1. Introduction
Most RNA viruses transmit their genetic information via single-stranded RNA and cause diseases such as the common cold, COVID-19, influenza, viral meningitis, encephalitis, heart failure, late onset diabetes (type1) and others. The financial burden by these viral diseases is tremendous.
One of these relevant viral pathogens is coxsackievirus B3 (CVB3), which belongs to the picornaviridae family and is a member of the Enterovirus genus. CVB3 causes meningitis [
1,
2] and pancreatitis [
3,
4], but most clinical relevance is attributed to pathogenesis of myocarditis in children and juveniles [
5,
6,
7,
8,
9]. Out of the six different serotypes (CVB1–6), CVB3 is the predominant agent of viral myocarditis being responsible for 20–40% of acute heart infections. Furthermore, prolonged viral persistence potentially ends up in dilated cardiomyopathy (DCM) [
10]. The clinical phenotype depends on unclear host factors and differs widely from unapparent inflammation and rapid viral clearance to acute congestive heart failure or viral persistence leading to chronic inflammation with progressive chronic disease [
11,
12].
Despite the increasing and profound knowledge regarding viral myocarditis, it is still a challenge to identify further host factors, which contribute to the development of chronic myocarditis. A solid diagnosis and treatment of patients demands a better understanding of the inflammatory process and the impact of the viral proteins on the host cell [
13,
14]. Murine models of CVB3-induced myocarditis help in gaining information about the interactions between the host cell and viral proteins, respectively, to determine the affected host factors. Most experimental approaches use susceptible mouse strains such as A.BY/SnJ mice or BALB/c H-2
d and provide unique insight into the acute infection phase or subacute phase of the inflammation [
15,
16]. CVB3 is a relevant example causing acute viral myocarditis and the viral life cycle of CVB3 has been extensively studied [
17]. However, the development of chronic infection or the determining of host-signaling factors involved in the pathogenesis of DCM need to be explored further. Murine models mimicking enteroviral infection running a chronic course are accompanied by virus RNA persistence, limited viral plus-strand RNA synthesis and protein expression and chronic inflammation [
18].
According to present knowledge, different mechanisms might be involved in progressive pathogenesis of viral myocarditis. For instance, the direct cell death of cardiomyocytes is induced by CVB3 due to cell lysis. Furthermore, an overshoot reaction of infiltrating immune cells is anticipated [
17]. Cardiac remodeling due to prolonged chronic inflammation is caused by dysregulation of different matrix metalloproteinases (MMP) depending on the examined mouse strain [
16,
19,
20,
21]. In addition, an enhanced generation of reactive oxygen species (ROS) induced by CVB3 within the cytoplasm results in an increased permeability of the mitochondrial outer membrane and leads to a destabilization of the mitochondria with severe consequences to the physiology of cardiomyocytes [
22]. Yet, there remains a missing link between pathogenesis, cardiac remodeling and contractile dysfunction.
Recently, we established an inducible CVB3-transgenic non-infectious human iPSC model [
23]. In human iPSC-derived cardiomyocytes, we found reduced and irregular beating rates, impaired mitochondrial function as indicated by increased ROS-production and altered β–adrenergic repolarization response in four-week-maturated cardiomyocytes expressing the non-infectious CVB3 variant, CVB3DVP0, which lacks correct VP0 capsid protein precursor cleavage and so hinders the correct formation of the viral capsid [
24].
Here, we present an analogous transgenic mouse model that allows homogenous virus protein expression in the whole heart mimicking chronic myocarditis leading to a mild DCM accompanied by decreased cardiac output and ejection fraction in vivo. We investigated the active and passive features of cardiac CVB3ΔVP0 expression and characterized this novel transgenic model in vivo and in vitro. Our data provide prominent evidence for exclusive, CVB3-protein expression-based regulation of mitochondrial degeneration and dysfunction. Associated pathophysiological heart parameters resulted from a decreased contractile force and the disruption of excitation-contraction coupling especially in the late phase of contraction under stress as a consequence of an impaired calcium homeostasis. Together, our data show how the viral protein expression in cardiomyocytes determines the outcome of chronic viral myocarditis in vivo and in vitro.
2. Materials and Methods
2.1. Transgenic CVB3ΔVP0 Mice
Generation of heterozygous transgenic CVB3ΔVP0 mice (TG) has been reported previously [
25]. Breeding was performed in accordance with German guidelines. Housing and animal experiments were approved by the Landesamt für Natur, Umwelt und Verbraucherschutz NRW and were performed in accordance with the guidelines of the Westfälische Wilhelms-University Münster. Unless not otherwise quoted, all described experiments were performed on male mice at 25 ± 2 weeks of age and the genotype of every animal was confirmed by polymerase chain reaction (PCR). All mice were on C57Bl/6 J background and littermates or imported mice (Charles River) were used as wildtype (WT) controls. All animals were provided food and water ad libitum.
