. Front. Cell Dev. Biol. 2022, 10, 887100. [Google Scholar] [CrossRef] [PubMed]
- Devenport, D. The Cell Biology of Planar Cell Polarity. J. Cell Biol. 2014, 207, 171–179. [Google Scholar] [CrossRef]
- Gao, B.; Song, H.; Bishop, K.; Elliot, G.; Garrett, L.; English, M.A.; Andre, P.; Robinson, J.; Sood, R.; Minami, Y.; et al. Wnt Signaling Gradients Establish Planar Cell Polarity by Inducing Vangl2 Phosphorylation through Ror2. Dev. Cell 2011, 20, 163–176. [Google Scholar] [CrossRef]
- Yang, W.; Garrett, L.; Feng, D.; Elliott, G.; Liu, X.; Wang, N.; Wong, Y.M.; Choi, N.T.; Yang, Y.; Gao, B. Wnt-Induced Vangl2 Phosphorylation Is Dose-Dependently Required for Planar Cell Polarity in Mammalian Development. Cell Res. 2017, 27, 1466–1484. [Google Scholar] [CrossRef]
- Schlessinger, K.; Hall, A.; Tolwinski, N. Wnt Signaling Pathways Meet Rho GTPases. Genes Dev. 2009, 23, 265–277. [Google Scholar] [CrossRef]
- Wang, J.; Feng, D.; Gao, B. An Overview of Potential Therapeutic Agents Targeting WNT/PCP Signaling. Handb. Exp. Pharmacol. 2021, 269, 175–213. [Google Scholar] [CrossRef]
- Rogers, S.; Scholpp, S. Vertebrate Wnt5a-At the Crossroads of Cellular Signalling. Semin. Cell Dev. Biol. 2022, 125, 3–10. [Google Scholar] [CrossRef]
- Kühl, M.; Sheldahl, L.C.; Park, M.; Miller, J.R.; Moon, R.T. The Wnt/Ca2+ Pathway: A New Vertebrate Wnt Signaling Pathway Takes Shape. Trends Genet. 2000, 16, 279–283. [Google Scholar] [CrossRef]
- Kühl, M. The WNT/Calcium Pathway: Biochemical Mediators, Tools and Future Requirements. Front. Biosci. J. Virtual Libr. 2004, 9, 967–974. [Google Scholar] [CrossRef] [PubMed]
- De, A. Wnt/Ca2+ Signaling Pathway: A Brief Overview. Acta Biochim. Biophys. Sin. 2011, 43, 745–756. [Google Scholar] [CrossRef] [PubMed]
- Vuong, L.T.; Mlodzik, M. Different Strategies by Distinct Wnt-Signaling Pathways in Activating a Nuclear Transcriptional Response. Curr. Top. Dev. Biol. 2022, 149, 59–89. [Google Scholar] [CrossRef] [PubMed]
- Correction: The CBio Cancer Genomics Portal: An Open Platform for Exploring Multidimensional Cancer Genomics Data. Cancer Discov. 2012, 2, 960. [CrossRef]
- Joslyn, G.; Carlson, M.; Thliveris, A.; Albertsen, H.; Gelbert, L.; Samowitz, W.; Groden, J.; Stevens, J.; Spirio, L.; Robertson, M. Identification of Deletion Mutations and Three New Genes at the Familial Polyposis Locus. Cell 1991, 66, 601–613. [Google Scholar] [CrossRef]
- Segditsas, S.; Tomlinson, I. Colorectal Cancer and Genetic Alterations in the Wnt Pathway. Oncogene 2006, 25, 7531–7537. [Google Scholar] [CrossRef]
- Song, M.; Chan, A.T.; Sun, J. Influence of the Gut Microbiome, Diet, and Environment on Risk of Colorectal Cancer. Gastroenterology 2020, 158, 322–340. [Google Scholar] [CrossRef]
- Afrin, S.; Giampieri, F.; Gasparrini, M.; Forbes-Hernandez, T.Y.; Cianciosi, D.; Reboredo-Rodriguez, P.; Zhang, J.; Manna, P.P.; Daglia, M.; Atanasov, A.G.; et al. Dietary Phytochemicals in Colorectal Cancer Prevention and Treatment: A Focus on the Molecular Mechanisms Involved. Biotechnol. Adv. 2020, 38, 107322. [Google Scholar] [CrossRef]
- Seiwert, N.; Heylmann, D.; Hasselwander, S.; Fahrer, J. Mechanism of Colorectal Carcinogenesis Triggered by Heme Iron from Red Meat. Biochim. Biophys. Acta-Rev. Cancer 2020, 1873, 188334. [Google Scholar] [CrossRef]
- Liu, F.; Zuo, X.; Liu, Y.; Deguchi, Y.; Moussalli, M.J.; Chen, W.; Yang, P.; Wei, B.; Tan, L.; Lorenzi, P.L.; et al. Suppression of Membranous LRP5 Recycling, Wnt/Beta-Catenin Signaling, and Colon Carcinogenesis by 15-LOX-1 Peroxidation of Linoleic Acid in PI3P. Cell Rep. 2020, 32, 108049. [Google Scholar] [CrossRef]
- Kim, T.-Y.; Kim, S.; Kim, Y.; Lee, Y.-S.; Lee, S.; Lee, S.-H.; Kweon, M.-N. A High-Fat Diet Activates the BAs-FXR Axis and Triggers Cancer-Associated Fibroblast Properties in the Colon. Cell. Mol. Gastroenterol. Hepatol. 2022, 13, 1141–1159. [Google Scholar] [CrossRef] [PubMed]
- Rijo-Ferreira, F.; Takahashi, J.S. Genomics of Circadian Rhythms in Health and Disease. Genome Med. 2019, 11, 82. [Google Scholar] [CrossRef] [PubMed]
- Koike, N.; Yoo, S.-H.; Huang, H.-C.; Kumar, V.; Lee, C.; Kim, T.-K.; Takahashi, J.S. Transcriptional Architecture and Chromatin Landscape of the Core Circadian Clock in Mammals. Science 2012, 338, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Altman, B.J.; Hsieh, A.L.; Sengupta, A.; Krishnanaiah, S.Y.; Stine, Z.E.; Walton, Z.E.; Gouw, A.M.; Venkataraman, A.; Li, B.; Goraksha-Hicks, P.; et al. MYC Disrupts the Circadian Clock and Metabolism in Cancer Cells. Cell Metab. 2015, 22, 1009–1019. [Google Scholar] [CrossRef]
- Chun, S.K.; Fortin, B.M.; Fellows, R.C.; Habowski, A.N.; Verlande, A.; Song, W.A.; Mahieu, A.L.; Lefebvre, A.E.Y.T.; Sterrenberg, J.N.; Velez, L.M.; et al. Disruption of the Circadian Clock Drives Apc Loss of Heterozygosity to Accelerate Colorectal Cancer. Sci. Adv. 2022, 8, eabo2389. [Google Scholar] [CrossRef] [PubMed]
- Bennedsen, A.L.B.; Furbo, S.; Bjarnsholt, T.; Raskov, H.; Gogenur, I.; Kvich, L. The Gut Microbiota Can Orchestrate the Signaling Pathways in Colorectal Cancer. Apmis 2022, 130, 121–139. [Google Scholar] [CrossRef]
- Cai, J.; Sun, L.; Gonzalez, F.J. Gut Microbiota-Derived Bile Acids in Intestinal Immunity, Inflammation, and Tumorigenesis. Cell Host Microbe 2022, 30, 289–300. [Google Scholar] [CrossRef]
- Liu, X.; Lu, R.; Wu, S.; Sun, J. Salmonella Regulation of Intestinal Stem Cells through the Wnt/Beta-Catenin Pathway. FEBS Lett. 2010, 584, 911–916. [Google Scholar] [CrossRef]
- Wang, J.; Lu, R.; Fu, X.; Dan, Z.; Zhang, Y.-G.; Chang, X.; Liu, Q.; Xia, Y.; Liu, X.; Sun, J. Novel Regulatory Roles of Wnt1 in Infection-Associated Colorectal Cancer. Neoplasia 2018, 20, 499–509. [Google Scholar] [CrossRef]
