
Alpha1A- and Beta3-Adrenoceptors Interplay in Adipose Multipotent Mesenchymal Stromal Cells: A Novel Mechanism of Obesity-Driven Hypertension
 , , , , , ,
, , , , , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. MSCs Isolation and Culturing
2.2. Immunofluorescent Detection of Alpha1A-Adrenoceptor
2.3. Western-Blotting
2.4. Single-Cell Droplet-Based RNA-Seq Library Preparation and Sequencing
2.5. Collagen Gel Contraction Assay
2.6. MSCs Treatment and Ca2+ Imaging
2.7. CRISPR/Cas9-Mediated Knockout of ADRB2/ADRB3 Genes in ASC52telo Cell Line
2.8. Quantification and Statistical Analysis
2.8.1. Analysis and Quality Control of Single-Cell RNA-Seq Data
2.8.2. Data Representation and Statistical Analysis
3. Results
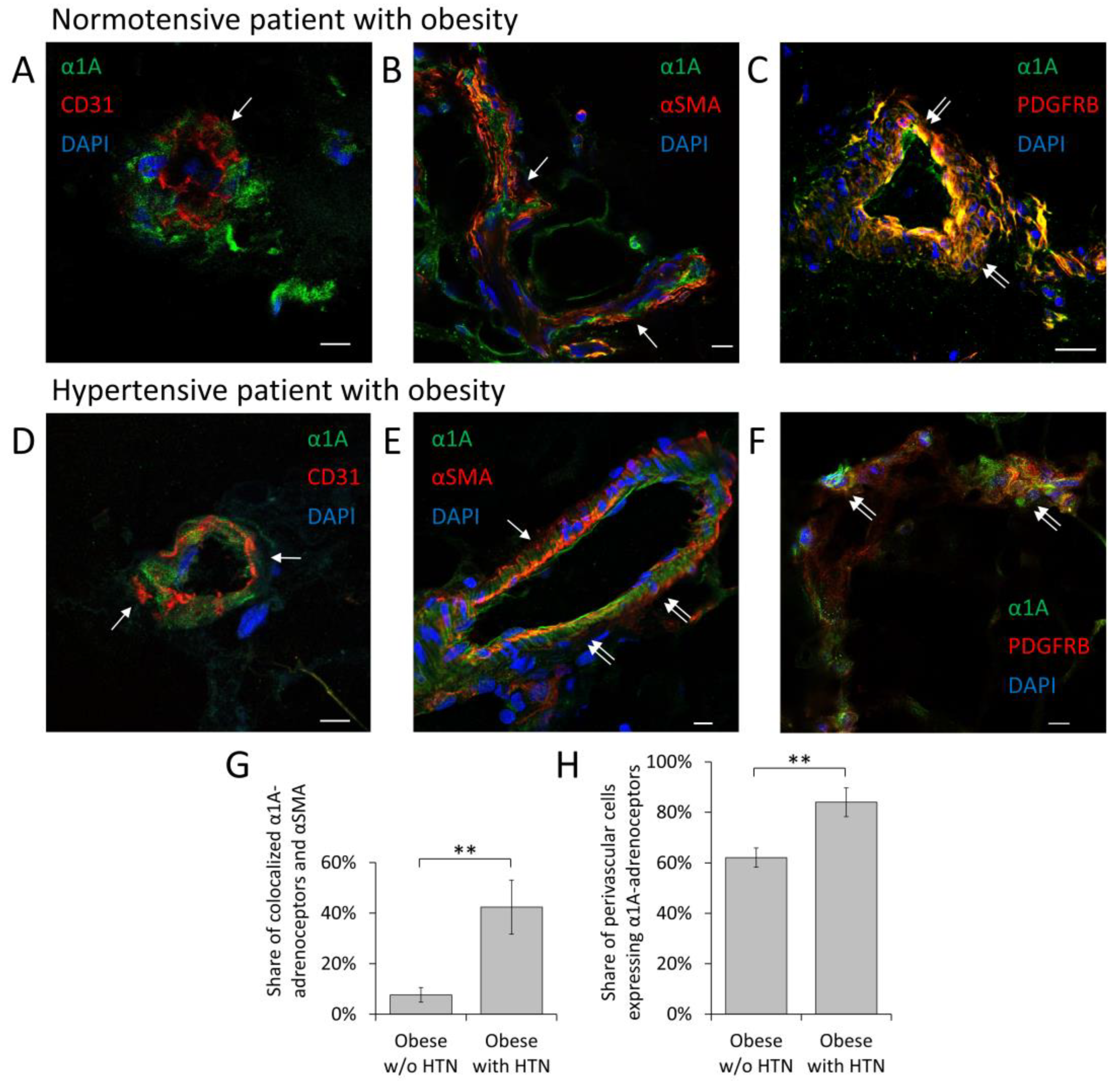
3.1. α1A-Adrenoceptor Is a Predominant Adrenoceptor Isoform Expressed by MSCs in Adipose Tissue Vessels
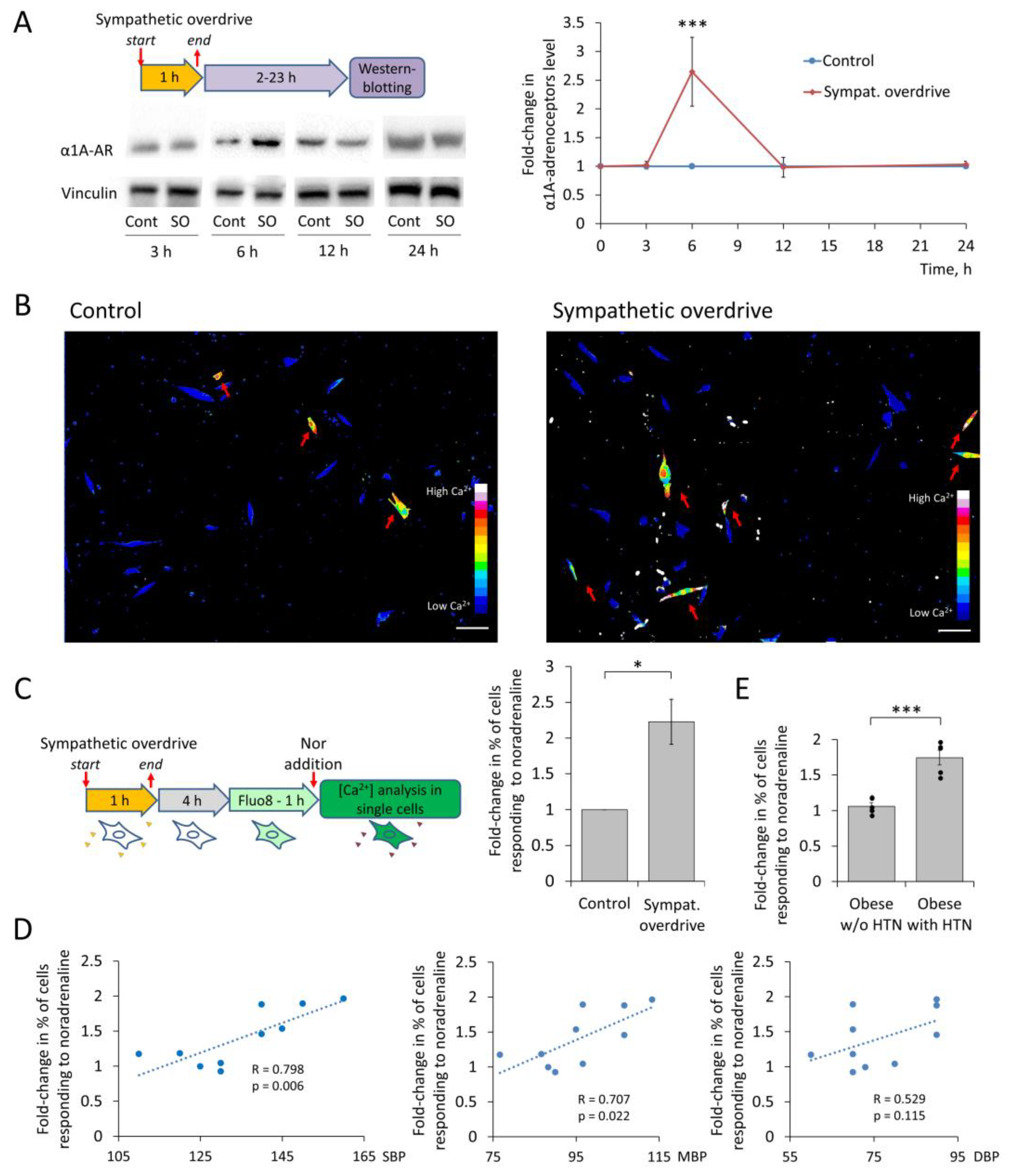
3.2. MSCs Sensitivity to Noradrenaline after Sympathetic Overdrive Correlates with Elevated Blood Pressure in Patients with Obesity