2.2. Cardiac Magnetic-Resonance-Imaging (CMR)
In vivo MR imaging was performed at 9.4 T on a Bruker BioSpec 94/20 (Ettlingen, Germany) equipped with a 1 T/m gradient system and ParaVision 5.1 operating software including the IntraGate software for sequence acquisition and reconstruction. A 35 mm volume coil was used for data acquisition. To measure the systolic heart function, a stack of contiguous short-axis slices was acquired to cover the entire right and left ventricles by using the self-gated cine FLASH (IntraGate FLASH, Bruker, Ettlingen, Germany) sequence with the following parameters: slice thickness, 1 mm; number of slices, 9; matrix size, 232 × 232; field of view, 30 × 30 mm2; averages, 1; TE, 3 ms; flip angle, 15°; pulse shape, Gaussian; pulse length, 0.5 ms; TR, 7.4 ms; number of repetitions, 100; acquisition bandwidth, 75 kHz and scan time per slice, 2 min 54 s. The diastolic function was measured by using the self-gated cine UTE (IntraGate UTE) sequence with the following parameters: slice thickness, 1 mm; number of slices, 9; matrix size, 156 × 156; field of view, 20 × 20 mm2; averages, 1; TE, 0.314 ms; TR, 6.2 ms; flip angle, 15°; pulse shape, Gaussian; pulse length, 0.3 ms; number of projections, 246; polar undersampling, 2.0; number of movie cycles, 1000; effective time resolution, 2 ms (50–70 frames/cardiac cycle); acquisition bandwidth, 100 kHz and scan time per slice, 25 min 25 s.
During MRI measurements animals were anesthetized with isoflurane (1.5–2.5 vol% isoflurane, 0.7/0.3 air/O2 mixture) and were monitored for core body temperature and respiration rate using an MRI compatible monitoring system (SA Instruments, Stony Brook, NY, USA).
2.3. CMR Analysis
Volumetric analysis of the left and right myocardium and ventricles was performed using Amira (version 5.4.0, Visage Imaging GmbH, Berlin, Germany) and Segment (version v1.9, Medviso AB, Lund, Sweden) software. To determine the systolic function, the regions of interest in the MR FLASH images were selected manually on the end-diastolic and end-systolic frame of each slice by tracing the epicardial and endocardial borders. In order to obtain the global parameters of the entire heart the volume for each frame was calculated as the sum of the area of interest in each slice multiplied by the slice thickness. Stroke volume, cardiac output per minute and the ejection faction were calculated from the blood volume, determined in the end-systolic and end-diastolic phase. Left ventricular (LV) and RV mass was calculated by multiplying the volume of the LV and RV myocardium by the tissue density of 1.05 g/cm3. To analyze diastolic function, a central slice in short axis view of the left ventricle was segmented in high temporal resolution using the segment. The resulting volume–time curve was fitted with a spline function and the corresponding first derivative calculated, providing the early (E) and late (A) filling rate.
2.3.1. Doppler-Echocardiography
Cardiac Doppler echocardiography was performed on a specialized ultrasound system (VEVO 2100; VisualSonics Inc., Ontario, Canada). Following induction of anesthesia with isoflurane (2.0% isoflurane, 98% O2), examinations were performed with the system’s 40 MHz linear probe. ECG-triggered parasternal short-axis views were obtained together with a parasternal long-axis view. Routinely pulsed-wave Doppler signals of the velocity time integral of the mitral valve inflow, the aortic outflow and the pulmonic outflow were obtained. Ultrasound image analyses were performed according to the American Society of Echocardiography’s (ASE) leading edge method (using the VEVO 2100 implemented image analyses software.
2.3.2. Electrocardiographic Recordings
Mice were anaesthetized with 1.5% isoflurane in a mixture of gases containing 33% oxygen and 66% nitrous oxide and spontaneously breathing while body temperature was retained between 37 and 38 °C. Sufficient depth of anesthesia was confirmed by frequent toe pinching. Recordings of six-lead surface ECG were performed using subcutaneous needle electrodes. Signals were recorded unfiltered with scanning rate of 1 k/s (1 kHz). After a baseline ECG for 5 min a β-adrenergic stimulation was achieved by intraperitoneal injection of isoproterenol (2 mg kg−1 body weight; Sigma-Aldrich) dissolved in water and diluted in saline solution. After drug application the ECG recordings were continued and analyzed for an additional 20 min. Data analysis was performed with LabChart Pro 8.0.9 (ADinstruments, Dunedin, New Zealand) and the signal was digitally filtered using a band-pass filter type with high cut-off frequencies of 500 Hz and low cut-off frequencies of 15 Hz. In a timeframe of 15 s, R waves >150 consecutive heartbeats were aligned to average signal ECGs and the results were analyzed and revised manually using LabChart Pro 8.0.9.
2.3.3. Isolation of Ventricular Cardiomyocytes
Sedation and euthanasia were performed by intraperitoneal injection of ketamine (65 mg/kg bodyweight) and xylazine (13 mg/kg bodyweight). Following thoracotomy, the beating heart was excised from the thoracic cavity and the exposed aorta was cannulated in a timely manner. The extracted hearts were fixed to a Langendorff apparatus and perfused for 5 min at a flow velocity of 2.5 mL/min with prewarmed Ca2+-free buffer (buffer A) composed of (in mM) 133.47 NaCl, 3.99 KCl, 1.37 NaH2PO4 × 1 H2O, 1.0 MgCl2 × 6 H2O, 10 HEPES and 10 D(+)-glucose × 1 H2O (pH 7.47 adjusted with NaOH) additionally containing heparin (14.3 IE/mL heparin-sodium) and 10 mM ATPase-inhibitor 2,3-butanedione-monoxime (BDM). The enzymatic digestion was performed for 6 min with buffer A supplemented with 0.8 mg/mL collagenase (230 U/mg CLS-2 Typ II, Worthington, Lakewood, NJ, USA) and 12.5 µM CaCl2. Finally, the digested ventricles were dissected into several small pieces and separated by gravity flow through a nylon mesh into ventricular myocytes.