- Iftekhar, A.; Berger, H.; Bouznad, N.; Heuberger, J.; Boccellato, F.; Dobrindt, U.; Hermeking, H.; Sigal, M.; Meyer, T.F. Genomic Aberrations after Short-Term Exposure to Colibactin-Producing E. Coli Transform Primary Colon Epithelial Cells. Nat. Commun. 2021, 12, 1003. [Google Scholar] [CrossRef]
- Rubinstein, M.R.; Baik, J.E.; Lagana, S.M.; Han, R.P.; Raab, W.J.; Sahoo, D.; Dalerba, P.; Wang, T.C.; Han, Y.W. Fusobacterium Nucleatum Promotes Colorectal Cancer by Inducing Wnt/β-Catenin Modulator Annexin A1. EMBO Rep. 2019, 20, e47638. [Google Scholar] [CrossRef] [PubMed]
- Rubinstein, M.R.; Wang, X.; Liu, W.; Hao, Y.; Cai, G.; Han, Y.W. Fusobacterium Nucleatum Promotes Colorectal Carcinogenesis by Modulating E-Cadherin/β-Catenin Signaling via Its FadA Adhesin. Cell Host Microbe 2013, 14, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Huang, J.; Yu, T.; Fang, X.; Lou, L.; Xin, S.; Ji, L.; Jiang, F.; Lou, Y. Fusobacterium Nucleatum Promotes the Progression of Colorectal Cancer Through Cdk5-Activated Wnt/Beta-Catenin Signaling. Front. Microbiol. 2021, 11, 545251. [Google Scholar] [CrossRef]
- Cavallucci, V.; Palucci, I.; Fidaleo, M.; Mercuri, A.; Masi, L.; Emoli, V.; Bianchetti, G.; Fiori, M.E.; Bachrach, G.; Scaldaferri, F.; et al. Proinflammatory and Cancer-Promoting Pathobiont Fusobacterium Nucleatum Directly Targets Colorectal Cancer Stem Cells. Biomolecules 2022, 12, 1256. [Google Scholar] [CrossRef]
- Akimoto, N.; Ugai, T.; Zhong, R.; Hamada, T.; Fujiyoshi, K.; Giannakis, M.; Wu, K.; Cao, Y.; Ng, K.; Ogino, S. Rising Incidence of Early-Onset Colorectal Cancer-a Call to Action. Nat. Rev. Clin. Oncol. 2021, 18, 230–243. [Google Scholar] [CrossRef] [PubMed]
- Dejea, C.M.; Fathi, P.; Craig, J.M.; Boleij, A.; Taddese, R.; Geis, A.L.; Wu, X.; Shields, C.E.D.; Hechenbleikner, E.M.; Huso, D.L.; et al. Patients with Familial Adenomatous Polyposis Harbor Colonic Biofilms Containing Tumorigenic Bacteria. Science 2018, 359, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Wang, X.; Sun, J.; Lin, W.; Chen, L.; Liu, S.; Wu, X.; Shi, L.; Xu, P.; Cai, X.; et al. IRF3 Prevents Colorectal Tumorigenesis via Inhibiting the Nuclear Translocation of Beta-Catenin. Nat. Commun. 2020, 11, 5762. [Google Scholar] [CrossRef]
- Fantini, M.C.; Guadagni, I. From Inflammation to Colitis-Associated Colorectal Cancer in Inflammatory Bowel Disease: Pathogenesis and Impact of Current Therapies. Dig. Liver Dis. 2021, 53, 558–565. [Google Scholar] [CrossRef]
- Kaplan, G.G. The Global Burden of IBD: From 2015 to 2025. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 720–727. [Google Scholar] [CrossRef]
- Foersch, S.; Neurath, M.F. Colitis-Associated Neoplasia: Molecular Basis and Clinical Translation. Cell. Mol. Life Sci. 2014, 71, 3523–3535. [Google Scholar] [CrossRef]
- Cooper, H.S.; Everley, L.; Chang, W.C.; Pfeiffer, G.; Lee, B.; Murthy, S.; Clapper, M.L. The Role of Mutant Apc in the Development of Dysplasia and Cancer in the Mouse Model of Dextran Sulfate Sodium-Induced Colitis. Gastroenterology 2001, 121, 1407–1416. [Google Scholar] [CrossRef]
- Nagao-Kitamoto, H.; Kitamoto, S.; Kamada, N. Inflammatory Bowel Disease and Carcinogenesis. Cancer Metastasis Rev. 2022, 41, 301–316. [Google Scholar] [CrossRef] [PubMed]
- Ben-Neriah, Y.; Karin, M. Inflammation Meets Cancer, with NF-Kappa B as the Matchmaker. Nat. Immunol. 2011, 12, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Lasry, A.; Zinger, A.; Ben-Neriah, Y. Inflammatory Networks Underlying Colorectal Cancer. Nat. Immunol. 2016, 17, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Dyson, J.K.; Rutter, M.D. Colorectal Cancer in Inflammatory Bowel Disease: What Is the Real Magnitude of the Risk? World J. Gastroenterol. 2012, 18, 3839–3848. [Google Scholar] [CrossRef]
- Triantafillidis, J.K.; Nasioulas, G.; Kosmidis, P.A. Colorectal Cancer and Inflammatory Bowel Disease: Epidemiology, Risk Factors, Mechanisms of Carcinogenesis and Prevention Strategies. Anticancer Res. 2009, 29, 2727–2737. [Google Scholar]
- Rajamaki, K.; Taira, A.; Katainen, R.; Valimaki, N.; Kuosmanen, A.; Plaketti, R.-M.; Seppala, T.T.; Ahtiainen, M.; Wirta, E.-V.; Vartiainen, E.; et al. Genetic and Epigenetic Characteristics of Inflammatory Bowel Disease-Associated Colorectal Cancer. Gastroenterology 2021, 161, 592–607. [Google Scholar] [CrossRef]
- Tang, F.; Cao, F.; Lu, C.; He, X.; Weng, L.; Sun, L. Dvl2 Facilitates the Coordination of NF-Kappa B and Wnt Signaling to Promote Colitis-Associated Colorectal Progression. Cancer Sci. 2022, 113, 565–575. [Google Scholar] [CrossRef]
- Zheng, L.; Duan, S.-L.; Wen, X.-L.; Dai, Y.-C. Molecular Regulation after Mucosal Injury and Regeneration in Ulcerative Colitis. Front. Mol. Biosci. 2022, 9, 1015049. [Google Scholar] [CrossRef]
- Guenther, C.; Neumann, H.; Neurath, M.F.; Becker, C. Apoptosis, Necrosis and Necroptosis: Cell Death Regulation in the Intestinal Epithelium. Gut 2013, 62, 1062–1071. [Google Scholar] [CrossRef]
- Qi, J.; Cui, D.; Wu, Q.-N.; Zhao, Q.; Chen, Z.-H.; Li, L.; Birchmeier, W.; Yu, Y.; Tao, R. Targeting Wnt/β-Catenin Signaling by TET1/FOXO4 Inhibits Metastatic Spreading and Self-Renewal of Cancer Stem Cells in Gastric Cancer. Cancers 2022, 14, 3232. [Google Scholar] [CrossRef]
- Sahebdel, F.; Parvaneh Tafreshi, A.; Arefian, E.; Roussa, E.; Nadri, S.; Zeynali, B. Wnt/β-Catenin Signaling Pathway Is Involved in Early Dopaminergic Differentiation of Trabecular Meshwork-Derived Mesenchymal Stem Cells. J. Cell. Biochem. 2022, 123, 1120–1129. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Dong, H.; Wang, P.; Xu, Z.; Xian, J.; Chen, J.; Wu, H.; Lou, Y.; Lin, D.; Zhong, B. IL-36 γ and IL-36Ra Reciprocally Regulate Colon Inflammation and Tumorigenesis by Modulating the Cell–Matrix Adhesion Network and Wnt Signaling. Adv. Sci. 2022, 9, 2103035. [Google Scholar] [CrossRef] [PubMed]