2.4. Patch-Clamp Technique
Dissociated ventricular cardiomyocytes were electrophysically studied using whole-cell configuration of the patch-clamp technique. Furthermore, 200 µL cell suspension was added to 2 mL extracellular solution in an experimental glass-bottom dish mounted on an inverted microscope (Axiovert 200, Carl Zeiss, Cambridge, United Kingdom). The extracellular solution containing (in mM): 136 NaCl, 5.4 KCl, 0.33 NaH2PO4 × 1 H2O, 1.0 MgCl2 × 6 H2O, 10 HEPES, 10 D(+)-glucose × 1 H2O and 1 CaCl2 × 1 H2O adjusted to pH 7.4 with NaOH. Borosilicate glass capillaries, GB150TF-8P (Harvard Apparatus, Massachusetts, MA, USA), were pulled with a horizontal P-97 Micropipette Puller (Sutter Instruments, Novato, CA, USA) and were used as patch pipettes. Tip resistance was 1.5–3.5 MΩ. The pipette was filled with intracellular solution containing (in mM): 110 CsCl, 30 TEA-Cl, 10 NaCl, 10 HEPES and 5 Mg-ATP adjusted to pH 7.2 with CsOH. For data acquisition, an EPC 10 USB Amplifier (HEKA, Lambrecht, Germany) was connected to a computer using PatchMaster Software Version 1.2. (HEKA). For recording of action potentials (AP), a high resistance seal was established and finally the ruptured patch clamp method was used. After leak current compensation, the cells were stimulated successively with increasing frequencies starting from 0.5 to 10 Hz, respectively, for 1 min. After achieving Ca2+-overload, one action potential was triggered for early detection after depolarizations or delayed after depolarizations.
2.5. Cellular Ca2+ Transient [Ca]I/Calcium Imaging/Shortening
Ventricular cardiomyocytes were incubated for 10 min with the fluorescent Ca2+ indicator Indo-1 (9 µM; Indo-1/AM, Molecular Probes, Thermo Fisher Scientific, Waltham, MA, USA) at room temperature. Indo-1-loaded cells were transferred to a recording chamber filled with Tyrode’s solution (containing in mM: 140 NaCl, 5.8 KCl, 0.5 KH2PO4, 0.4 Na2HPO4 × 2 H2O, 0.9 MgSO4 × 7 H2O, 10 HEPES, 10 glucose, 2 CaCl2, pH 7.3 adjusted with NaOH) and mounted on the stage of an inverted microscope (Eclipse Ti-S, Nikon, Tokyo, Japan). After allowing cells to settle down for 5 min and washing out excess dye by perfusion with Tyrode’s solution, cardiomyocytes were field-stimulated for 10 min at 0.5 Hz (Myopacer, Ionoptix, Milton, MA, USA) before recording started. Intracellular Ca2+ transients and sarcomere shortening were recorded simultaneously (Myocyte Calcium and Contractility System, Ionoptix, Milton, MA, USA). Indo-1 was excited at 340 nm by a continuous light source (UVICO, Rapp OptoElectronic, Wedel, Germany) equipped with respective low- and bandpass filters. The emitted fluorescence (405 and 495 nm) was detected by photomultipliers. Sarcomere shortening was captured with a CCD camera. Data recording and analysis was performed with Ionwizard software Version 6.3. (Ionoptix, Milton, MA, USA). Transients from at least 10 cardiomyocytes were recorded under basal conditions before cells were superfused with Tyrodes’s solution containing 10−6 M isoproterenol. After reaching steady-state conditions, another 10 cardiomyocytes were recorded under exposure to isoproterenol.
2.6. Light Microscopy
After the isolation procedure, ventricular cardiomyocytes were transferred into 35 × 10 mm2 cell culture dishes, settled for 5 min at RT and examined for quality and quantity yield of the isolation. Cell areas and lengths were detected using a microscope (Eclipse Ti-S, Nikon, Tokyo, Japan) with 20× DIC M N2 objective and connected CoolSNAP HQ2 camera.
2.7. Electron Microscopy
Mice were anesthetized and perfused transcardially with PBS for 60 s followed by 2.5% (v/v) glutaraldehyde (GA) for electron microscopy. The heart was dissected en bloc with the aid of a binocular microscope, avoiding any traction of the tissues. After post fixation for 1 day in GA, the tissue was washed in PBS for 10 min. Specimens were then incubated with Dalton for 2 h followed by washing in PBS for 10 min. After dehydration in an ascending ethanol series, the tissue was embedded in Epon, at ratios of propylene oxide and Epon, first 3:1 for 45 min, 1:1 for 1 h and then 1:3 for 2 h and as the last step, pure Epon overnight. Sections were cut with an Ultracut E Reichert-Jung, collected on formvar-coated grids and contrasted with uranyl acetate. Samples were analyzed with a Philips EM 420 (Philips, Holland) transmission electron microscope equipped with a digital CCD camera (Model 792 BioScan; Gatan, USA) and photographic plates system processed with a Ditabis Micron System. Measurements were performed with the aid of ImageJ Version 1.49. The collected data were analyzed with Statistica Version 12.7, by StatSoft. A t-test for independent sampling was performed.
2.8. Genome-Wide Expression Analysis
Isolated ventricular cardiomyocytes (isolated as described above) from three WT and two CVB3ΔVPO mice were shock-frozen in liquid nitrogen and stored at −80 °C till experimental usage. Total RNA was extracted using NucleoSpin RNA (Macherey-Nagel; including on-column DNAse treatment) and quantified by UV-spectrophotometry. Labelled cRNA was prepared from 250 ng DNA-free RNA samples using TotalPrep linear RNA amplification kits (Life Technologies #AMIL1791) and validated for integrity by capillary electrophoresis using an Agilent 2100 Bioanalyzer (Agilent technologies, Boeblingen, Germany). Microarray hybridizations on MouseWG-6 v2.0 Expression BeadChip (6 samples per BeadChip, >45,000 mouse targets per sample) were performed following manufacturer´s recommendations. Cy3-stained chips were scanned using HiScan SQ instrumentation. Acquired data were processed using GenomeStudio software Version 2.0.5. Differential expression analysis was performed between CVB3ΔVPO and WT samples after background subtraction using cubic spline normalization and Illumina Custom error model without FDR. Transcripts with expression fold change ≥2 and significant p-values of ≤0.01 were considered significantly dysregulated in CVB3ΔVPO ventricular cardiomyocytes compared to WT.
2.9. RT-qPCR
The mRNA expression level results obtained from genome-wide expression analysis were validated by using real-time polymerase chain reaction (RT-qPCR). Total RNA from ventricular cardiomyocytes from additional 5 WT and 5 CVB3ΔVPO mice was isolated as described above. Furthermore, 350 ng total RNA was reverse-transcribed into cDNA using oligo-dT
15 primers and M-MLV reverse transcriptase (Affymetrix, Santa Clara, CA, USA) at 42 °C. All primer pairs were designed with Primer3 software 4 February 2015 (
http://primer3.ut.ee/, accessed on 4 February 2015) and validated with respect to efficiency and specificity. Primers used are listed in
Table 1.
Experiments were conducted using iTaq Universal SYBR Green Supermix (Biorad) on an Applied Biosystems 7900HT Fast Real-Time PCR System (Life Technologies, Darmstadt, German). All measurements were performed in duplicate. Relative quantification of expression was performed according to the “ΔΔCt method” [
26] by employing beta-actin (
Actb) for normalization. Two-sided Student’s
t tests were performed on the basis of ΔCt values from biological replicates.
5. Discussion
Persistent CVB3 infection represents a relatively common causative or modulatory basis of chronic myocarditis. CVB3 infections cause a variety of host reactions including immune cell responses and fibrosis in the heart of patients as well as of mice. Acute CVB infection is known to induce lysis of cardiac myocytes. In order to study persistent CVB3 infection we established a transgenic mouse model with CVB3ΔVP0 expression limited to cardiomyocytes. This model expresses the CVB3ΔVP0 variant which generates viral proteins but cannot form capsids due to restricted VP0 precursor protein cleavage and is therefore considered non-infectious [
25]. This unique mouse model mimics a uniformly infected heart with regard to CVB3 protein expression in cardiomyocytes, which allowed screening for functional and mechanistic effects in the whole heart (in vivo) and also in single cardiomyocytes (in vitro).
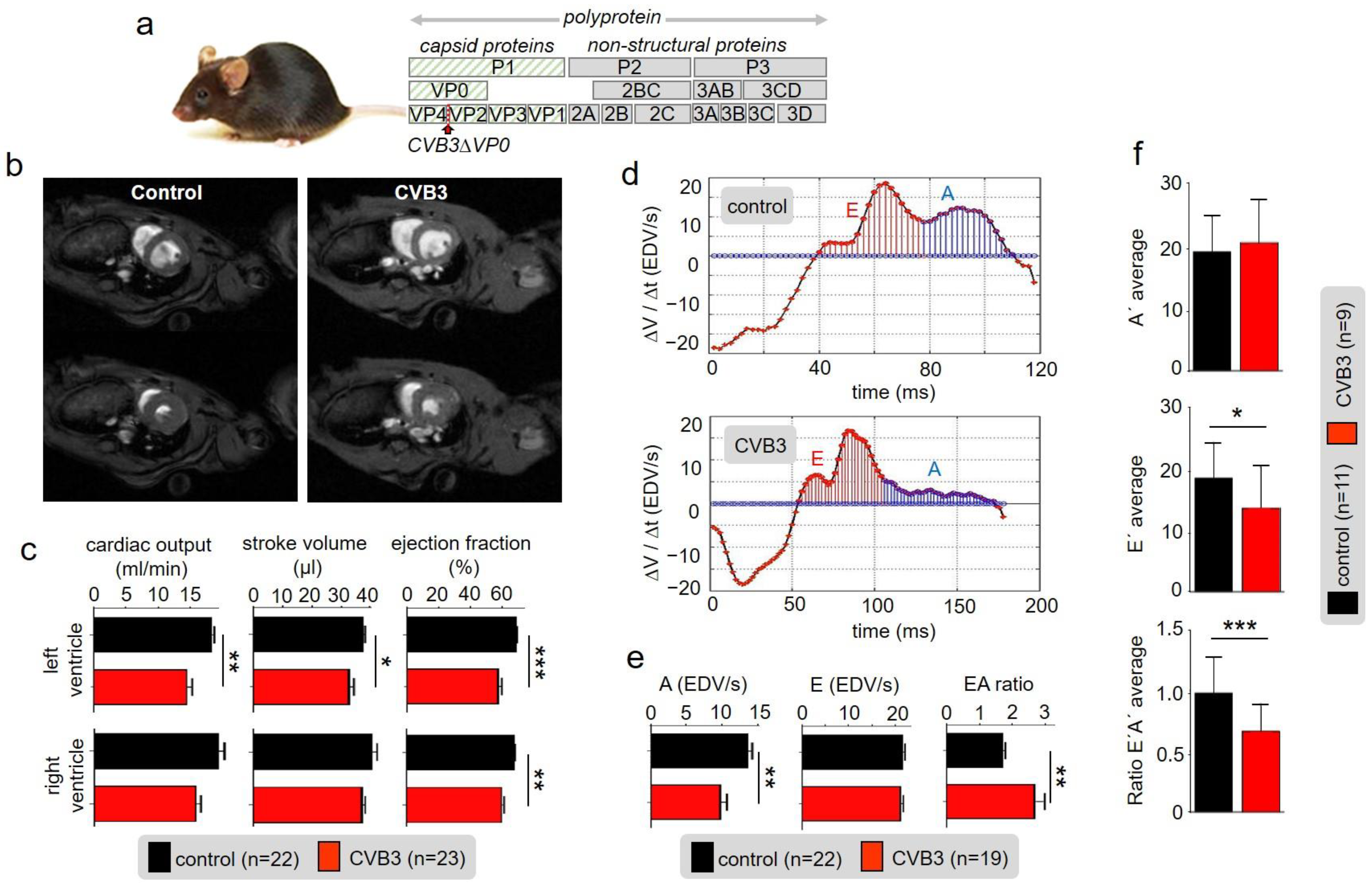
In this study, we found markedly reduced cardiac performance in transgenic CVB3-expressing hearts. Despite normal heart mass and normal ejection fraction, stroke volume and cardiac output were reduced suggesting that force generation in the late-phase diastole is reduced (
Figure 1b,c). The end-systolic blood volume remaining in the ventricles was increased indicating that cardiac elasticity is reduced. Thus, marked changes in cardiac performance are associated with expression of CVB3 proteins in murine cardiomyocytes, which requires further investigation.