- Boege, Y.; Malehmir, M.; Healy, M.E.; Bettermann, K.; Lorentzen, A.; Vucur, M.; Ahuja, A.K.; Bohm, F.; Mertens, J.C.; Shimizu, Y.; et al. A Dual Role of Caspase-8 in Triggering and Sensing Proliferation-Associated DNA Damage, a Key Determinant of Liver Cancer Development. Cancer Cell 2017, 32, 342–359. [Google Scholar] [CrossRef] [PubMed]
- Healy, M.E.; Boege, Y.; Hodder, M.C.; Bohm, F.; Malehmir, M.; Scherr, A.-L.; Jetzer, J.; Chan, L.K.; Parrotta, R.; Jacobs, K.; et al. MCL1 Is Required for Maintenance of Intestinal Homeostasis and Prevention of Carcinogenesis in Mice. Gastroenterology 2020, 159, 183–199. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Komai, K.; Mise-Omata, S.; Iizuka-Koga, M.; Noguchi, Y.; Kondo, T.; Sakai, R.; Matsuo, K.; Nakayama, T.; Yoshie, O.; et al. Brain Regulatory T Cells Suppress Astrogliosis and Potentiate Neurological Recovery. Nature 2019, 565, 246–250. [Google Scholar] [CrossRef] [PubMed]
- Sefik, E.; Geva-Zatorsky, N.; Oh, S.; Konnikova, L.; Zemmour, D.; McGuire, A.M.; Burzyn, D.; Ortiz-Lopez, A.; Lobera, M.; Yang, J.; et al. Individual Intestinal Symbionts Induce a Distinct Population of RORγ + Regulatory T Cells. Science 2015, 349, 993–997. [Google Scholar] [CrossRef]
- van Loosdregt, J.; Fleskens, V.; Tiemessen, M.M.; Mokry, M.; van Boxtel, R.; Meerding, J.; Pals, C.E.G.M.; Kurek, D.; Baert, M.R.M.; Delemarre, E.M.; et al. Canonical Wnt Signaling Negatively Modulates Regulatory T Cell Function. Immunity 2013, 39, 298–310. [Google Scholar] [CrossRef]
- Samstein, R.M.; Arvey, A.; Josefowicz, S.Z.; Peng, X.; Reynolds, A.; Sandstrom, R.; Neph, S.; Sabo, P.; Kim, J.M.; Liao, W.; et al. Foxp3 Exploits a Pre-Existent Enhancer Landscape for Regulatory T Cell Lineage Specification. Cell 2012, 151, 153–166. [Google Scholar] [CrossRef]
- Xing, S.; Gai, K.; Li, X.; Shao, P.; Zeng, Z.; Zhao, X.; Zhao, X.; Chen, X.; Paradee, W.J.; Meyerholz, D.K.; et al. Tcf1 and Lef1 Are Required for the Immunosuppressive Function of Regulatory T Cells. J. Exp. Med. 2019, 216, 847–866. [Google Scholar] [CrossRef]
- Quandt, J.; Arnovitz, S.; Haghi, L.; Woehlk, J.; Mohsin, A.; Okoreeh, M.; Mathur, P.S.; Emmanuel, A.O.; Osman, A.; Krishnan, M.; et al. Wnt–β-Catenin Activation Epigenetically Reprograms Treg Cells in Inflammatory Bowel Disease and Dysplastic Progression. Nat. Immunol. 2021, 22, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, R.; van Driel, M.S.; Vermeulen, L.; van Neerven, S.M. Intestinal Stem Cell Dynamics in Homeostasis and Cancer. Trends Cancer 2022, 8, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Pashirzad, M.; Sathyapalan, T.; Sheikh, A.; Kesharwani, P.; Sahebkar, A. Cancer Stem Cells: An Overview of the Pathophysiological and Prognostic Roles in Colorectal Cancer. Process Biochem. 2022, 115, 19–29. [Google Scholar] [CrossRef]
- Batlle, E.; Clevers, H. Cancer Stem Cells Revisited. Nat. Med. 2017, 23, 1124–1134. [Google Scholar] [CrossRef] [PubMed]
- Ricci-Vitiani, L.; Fabrizi, E.; Palio, E.; De Maria, R. Colon Cancer Stem Cells. J. Mol. Med. 2009, 87, 1097. [Google Scholar] [CrossRef] [PubMed]
- Silva, V.R.; Santos, L.d.S.; Dias, R.B.; Quadros, C.A.; Bezerra, D.P. Emerging Agents That Target Signaling Pathways to Eradicate Colorectal Cancer Stem Cells. Cancer Commun. 2021, 41, 1275–1313. [Google Scholar] [CrossRef] [PubMed]
- Katoh, M.; Katoh, M. WNT Signaling and Cancer Stemness. Essays Biochem. 2022, 66, 319–331. [Google Scholar] [CrossRef]
- Sato, T.; van Es, J.H.; Snippert, H.J.; Stange, D.E.; Vries, R.G.; van den Born, M.; Barker, N.; Shroyer, N.F.; van de Wetering, M.; Clevers, H. Paneth Cells Constitute the Niche for Lgr5 Stem Cells in Intestinal Crypts. Nature 2011, 469, 415–418. [Google Scholar] [CrossRef]
- McCarthy, N.; Kraiczy, J.; Shivdasani, R.A. Cellular and Molecular Architecture of the Intestinal Stem Cell Niche. Nat. Cell Biol. 2020, 22, 1033–1041. [Google Scholar] [CrossRef]
- van Neerven, S.M.; Vermeulen, L. The Interplay between Intrinsic and Extrinsic Wnt Signaling in Controlling Intestinal Transformation. Differentiation 2019, 108, 17–23. [Google Scholar] [CrossRef]
- Nusse, R.; Clevers, H. Wnt/β-Catenin Signaling, Disease, and Emerging Therapeutic Modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef] [PubMed]
- Cheung, P.; Xiol, J.; Dill, M.T.; Yuan, W.-C.; Panero, R.; Roper, J.; Osorio, F.G.; Maglic, D.; Li, Q.; Gurung, B.; et al. Regenerative Reprogramming of the Intestinal Stem Cell State via Hippo Signaling Suppresses Metastatic Colorectal Cancer. Cell Stem Cell 2020, 27, 590–604.e9. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Duronio, G.N.; Yang, Y.; Bala, P.; Hebbar, P.; Spisak, S.; Sahgal, P.; Singh, H.; Zhang, Y.; Xie, Y.; et al. An Enhancer-Driven Stem Cell-Like Program Mediated by SOX9 Blocks Intestinal Differentiation in Colorectal Cancer. Gastroenterology 2022, 162, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Tomic, G.; Morrissey, E.; Kozar, S.; Ben-Moshe, S.; Hoyle, A.; Azzarelli, R.; Kemp, R.; Chilamakuri, C.S.R.; Itzkovitz, S.; Philpott, A.; et al. Phospho-Regulation of ATOH1 Is Required for Plasticity of Secretory Progenitors and Tissue Regeneration. Cell Stem Cell 2018, 23, 436–443.e7. [Google Scholar] [CrossRef] [PubMed]
- de Sousa e Melo, F.; Kurtova, A.V.; Harnoss, J.M.; Kljavin, N.; Hung, J.; Anderson, J.E.; Storm, E.E.; Modrusan, Z.; Koeppen, H.; Dijkgraaf, G.J.P.; et al. A Distinct Role for Lgr5(+) Stem Cells in Primary and Metastatic Colon Cancer. Nature 2017, 543, 676–680. [Google Scholar] [CrossRef]