Although reduced function of atria was observed here, we concentrated on the ventricles that are responsible for about 85% of cardiac performance. Isolated relaxed ventricular myocytes expressing CVB3 proteins are slightly elongated, supposedly in part resulting from reduced gene expression of whole titin, a key component of cardiomyocyte elasticity (
Figure 3a,b). Reduced cardiomyocyte elasticity will reduce whole heart elasticity and might cause increased end-systolic volume and limit the overall cardiac performance in animals (see
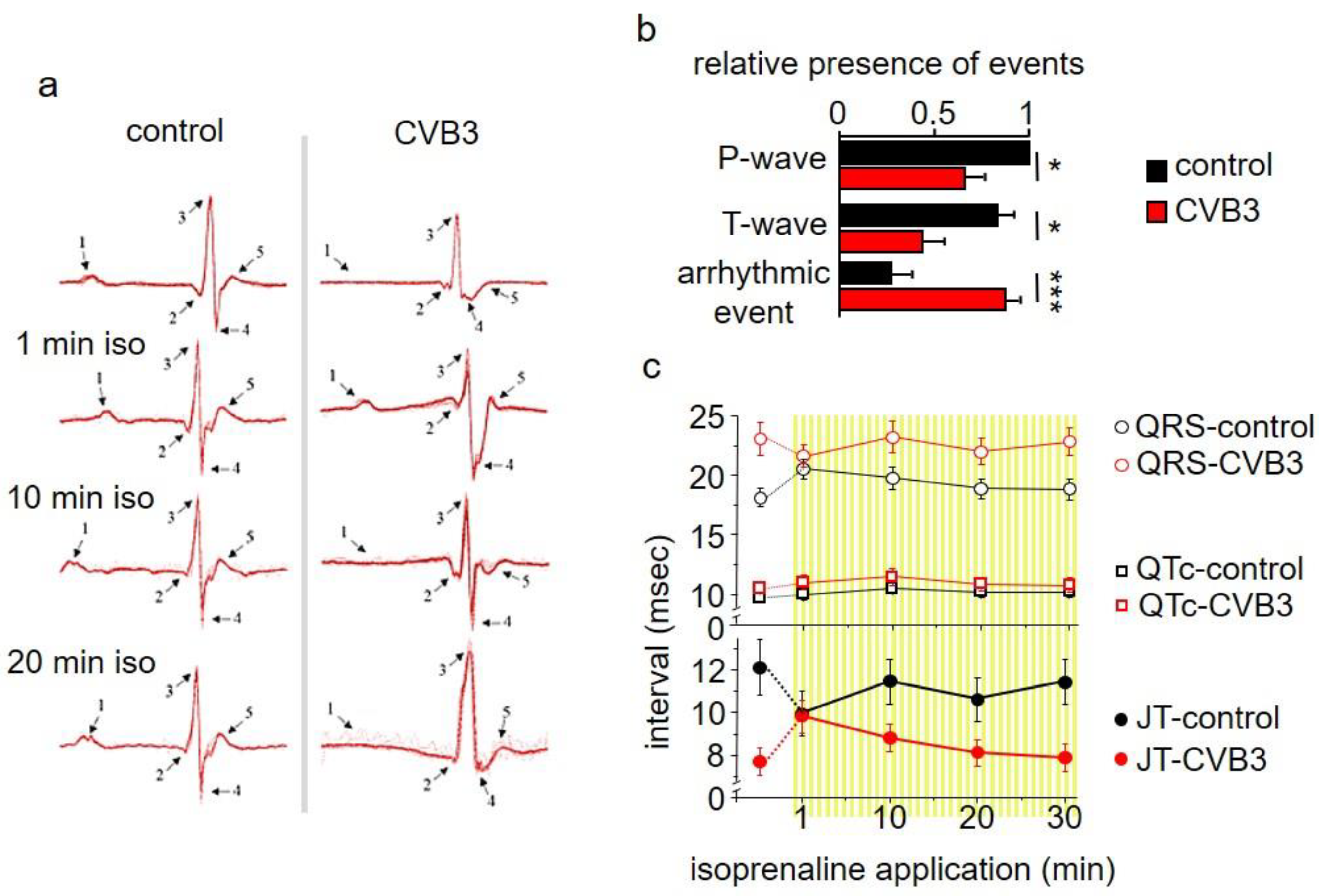
Figure 1b–d). In ECG recordings a mild QTc interval extension was observed (
Figure 2c). This effect could be transiently compensated by β-adrenergic stimulation via isoprenaline application. This re-occurrence of QTc interval extension suggests that transgenic CVB3-expressing animals underwent decompensation. Beside its transiently beneficial effect, β-adrenergic activation triggered distinct changes in ECG morphology and lethal arrhythmias predominantly in transgenic CVB3 protein-expressing mice (
Figure 2a–c). The common physiological acute stress reaction in the heart is mediated by adrenergic signaling and the investigated CVB3 animal model mirrors the clinical situation well, as chronic myocarditis patients are often clinically stable in relaxed situations but often suffer life threatening decompensation under stress situations [
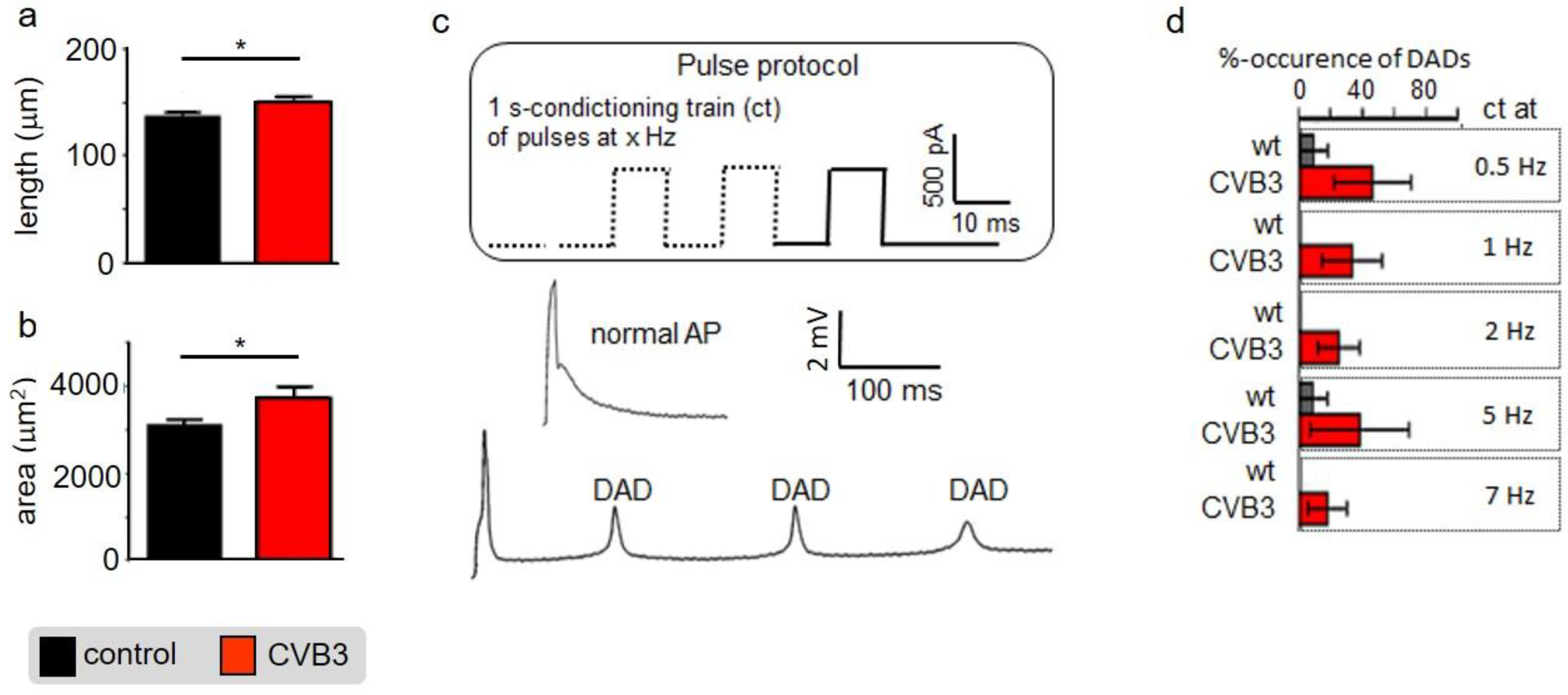
29]. Single cell electrophysiology suggests that CVB3-protein expression sensitized cardiomyocytes for Ca
2+-overload, a pivotal factor in arrhythmogenesis (
Figure 3). Increased (intra-)cellular Ca
2+-flux is one of the major effects of β-adrenergic stimulation and stimulated cytosolic Ca
2+-release from the sarcoplasmic reticulum induces facilitated contraction (positive inotropic effects). However, the increased cytosolic Ca
2+ has to be efficiently removed from the cytosol into the intracellular store units formed by the sarcoplasmic reticulum and to a lesser extent the mitochondria. The Ca