- Sansom, O.J.; Meniel, V.S.; Muncan, V.; Phesse, T.J.; Wilkins, J.A.; Reed, K.R.; Vass, J.K.; Athineos, D.; Clevers, H.; Clarke, A.R. Myc Deletion Rescues Apc Deficiency in the Small Intestine. Nature 2007, 446, 676–679. [Google Scholar] [CrossRef]
- Zhang, H.; Lin, M.; Dong, C.; Tang, Y.; An, L.; Ju, J.; Wen, F.; Chen, F.; Wang, M.; Wang, W.; et al. An MST4-p Beta-Catenin(Thr40) Signaling Axis Controls Intestinal Stem Cell and Tumorigenesis. Adv. Sci. 2021, 8, 2004850. [Google Scholar] [CrossRef]
- Angus-Hill, M.L.; Elbert, K.M.; Hidalgo, J.; Capecchi, M.R. T-Cell Factor 4 Functions as a Tumor Suppressor Whose Disruption Modulates Colon Cell Proliferation and Tumorigenesis. Proc. Natl. Acad. Sci. USA 2011, 108, 4914–4919. [Google Scholar] [CrossRef]
- Hrckulak, D.; Janeckova, L.; Lanikova, L.; Kriz, V.; Horazna, M.; Babosova, O.; Vojtechova, M.; Galuskova, K.; Sloncova, E.; Korinek, V. Wnt Effector TCF4 Is Dispensable for Wnt Signaling in Human Cancer Cells. Genes 2018, 9, 439. [Google Scholar] [CrossRef]
- Novoa Diaz, M.B.; Martin, M.J.; Gentili, C. Tumor Microenvironment Involvement in Colorectal Cancer Progression via Wnt/Beta-Catenin Pathway: Providing Understanding of the Complex Mechanisms of Chemoresistance. World J. Gastroenterol. 2022, 28, 3027–3046. [Google Scholar] [CrossRef]
- Kalra, H.; Gangoda, L.; Fonseka, P.; Chitti, S.V.; Liem, M.; Keerthikumar, S.; Samuel, M.; Boukouris, S.; Al Saffar, H.; Collins, C.; et al. Extracellular Vesicles Containing Oncogenic Mutant β-Catenin Activate Wnt Signalling Pathway in the Recipient Cells. J. Extracell. Vesicles 2019, 8, 1690217. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.-J.; Jang, G.-B.; Lee, H.-Y.; Park, S.-R.; Kim, J.-Y.; Nam, J.-S.; Hong, I.-S. The Wnt/β-Catenin Signaling/Id2 Cascade Mediates the Effects of Hypoxia on the Hierarchy of Colorectal-Cancer Stem Cells. Sci. Rep. 2016, 6, 22966. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Yang, M.; Li, Y.; Yang, F.; Feng, Y. Exosomes Derived from Hypoxic Colorectal Cancer Cells Transfer Wnt4 to Normoxic Cells to Elicit a Prometastatic Phenotype. Int. J. Biol. Sci. 2018, 14, 2094–2102. [Google Scholar] [CrossRef]
- Cheng, R.; Sun, B.; Liu, Z.; Zhao, X.; Qi, L.; Li, Y.; Gu, Q. Wnt5a Suppresses Colon Cancer by Inhibiting Cell Proliferation and Epithelial-Mesenchymal Transition. J. Cell. Physiol. 2014, 229, 1908–1917. [Google Scholar] [CrossRef]
- Liu, Q.; Yang, C.; Wang, S.; Shi, D.; Wei, C.; Song, J.; Lin, X.; Dou, R.; Bai, J.; Xiang, Z.; et al. Wnt5a-Induced M2 Polarization of Tumor-Associated Macrophages via IL-10 Promotes Colorectal Cancer Progression. Cell Commun. Signal. 2020, 18, 51. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Song, J.; Pan, Y.; Shi, D.; Yang, C.; Wang, S.; Xiong, B. Wnt5a/CaMKII/ERK/CCL2 Axis Is Required for Tumor-Associated Macrophages to Promote Colorectal Cancer Progression. Int. J. Biol. Sci. 2020, 16, 1023–1034. [Google Scholar] [CrossRef]
- Kasprzak, A. Angiogenesis-Related Functions of Wnt Signaling in Colorectal Carcinogenesis. Cancers 2020, 12, 3601. [Google Scholar] [CrossRef]
- Baeriswyl, V.; Christofori, G. The Angiogenic Switch in Carcinogenesis. Semin. Cancer Biol. 2009, 19, 329–337. [Google Scholar] [CrossRef]
- Hillen, F.; Griffioen, A.W. Tumour Vascularization: Sprouting Angiogenesis and Beyond. Cancer Metastasis Rev. 2007, 26, 489–502. [Google Scholar] [CrossRef]
- Watnick, R.S. The Role of the Tumor Microenvironment in Regulating Angiogenesis. Cold Spring Harb. Perspect. Med. 2012, 2, a006676. [Google Scholar] [CrossRef]
- Kramer, N.; Schmöllerl, J.; Unger, C.; Nivarthi, H.; Rudisch, A.; Unterleuthner, D.; Scherzer, M.; Riedl, A.; Artaker, M.; Crncec, I.; et al. Autocrine WNT2 Signaling in Fibroblasts Promotes Colorectal Cancer Progression. Oncogene 2017, 36, 5460–5472. [Google Scholar] [CrossRef] [PubMed]
- Unterleuthner, D.; Neuhold, P.; Schwarz, K.; Janker, L.; Neuditschko, B.; Nivarthi, H.; Crncec, I.; Kramer, N.; Unger, C.; Hengstschlaeger, M.; et al. Cancer-Associated Fibroblast-Derived WNT2 Increases Tumor Angiogenesis in Colon Cancer. Angiogenesis 2020, 23, 159–177. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Shu, H.; Zhang, L.; Xiong, J. Long Noncoding RNA GAS5 Inhibits Angiogenesis and Metastasis of Colorectal Cancer through the Wnt/Β-catenin Signaling Pathway. J. Cell. Biochem. 2019, 120, 6937–6951. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Zhao, H.; Qu, B.; Chu, X.; Xin, X.; Zhang, Q.; Li, W.; Yang, S. TRIM24 Promotes Colorectal Cancer Cell Progression via the Wnt/Beta-Catenin Signaling Pathway Activation. Am. J. Transl. Res. 2022, 14, 831–848. [Google Scholar] [PubMed]
- Yang, D.; Li, Q.; Shang, R.; Yao, L.; Wu, L.; Zhang, M.; Zhang, L.; Xu, M.; Lu, Z.; Zhou, J.; et al. WNT4 Secreted by Tumor Tissues Promotes Tumor Progression in Colorectal Cancer by Activation of the Wnt/Beta-Catenin Signalling Pathway. J. Exp. Clin. Cancer Res. 2020, 39, 251. [Google Scholar] [CrossRef]
- EL Andaloussi, S.; Mäger, I.; Breakefield, X.O.; Wood, M.J.A. Extracellular Vesicles: Biology and Emerging Therapeutic Opportunities. Nat. Rev. Drug Discov. 2013, 12, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Yamada, N.; Kuranaga, Y.; Kumazaki, M.; Shinohara, H.; Taniguchi, K.; Akao, Y. Colorectal Cancer Cell-Derived Extracellular Vesicles Induce Phenotypic Alteration of T Cells into Tumor-Growth Supporting Cells with Transforming Growth Factor-Β1-Mediated Suppression. Oncotarget 2016, 7, 27033–27043. [Google Scholar] [CrossRef] [PubMed]
- Hong, B.S.; Cho, J.-H.; Kim, H.; Choi, E.-J.; Rho, S.; Kim, J.; Kim, J.H.; Choi, D.-S.; Kim, Y.-K.; Hwang, D.; et al. Colorectal Cancer Cell-Derived Microvesicles Are Enriched in Cell Cycle-Related MRNAs That Promote Proliferation of Endothelial Cells. BMC Genom. 2009, 10, 556. [Google Scholar] [CrossRef]
- Ji, H.; Greening, D.W.; Barnes, T.W.; Lim, J.W.; Tauro, B.J.; Rai, A.; Xu, R.; Adda, C.; Mathivanan, S.; Zhao, W.; et al. Proteome Profiling of Exosomes Derived from Human Primary and Metastatic Colorectal Cancer Cells Reveal Differential Expression of Key Metastatic Factors and Signal Transduction Components. Proteomics 2013, 13, 1672–1686. [Google Scholar] [CrossRef]
- Huang, Z.; Feng, Y. Exosomes Derived from Hypoxic Colorectal Cancer Cells Promote Angiogenesis Through Wnt4-Induced β-Catenin Signaling in Endothelial Cells. Oncol. Res. 2017, 25, 651–661. [Google Scholar] [CrossRef]
- Winkler, J.; Abisoye-Ogunniyan, A.; Metcalf, K.J.; Werb, Z. Concepts of Extracellular Matrix Remodelling in Tumour Progression and Metastasis. Nat. Commun. 2020, 11, 5120. [Google Scholar] [CrossRef] [PubMed]
- Vu, T.; Datta, P. Regulation of EMT in Colorectal Cancer: A Culprit in Metastasis. Cancers 2017, 9, 171. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Lai, Q.; He, C.; Fang, Y.; Yan, Q.; Zhang, Y.; Wang, X.; Gu, C.; Wang, Y.; Ye, L.; et al. RUNX1 Promotes Tumour Metastasis by Activating the Wnt/β-Catenin Signalling Pathway and EMT in Colorectal Cancer. J. Exp. Clin. Cancer Res. 2019, 38, 334. [Google Scholar] [CrossRef] [PubMed]
- McCrea, P.D.; Gottardi, C.J. Beyond β-Catenin: Prospects for a Larger Catenin Network in the Nucleus. Nat. Rev. Mol. Cell Biol. 2016, 17, 55–64. [Google Scholar] [CrossRef]
- Basu, S.; Cheriyamundath, S.; Ben-Ze’ev, A. Cell–Cell Adhesion: Linking Wnt/β-Catenin Signaling with Partial EMT and Stemness Traits in Tumorigenesis. F1000Research 2018, 7, 1488. [Google Scholar] [CrossRef]
- Valenta, T.; Hausmann, G.; Basler, K. The Many Faces and Functions of β-Catenin. EMBO J. 2012, 31, 2714–2736. [Google Scholar] [CrossRef]
- Zhu, Y.; Gan, Y.; Zou, R.; Sha, H.; Lu, Y.; Zhang, Y.; Feng, J. RNF128 Suppresses the Malignancy of Colorectal Cancer Cells via Inhibition of Wnt/Beta-Catenin Signaling. Am. J. Transl. Res. 2021, 13, 13567–13578. [Google Scholar]
- Meng, F.; Liu, X.; Lin, C.; Xu, L.; Liu, J.; Zhang, P.; Zhang, X.; Song, J.; Yan, Y.; Ren, Z.; et al. SMYD2 Suppresses APC2 Expression to Activate the Wnt/Beta-Catenin Pathway and Promotes Epithelial-Mesenchymal Transition in Colorectal Cancer. Am. J. Cancer Res. 2020, 10, 997–1011. [Google Scholar]
- Tang, Q.; Chen, J.; Di, Z.; Yuan, W.; Zhou, Z.; Liu, Z.; Han, S.; Liu, Y.; Ying, G.; Shu, X.; et al. TM4SF1 Promotes EMT and Cancer Stemness via the Wnt/Beta-Catenin/SOX2 Pathway in Colorectal Cancer. J. Exp. Clin. Cancer Res. 2020, 39, 232. [Google Scholar] [CrossRef]
- Cai, Z.; Mei, Y.; Jiang, X.; Shi, X. WDR74 Promotes Proliferation and Metastasis in Colorectal Cancer Cells through Regulating the Wnt/Beta-Catenin Signaling Pathway. Open Life Sci. 2021, 16, 920–929. [Google Scholar] [CrossRef]
- Zhang, L.; Ren, C.; Yang, Z.; Gong, L.; Wang, C.; Feng, M.; Guan, W. Forkhead Box S1 Mediates Epithelial-Mesenchymal Transition through the Wnt/Beta-Catenin Signaling Pathway to Regulate Colorectal Cancer Progression. J. Transl. Med. 2022, 20, 327. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, L.; Xu, X.; Han, P.; Wu, J.; Tian, X.; Li, M. FOXO4 Inhibits the Migration and Metastasis of Colorectal Cancer by Regulating the APC2/Beta-Catenin Axis. Front. Cell Dev. Biol. 2021, 9, 659731. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Gao, T.; Xie, J.; Huang, Z.; Zhang, X.; Yang, F.; Qi, W.; Yang, Z.; Zhou, T.; Gao, G.; et al. FUBP1 Promotes Colorectal Cancer Stemness and Metastasis via DVL1-Mediated Activation of Wnt/Beta-Catenin Signaling. Mol. Oncol. 2021, 15, 3490–3512. [Google Scholar] [CrossRef]
- Faubert, B.; Solmonson, A.; DeBerardinis, R.J. Metabolic Reprogramming and Cancer Progression. Science 2020, 368, eaaw5473. [Google Scholar] [CrossRef]
- Martinez-Reyes, I.; Chandel, N.S. Cancer Metabolism: Looking Forward. Nat. Rev. Cancer 2021, 21, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, K.; Oi, R.; Okuzaki, D.; Motooka, D.; Shinohara, M.; Nojima, S.; Morii, E. Mitochondrial Matrix Protein C14orf159 Attenuates Colorectal Cancer Metastasis by Suppressing Wnt/Beta-Catenin Signalling. Br. J. Cancer 2021, 125, 1699–1711. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, K.; Nojima, S.; Tahara, S.; Kurashige, M.; Kawasaki, K.; Hori, Y.; Taniguchi, M.; Umakoshi, Y.; Okuzaki, D.; Wada, N.; et al. Serine Racemase Enhances Growth of Colorectal Cancer by Producing Pyruvate from Serine. Nat. Metab. 2020, 2, 81–96. [Google Scholar] [CrossRef]
- Zhu, Y.-M.; Li, Q.; Gao, X.-Z.; Meng, X.; Sun, L.-L.; Shi, Y.; Lu, E.-T.; Zhang, Y. C14orf159 Suppresses Gastric Cancer Cells’ Invasion and Proliferation by Inactivating ERK Signaling. Cancer Manag. Res. 2019, 11, 1717–1723. [Google Scholar] [CrossRef]
- Bakir, B.; Chiarella, A.M.; Pitarresi, J.R.; Rustgi, A.K. EMT, MET, Plasticity, and Tumor Metastasis. Trends Cell Biol. 2020, 30, 764–776. [Google Scholar] [CrossRef]
- Bimonte, S.; Barbieri, A.; Leongito, M.; Piccirillo, M.; Giudice, A.; Pivonello, C.; De Angelis, C.; Granata, V.; Palaia, R.; Izzo, F. Curcumin AntiCancer Studies in Pancreatic Cancer. Nutrients 2016, 8, 433. [Google Scholar] [CrossRef]
- Dongre, A.; Weinberg, R.A. New Insights into the Mechanisms of Epithelial-Mesenchymal Transition and Implications for Cancer. Nat. Rev. Mol. Cell Biol. 2019, 20, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Li, Q.; Liu, Y.; Zhang, H.; Wang, Q.; Chen, Y.; Shi, X.; Li, J.; Zhang, H.; Zhang, Y.; et al. Activation of WNT7b Autocrine Eases Metastasis of Colorectal Cancer via Epithelial to Mesenchymal Transition and Predicts Poor Prognosis. BMC Cancer 2021, 21, 180. [Google Scholar] [CrossRef] [PubMed]