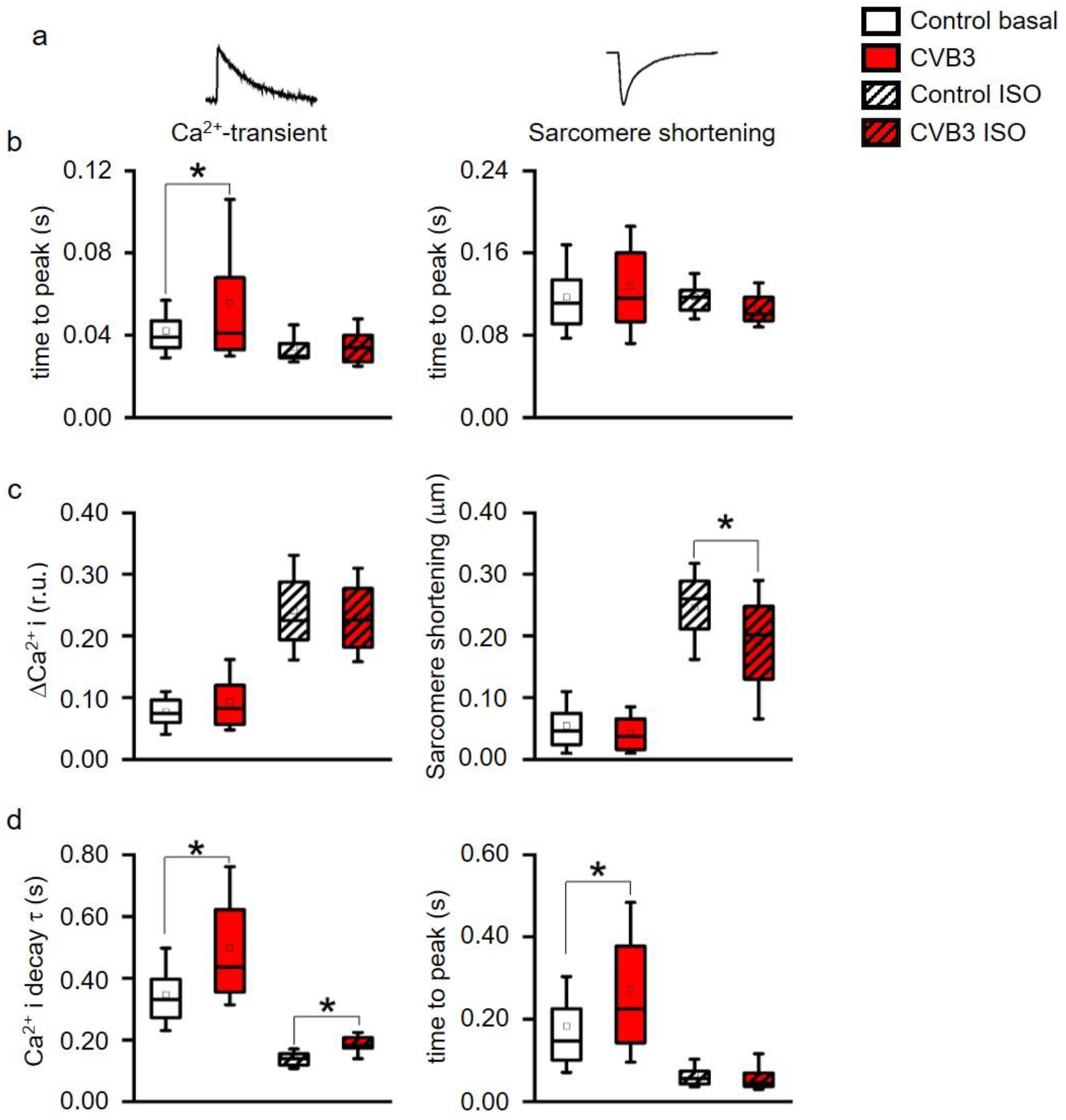
2+-contraction coupling is highly controlled in healthy cardiomyocytes but uncoupled in CVB3ΔVP0 cardiomyocytes, supposedly due to insufficient Ca
2+-reuptake (
Figure 4b–d). Both, normal action potential morphology at rest and inconspicuous immune staining’s of ion channels involved in cardiac repolarizations, suggest no obvious alterations in ion channels in CVB3ΔVP0 cardiomyocytes (
Figure 3,
Supplementary Materials Figure S1). Thus, Ca
2+-overload due to an unknown mechanism probably is the pro-arrhythmic basis of stress triggered arrhythmias under chronic viral heart infection (
Figure 2,
Figure 3 and
Figure 4).
Cellular phenotype and subcellular structure were analyzed via electron microscopy. Cell length and area were significantly increased in CVB3ΔVP0-expressing cells (
Figure 3a,b), whereas mitochondrial area was significantly decreased and sarcomeric structure appeared heavily de-organized (
Figure 5).