- Pergolizzi, M.; Bizzozero, L.; Maione, F.; Maldi, E.; Isella, C.; Macagno, M.; Mariella, E.; Bardelli, A.; Medico, E.; Marchio, C.; et al. The Neuronal Protein Neuroligin 1 Promotes Colorectal Cancer Progression by Modulating the APC/Beta-Catenin Pathway. J. Exp. Clin. Cancer Res. 2022, 41, 266. [Google Scholar] [CrossRef]
- Xu, C.; Ding, Y.; Wang, K.; Hao, M.; Li, H.; Ding, L. Claudin-7 Deficiency Promotes Stemness Properties in Colorectal Cancer through Sox9-Mediated Wnt/Beta-Catenin Signalling. J. Transl. Med. 2021, 19, 311. [Google Scholar] [CrossRef]
- Nie, X.; Liu, H.; Ye, W.; Wei, X.; Fan, L.; Ma, H.; Li, L.; Xue, W.; Qi, W.; Wang, Y.-D.; et al. LRP5 Promotes Cancer Stem Cell Traits and Chemoresistance in Colorectal Cancer. J. Cell. Mol. Med. 2022, 26, 1095–1112. [Google Scholar] [CrossRef] [PubMed]
- VanderVorst, K.; Dreyer, C.A.; Konopelski, S.E.; Lee, H.; Ho, H.-Y.H.; Carraway, K.L. Wnt/PCP Signaling Contribution to Carcinoma Collective Cell Migration and Metastasis. Cancer Res. 2019, 79, 1719–1729. [Google Scholar] [CrossRef] [PubMed]
- Gross, J.C.; Chaudhary, V.; Bartscherer, K.; Boutros, M. Active Wnt Proteins Are Secreted on Exosomes. Nat. Cell Biol. 2012, 14, 1036–1045. [Google Scholar] [CrossRef]
- Wang, F.-W.; Cao, C.-H.; Han, K.; Zhao, Y.-X.; Cai, M.-Y.; Xiang, Z.-C.; Zhang, J.-X.; Chen, J.-W.; Zhong, L.-P.; Huang, Y.; et al. APC-Activated Long Noncoding RNA Inhibits Colorectal Carcinoma Pathogenesis through Reduction of Exosome Production. J. Clin. Investig. 2019, 129, 727–743. [Google Scholar] [CrossRef]
- Le Floch, N.; Rivat, C.; De Wever, O.; Bruyneel, E.; Mareel, M.; Dale, T.; Gespach, C. The Proinvasive Activity of Wnt-2 Is Mediated through a Noncanonical Wnt Pathway Coupled to GSK-3β and C- Jun/AP-1 Signaling. FASEB J. 2005, 19, 144–146. [Google Scholar] [CrossRef]
- Zhang, Y.; Lin, L.; Jin, Y.; Lin, Y.; Cao, Y.; Zheng, C. Overexpression of WNT5B Promotes COLO 205 Cell Migration and Invasion through the JNK Signaling Pathway. Oncol. Rep. 2016, 36, 23–30. [Google Scholar] [CrossRef]
- Ouko, L.; Ziegler, T.R.; Gu, L.H.; Eisenberg, L.M.; Yang, V.M. Wnt11 Signaling Promotes Proliferation, Transformation, and Migration of IEC6 Intestinal Epithelial Cells. J. Biol. Chem. 2004, 279, 26707–26715. [Google Scholar] [CrossRef] [PubMed]
- Ueno, K.; Hazama, S.; Mitomori, S.; Nishioka, M.; Suehiro, Y.; Hirata, H.; Oka, M.; Imai, K.; Dahiya, R.; Hinoda, Y. Down-Regulation of Frizzled-7 Expression Decreases Survival, Invasion and Metastatic Capabilities of Colon Cancer Cells. Br. J. Cancer 2009, 101, 1374–1381. [Google Scholar] [CrossRef] [PubMed]
- Ueno, K.; Hiura, M.; Suehiro, Y.; Hazama, S.; Hirata, H.; Oka, M.; Imai, K.; Dahiya, R.; Hinoda, Y. Frizzled-7 as a Potential Therapeutic Target in Colorectal Cancer. Neoplasia 2008, 10, 697–705. [Google Scholar] [CrossRef]
- Nishioka, M.; Ueno, K.; Hazama, S.; Okada, T.; Sakai, K.; Suehiro, Y.; Okayama, N.; Hirata, H.; Oka, M.; Imai, K.; et al. Possible Involvement of Wnt11 in Colorectal Cancer Progression. Mol. Carcinog. 2013, 52, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Parsons, S.; Maldonado, E.B.; Prasad, V. Comparison of Drugs Used for Adjuvant and Metastatic Therapy of Colon, Breast, and Non-Small Cell Lung Cancers. JAMA Netw. Open 2020, 3, e202488. [Google Scholar] [CrossRef]
- Hirano, H.; Takashima, A.; Hamaguchi, T.; Shida, D.; Kanemitsu, Y. Current Status and Perspectives of Immune Checkpoint Inhibitors for Colorectal Cancer. Jpn. J. Clin. Oncol. 2021, 51, 10–19. [Google Scholar] [CrossRef]
- Zhu, G.-X.; Gao, D.; Shao, Z.-Z.; Chen, L.; Ding, W.-J.; Yu, Q.-F. Wnt/Beta-Catenin Signaling: Causes and Treatment Targets of Drug Resistance in Colorectal Cancer. Mol. Med. Rep. 2021, 23, 105. [Google Scholar] [CrossRef]
- Das, P.K.; Islam, F.; Lam, A.K. The Roles of Cancer Stem Cells and Therapy Resistance in Colorectal Carcinoma. Cells 2020, 9, 1392. [Google Scholar] [CrossRef]
- Yeoh, Y.; Low, T.Y.; Abu, N.; Lee, P.Y. Regulation of Signal Transduction Pathways in Colorectal Cancer: Implications for Therapeutic Resistance. PeerJ 2021, 9, e12338. [Google Scholar] [CrossRef]
- Liu, Y.-S.; Hsu, H.-C.; Tseng, K.-C.; Chen, H.-C.; Chen, S.-J. Lgr5 Promotes Cancer Stemness and Confers Chemoresistance through ABCB1 in Colorectal Cancer. Biomed. Pharmacother. 2013, 67, 791–799. [Google Scholar] [CrossRef]
- Liu, H.; Pan, Y.; Guo, G.; Hu, X.; Huang, K.; Zhang, X. Isolation of Side Population Cells and Detection of ABCG2 from SW480. Chin. J. Cancer Res. 2007, 19, 238–243. [Google Scholar] [CrossRef]
- Park, S.-Y.; Kim, J.-Y.; Jang, G.-B.; Choi, J.-H.; Kim, J.-H.; Lee, C.-J.; Lee, S.; Baek, J.-H.; Park, K.-K.; Kim, J.-M.; et al. Aberrant Activation of the CD45-Wnt Signaling Axis Promotes Stemness and Therapy Resistance in Colorectal Cancer Cells. Theranostics 2021, 11, 8755–8770. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Jiang, H.; Li, W.; Chen, L.; Zhu, W.; Xian, Y.; Han, Z.; Yin, L.; Liu, Y.; Wang, Y.; et al. FOXM1/DVL2/Snail Axis Drives Metastasis and Chemoresistance of Colorectal Cancer. Aging 2020, 12, 24424–24440. [Google Scholar] [CrossRef]
- Zhang, W.; Kong, L.; Zhu, H.; Sun, D.; Han, Q.; Yan, B.; Cui, Z.; Zhang, W.; Zhang, S.; Kang, X.; et al. Retinoic Acid-Induced 2 (RAI2) Is a Novel Antagonist of Wnt/Beta-Catenin Signaling Pathway and Potential Biomarker of Chemosensitivity in Colorectal Cancer. Front. Oncol. 2022, 12, 805290. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.; Yamada-Okabe, H.; Suzuki, M.; Natori, O.; Kato, A.; Matsubara, K.; Chen, Y.J.; Yamazaki, M.; Funahashi, S.; Yoshida, K.; et al. LGR5-Positive Colon Cancer Stem Cells Interconvert with Drug-Resistant LGR5-Negative Cells and Are Capable of Tumor Reconstitution. Stem Cells 2012, 30, 2631–2644. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Settleman, J. EMT, Cancer Stem Cells and Drug Resistance: An Emerging Axis of Evil in the War on Cancer. Oncogene 2010, 29, 4741–4751. [Google Scholar] [CrossRef]
- Zinzi, L.; Contino, M.; Cantore, M.; Capparelli, E.; Leopoldo, M.; Colabufo, N.A. ABC Transporters in CSCs Membranes as a Novel Target for Treating Tumor Relapse. Front. Pharmacol. 2014, 5, 163. [Google Scholar] [CrossRef]
- Kugimiya, N.; Nishimoto, A.; Hosoyama, T.; Ueno, K.; Enoki, T.; Li, T.-S.; Hamano, K. The C-MYC-ABCB5 Axis Plays a Pivotal Role in 5-Fluorouracilresistance in Human Colon Cancer Cells. J. Cell. Mol. Med. 2015, 19, 1569–1581. [Google Scholar] [CrossRef]
- Chikazawa, N.; Tanaka, H.; Tasaka, T.; Nakamura, M.; Tanaka, M.; Onishi, H.; Katano, M. Inhibition of Wnt Signaling Pathway Decreases Chemotherapy-Resistant Side-Population Colon Cancer Cells. Anticancer Res. 2010, 30, 2041–2048. [Google Scholar]
- Liang, Y.; Wang, B.; Chen, S.; Ye, Z.; Chai, X.; Li, R.; Li, X.; Kong, G.; Li, Y.; Zhang, X.; et al. Beta-1 Syntrophin (SNTB1) Regulates Colorectal Cancer Progression and Stemness via Regulation of the Wnt/Beta-Catenin Signaling Pathway. Ann. Transl. Med. 2021, 9, 1016. [Google Scholar] [CrossRef]
- Catalano, V.; Turdo, A.; Di Franco, S.; Dieli, F.; Todaro, M.; Stassi, G. Tumor and Its Microenvironment: A Synergistic Interplay. Semin. Cancer Biol. 2013, 23, 522–532. [Google Scholar] [CrossRef]
- Sleeman, J.P. The Metastatic Niche and Stromal Progression. Cancer Metastasis Rev. 2012, 31, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Mierke, C.T. The Matrix Environmental and Cell Mechanical Properties Regulate Cell Migration and Contribute to the Invasive Phenotype of Cancer Cells. Rep. Prog. Phys. 2019, 82, 064602. [Google Scholar] [CrossRef] [PubMed]
- Czekay, R.-P.; Cheon, D.-J.; Samarakoon, R.; Kutz, S.M.; Higgins, P.J. Cancer-Associated Fibroblasts: Mechanisms of Tumor Progression and Novel Therapeutic Targets. Cancers 2022, 14, 1231. [Google Scholar] [CrossRef]
- Hu, Y.-B.; Yan, C.; Mu, L.; Mil, Y.-L.; Zhao, H.; Hu, H.; Li, X.-L.; Tao, D.-D.; Wu, Y.-Q.; Gong, J.-P.; et al. Exosomal Wnt-Induced Dedifferentiation of Colorectal Cancer Cells Contributes to Chemotherapy Resistance. Oncogene 2019, 38, 1951–1965. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Campisi, J.; Higano, C.; Beer, T.M.; Porter, P.; Coleman, I.; True, L.; Nelson, P.S. Treatment-Induced Damage to the Tumor Microenvironment Promotes Prostate Cancer Therapy Resistance through WNT16B. Nat. Med. 2012, 18, 1359–1368. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhu, D.; Chen, F.; Qian, M.; Wei, H.; Chen, W.; Xu, J. SFRP2 Augments WNT16B Signaling to Promote Therapeutic Resistance in the Damaged Tumor Microenvironment. Oncogene 2016, 35, 4321–4334. [Google Scholar] [CrossRef]
- Takada, K.; Zhu, D.; Bird, G.H.; Sukhdeo, K.; Zhao, J.-J.; Mani, M.; Lemieux, M.; Carrasco, D.E.; Ryan, J.; Horst, D.; et al. Targeted Disruption of the BCL9/Beta-Catenin Complex Inhibits Oncogenic Wnt Signaling. Sci. Transl. Med. 2012, 4, 148ra117. [Google Scholar] [CrossRef]
- Dong, S.; Liang, S.; Cheng, Z.; Zhang, X.; Luo, L.; Li, L.; Zhang, W.; Li, S.; Xu, Q.; Zhong, M.; et al. ROS/PI3K/Akt and Wnt/Beta-Catenin Signalings Activate HIF-1 Alpha-Induced Metabolic Reprogramming to Impart 5-Fluorouracil Resistance in Colorectal Cancer. J. Exp. Clin. Cancer Res. 2022, 41, 15. [Google Scholar] [CrossRef]
- Liu, X.; Su, K.; Sun, X.; Jiang, Y.; Wang, L.; Hu, C.; Zhang, C.; Lu, M.; Du, X.; Xing, B. Sec62 Promotes Stemness and Chemoresistance of Human Colorectal Cancer through Activating Wnt/Beta-Catenin Pathway. J. Exp. Clin. Cancer Res. 2021, 40, 132. [Google Scholar] [CrossRef]
- Li, H.; Chen, J.; Liu, J.; Lai, Y.; Huang, S.; Zheng, L.; Fan, N. CPT2 Downregulation Triggers Stemness and Oxaliplatin Resistance in Colorectal Cancer via the ROS/Wnt/-Beta Catenin-Induced Glycolytic Metabolism. Exp. Cell Res. 2021, 409, 112892. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhao, C.; Wang, G.; Chen, J.; Ju, S.; Huang, J.; Wang, X. SNORD1C Maintains Stemness and 5-FU Resistance by Activation of Wnt Signaling Pathway in Colorectal Cancer. Cell Death Discov. 2022, 8, 200. [Google Scholar] [CrossRef] [PubMed]
- Kishore, C.; Bhadra, P. Current Advancements and Future Perspectives of Immunotherapy in Colorectal Cancer Research. Eur. J. Pharmacol. 2021, 893, 173819. [Google Scholar] [CrossRef]
- Yaghoubi, N.; Soltani, A.; Ghazvini, K.; Hassanian, S.M.; Hashemy, S.I. PD-1/PD-L1 Blockade as a Novel Treatment for Colorectal Cancer. Biomed. Pharmacother. 2019, 110, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Payandeh, Z.; Khalili, S.; Somi, M.H.; Mard-Soltani, M.; Baghbanzadeh, A.; Hajiasgharzadeh, K.; Samadi, N.; Baradaran, B. PD-1/PD-L1-Dependent Immune Response in Colorectal Cancer. J. Cell. Physiol. 2020, 235, 5461–5475. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Jin, L.; Chen, P.; Li, D.; Gao, W.; Dong, G. Colorectal Cancer Immunotherapy-Recent Progress and Future Directions. Cancer Lett. 2022, 545, 215816. [Google Scholar] [CrossRef] [PubMed]
- Fountzilas, E.; Kurzrock, R.; Vo, H.H.; Tsimberidou, A.-M. Wedding of Molecular Alterations and Immune Checkpoint Blockade: Genomics as a Matchmaker. J. Natl. Cancer Inst. 2021, 113, 1634–1647. [Google Scholar] [CrossRef]