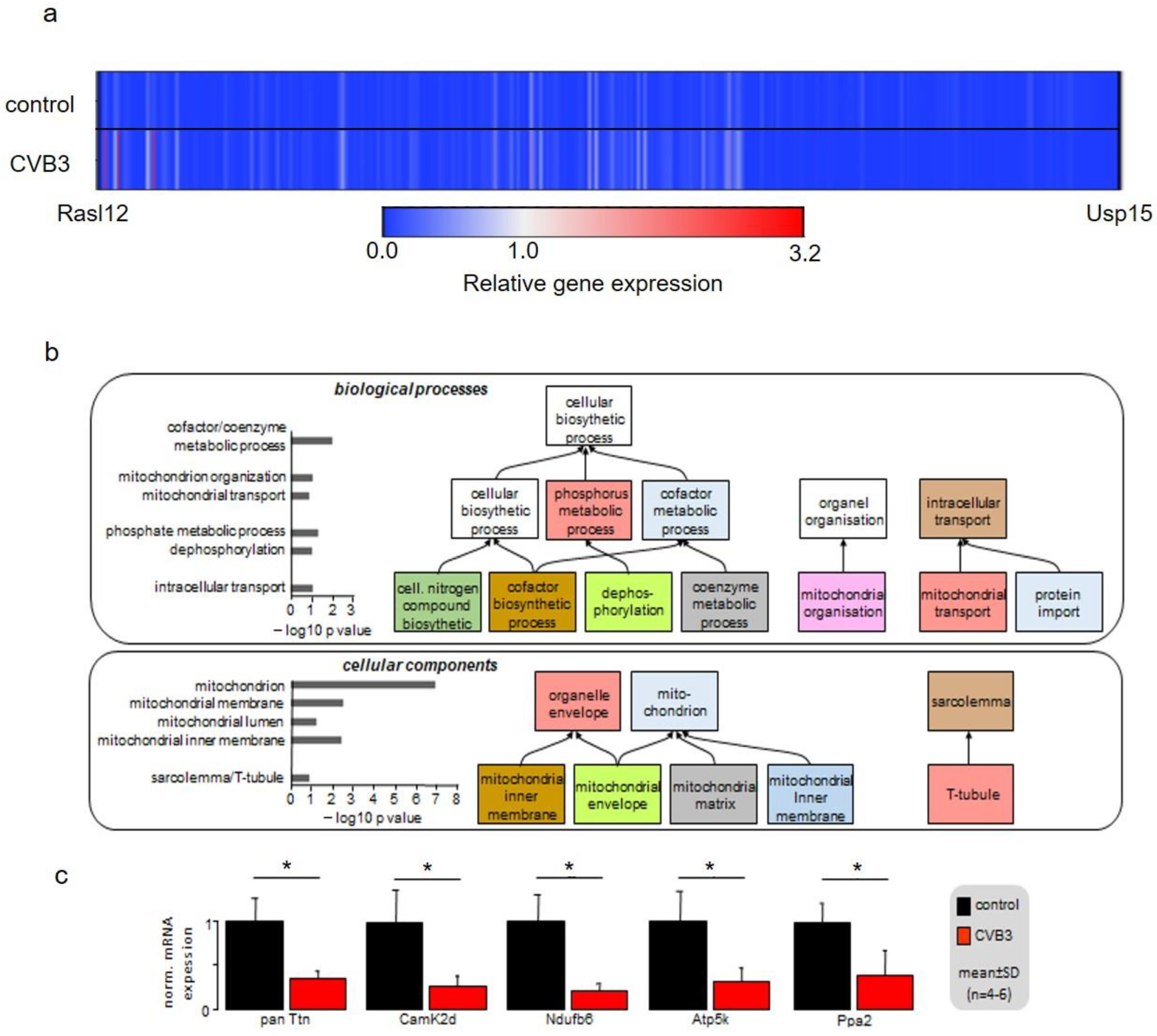
In order to identify the mechanism underlying these phenomena, gene array analyses were conducted. Heatmap illustration shows a clear tendency of transcriptional downregulation in CVB3ΔVP0-expressing cells compared to control cells (
Figure 6a). GO biological process and GO cellular components analyses identified y genes involved in mitochondrial function and structure to be predominantly downregulated (
Figure 6b). Mitochondria are the power plants of the cells including cardiac myocytes. Downregulation of Ndufb6, ATP5K and PPA2, that are key to normal mitochondrial complex I (Ndufb6-NADH dehydrogenase complex) and complex V (PPA2-cytochrome C oxidase complex, F-type ATPase-phosphorylation by ATP5K) function, were confirmed by RT-PCR (
Figure 6c). Further, CamKIδ, which is involved in mitochondrial gene reprogramming, was found downregulated in CVB3ΔVP0 cardiomyocytes (
Figure 6c) [
30]. CamKIδ regulation may represent an important mechanism contributing to mitotoxicity in CVB3ΔVP0 cardiac myocytes, but this still has to be further investigated. Electron microscopy (EM) uncovered a clear reduction of mitochondrial size, strongly supporting effects of CVB3ΔVP0 on mitochondrial structure (
Figure 5).
Recently, it was shown that the mitochondrial respiratory chain has a critical antiviral role in acute CVB3 infection [
31,
32]. Downregulation of energy metabolism may represent a host response effective for acute virus elimination as effective virus replication is highly ATP-dependent [
33,
34,
35]. In the cardiac system, ATP is crucial for two key processes in excitation–contraction coupling, the ATP-driven Ca
2+ reuptake into the sarcoplasmic reticulum and the ATP-driven contraction crossbridge cycle. Reuptake of Ca
2+ is mediated by ATP-driven sarcoplasmic/endoplasmic reticulum calcium ATPase (SERCA). During a contraction crossbridge cycle in myocytes, one ATP is required to form the bridge of myosin to the actin filament allowing for the power stroke and a second ATP to allow for the detachment of myosin. The whole cycle is initiated by binding of Ca
2+ to troponin C on the actin filaments. As mitochondrial ATP synthesis seems to be reduced by the host cell to downregulate viral replication, negative side effects on calcium-contraction coupling and ATP-dependent Ca
2+ uptake into the SR are expected. Thus, the observed features including stress-dependent decompensation seen in mice closely mimic the clinical situation.
In summary, in CVB3 protein-expressing cardiomyocytes, contractile elements and mitochondria are deregulated, which compromises excitation–contraction coupling in late phase action potentials. As a result, cardiac performance is reduced and stress-triggered decompensations with arrhythmic tendency evolve.



 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}