- Kim, C.W.; Chon, H.J.; Kim, C. Combination Immunotherapies to Overcome Intrinsic Resistance to Checkpoint Blockade in Microsatellite Stable Colorectal Cancer. Cancers 2021, 13, 4906. [Google Scholar] [CrossRef]
- Ganesh, S.; Shui, X.; Craig, K.P.; Park, J.; Wang, W.; Brown, B.D.; Abrams, M.T. RNAi-Mediated β-Catenin Inhibition Promotes T Cell Infiltration and Antitumor Activity in Combination with Immune Checkpoint Blockade. Mol. Ther. J. Am. Soc. Gene Ther. 2018, 26, 2567–2579. [Google Scholar] [CrossRef]
- Spranger, S.; Dai, D.; Horton, B.; Gajewski, T.F. Tumor-Residing Batf3 Dendritic Cells Are Required for Effector T Cell Trafficking and Adoptive T Cell Therapy. Cancer Cell 2017, 31, 711–723.e4. [Google Scholar] [CrossRef]
- Berraondo, P.; Ochoa, M.C.; Olivera, I.; Melero, I. Immune Desertic Landscapes in Hepatocellular Carcinoma Shaped by β-Catenin Activation. Cancer Discov. 2019, 9, 1003–1005. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Yu, X.; Xue, L.; Ge, X.; Zhao, W.; Peng, W. Intrinsic β-Catenin Signaling Suppresses CD8+ T-Cell Infiltration in Colorectal Cancer. Biomed. Pharmacother. Biomed. Pharmacother. 2019, 115, 108921. [Google Scholar] [CrossRef] [PubMed]
- Trujillo, J.A.; Sweis, R.F.; Bao, R.; Luke, J.J. T Cell–Inflamed versus Non-T Cell–Inflamed Tumors: A Conceptual Framework for Cancer Immunotherapy Drug Development and Combination Therapy Selection. Cancer Immunol. Res. 2018, 6, 990–1000. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Tian, T.; Kalland, K.-H.; Ke, X.; Qu, Y. Targeting Wnt/β-Catenin Signaling for Cancer Immunotherapy. Trends Pharmacol. Sci. 2018, 39, 648–658. [Google Scholar] [CrossRef]
- Pai, S.G.; Carneiro, B.A.; Mota, J.M.; Costa, R.; Leite, C.A.; Barroso-Sousa, R.; Kaplan, J.B.; Chae, Y.K.; Giles, F.J. Wnt/Beta-Catenin Pathway: Modulating Anticancer Immune Response. J. Hematol. Oncol. 2017, 10, 1–12. [Google Scholar] [CrossRef]
- Tang, Y.; Nan, N.; Gui, C.; Zhou, X.; Jiang, W.; Zhou, X. Blockage of PD-L1 by FERMT3-Mediated Wnt/Beta-Catenin Signalling Regulates Chemoresistance and Immune Evasion of Colorectal Cancer Cells. Clin. Exp. Pharmacol. Physiol. 2022, 49, 988–997. [Google Scholar] [CrossRef]
- Stanczak, A.; Stec, R.; Bodnar, L.; Olszewski, W.; Cichowicz, M.; Kozlowski, W.; Szczylik, C.; Pietrucha, T.; Wieczorek, M.; Lamparska-Przybysz, M. Prognostic Significance of Wnt-1, β-Catenin and E-Cadherin Expression in Advanced Colorectal Carcinoma. Pathol. Oncol. Res. 2011, 17, 955–963. [Google Scholar] [CrossRef]
- Matly, A.; Quinn, J.A.; McMillan, D.C.; Park, J.H.; Edwards, J. The Relationship between β-Catenin and Patient Survival in Colorectal Cancer Systematic Review and Meta-Analysis. Crit. Rev. Oncol./Hematol. 2021, 163, 103337. [Google Scholar] [CrossRef]
- Kriegl, L.; Horst, D.; Reiche, J.A.; Engel, J.; Kirchner, T.; Jung, A. LEF-1 and TCF4 Expression Correlate Inversely with Survival in Colorectal Cancer. J. Transl. Med. 2010, 8, 123. [Google Scholar] [CrossRef]
- Escobar, D.; Bushara, O.; Sun, L.; Liao, J.; Yang, G.-Y. Clinicopathologic Characteristics of FBXW7-Mutated Colorectal Adenocarcinoma and Association with Aberrant Beta-Catenin Localization. Hum. Pathol. 2022, 119, 51–58. [Google Scholar] [CrossRef]
- Dahlmann, M.; Monks, A.; Harris, E.D.; Kobelt, D.; Osterland, M.; Khaireddine, F.; Herrmann, P.; Kemmner, W.; Burock, S.; Walther, W.; et al. Combination of Wnt/Beta-Catenin Targets S100A4 and DKK1 Improves Prognosis of Human Colorectal Cancer. Cancers 2022, 14, 37. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Lv, G.; Xu, B.; Jiang, B. Overexpression of UBE2M through Wnt/Beta-Catenin Signaling Is Associated with Poor Prognosis and Chemotherapy Resistance in Colorectal Cancer. Transl. Cancer Res. 2020, 9, 5614–5625. [Google Scholar] [CrossRef] [PubMed]
- Shu, Y.; He, C.; Xiong, Y.; Lan, J.; Song, R. Paired-like Homeodomain Transcription Factor 2 Strengthens Chemoresistance in Colorectal Cancer by Activating the Wnt/Beta-Catenin Axis. Neoplasma 2021, 68, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Guinney, J.; Dienstmann, R.; Wang, X.; de Reyniès, A.; Schlicker, A.; Soneson, C.; Marisa, L.; Roepman, P.; Nyamundanda, G.; Angelino, P.; et al. The Consensus Molecular Subtypes of Colorectal Cancer. Nat. Med. 2015, 21, 1350–1356. [Google Scholar] [CrossRef] [PubMed]
- Caspi, M.; Wittenstein, A.; Kazelnik, M.; Shor-Nareznoy, Y.; Rosin-Arbesfeld, R. Therapeutic Targeting of the Oncogenic Wnt Signaling Pathway for Treating Colorectal Cancer and Other Colonic Disorders. Adv. Drug Deliv. Rev. 2021, 169, 118–136. [Google Scholar] [CrossRef] [PubMed]
- Lenz, H.; Kahn, M. Safely Targeting Cancer Stem Cells via Selective Catenin Coactivator Antagonism. Cancer Sci. 2014, 105, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Bendell, J.; Eckhardt, G.S.; Hochster, H.S.; Morris, V.K.; Strickler, J.; Kapoun, A.M.; Wang, M.; Xu, L.; McGuire, K.; Dupont, J.; et al. Initial Results from a Phase 1a/b Study of OMP-131R10, a First-in-Class Anti-RSPO3 Antibody, in Advanced Solid Tumors and Previously Treated Metastatic Colorectal Cancer (CRC). Eur. J. Cancer 2016, 69, S29–S30. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Y.; Li, X. Advances of Wnt Signalling Pathway in Colorectal Cancer. Cells 2023, 12, 447. https://doi.org/10.3390/cells12030447
Zhu Y, Li X. Advances of Wnt Signalling Pathway in Colorectal Cancer. Cells. 2023; 12(3):447. https://doi.org/10.3390/cells12030447
Chicago/Turabian StyleZhu, Yaoyao, and Xia Li. 2023. "Advances of Wnt Signalling Pathway in Colorectal Cancer" Cells 12, no. 3: 447. https://doi.org/10.3390/cells12030447
APA StyleZhu, Y., & Li, X. (2023). Advances of Wnt Signalling Pathway in Colorectal Cancer. Cells, 12(3), 447. https://doi.org/10.3390/cells12030447