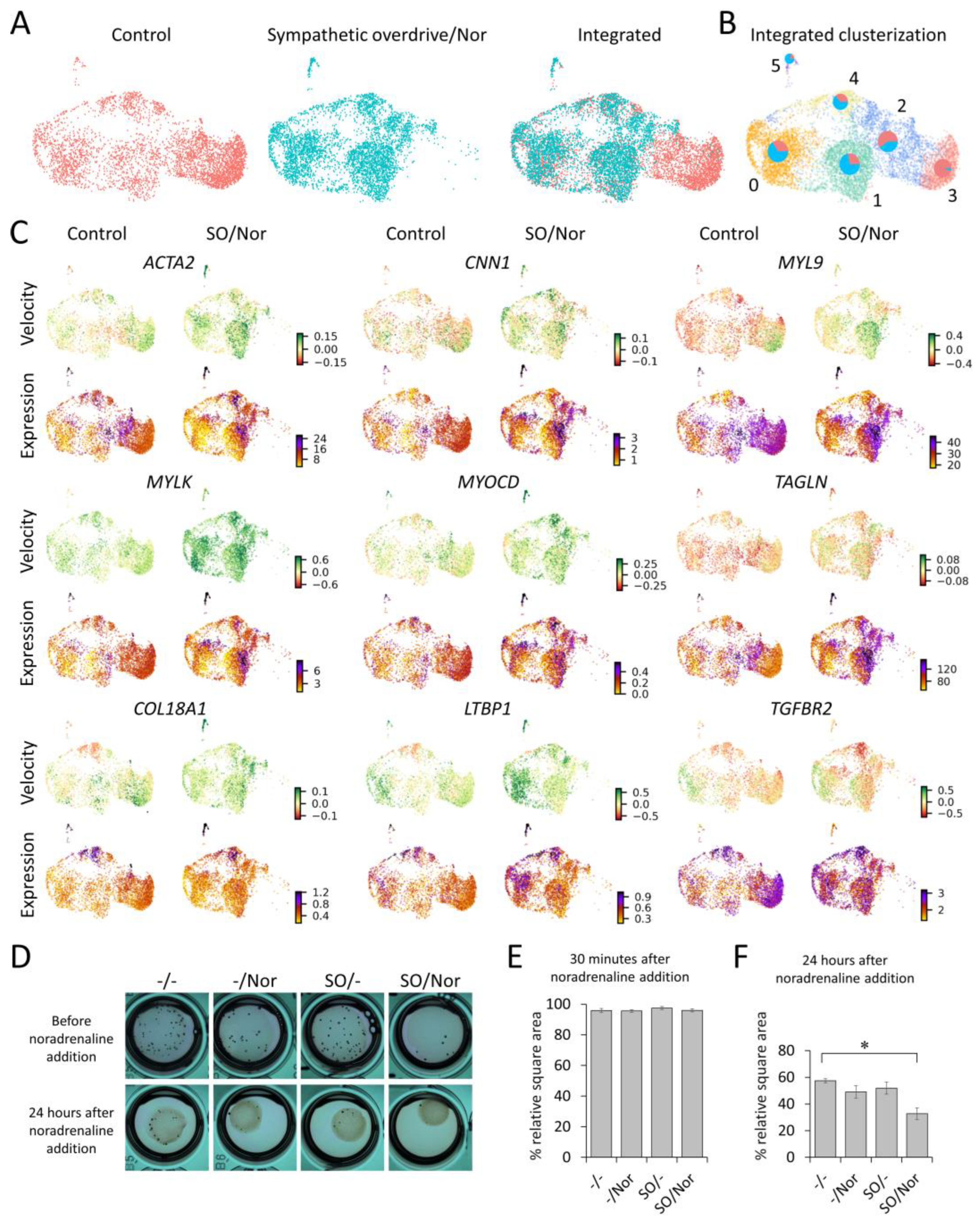
3.3. Single Cell RNA-Seq Reveals MSCs Phenotype Shift towards Contractile Cells
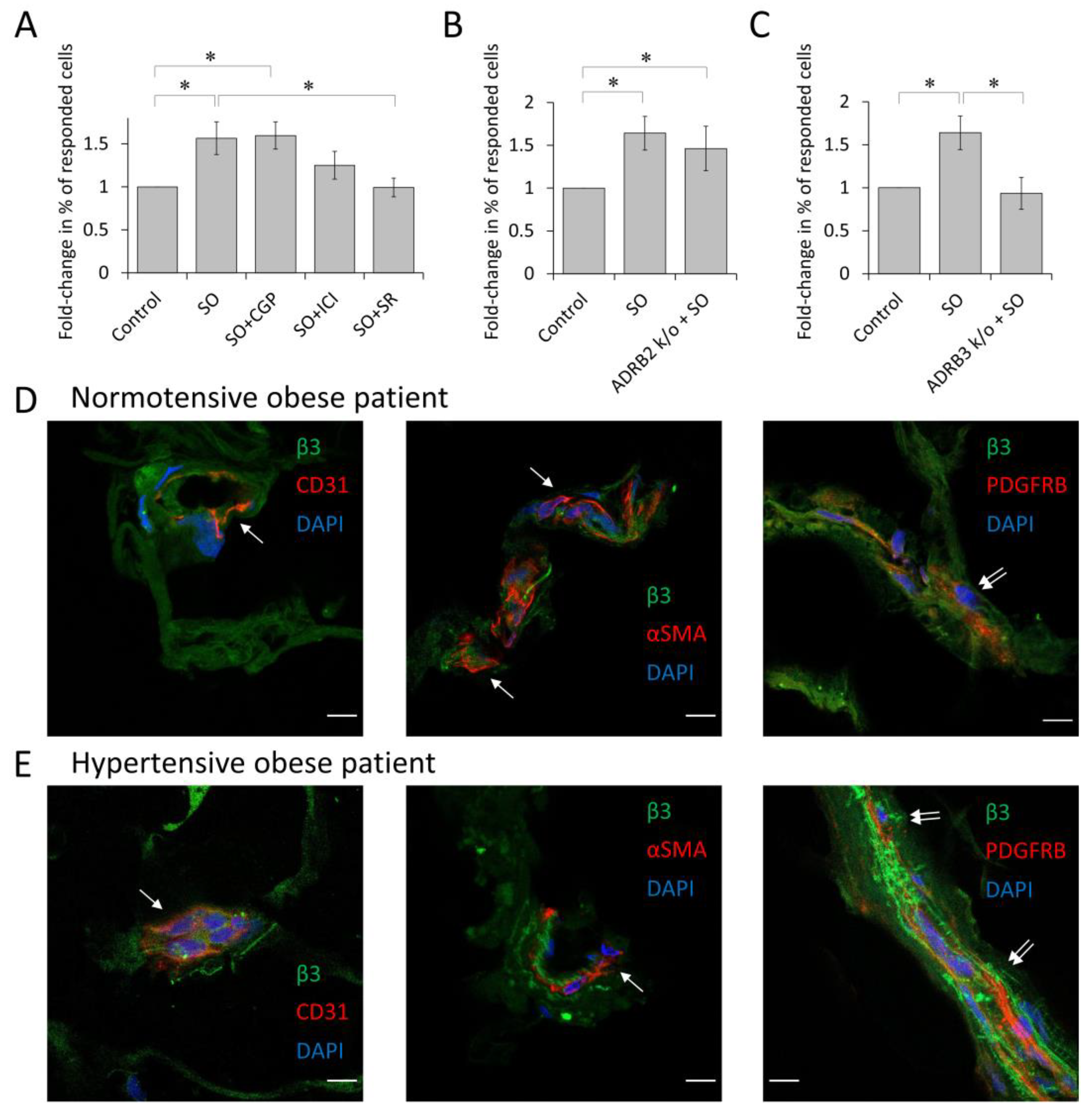
3.4. β3-Adrenoceptor Is Responsible for α1A-Adrenoceptor Elevation upon Sympathetic Overdrive
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Psaty, B.M.; Manolio, T.A.; Smith, N.L.; Heckbert, S.R.; Gottdiener, J.S.; Burke, G.L.; Weissfeld, J.; Enright, P.; Lumley, T.; Powe, N.; et al. Time Trends in High Blood Pressure Control and the Use of Antihypertensive Medications in Older Adults: The Cardiovascular Health Study. Arch. Intern. Med. 2002, 162, 2325–2332. [Google Scholar] [CrossRef]
- Psaty, B.M.; Furberg, C.D.; Kuller, L.H.; Cushman, M.; Savage, P.J.; Levine, D.; O’Leary, D.H.; Bryan, R.N.; Anderson, M.; Lumley, T. Association Between Blood Pressure Level and the Risk of Myocardial Infarction, Stroke, and Total Mortality: The Cardiovascular Health Study. Arch. Intern. Med. 2001, 161, 1183–1192. [Google Scholar] [CrossRef]
- Touyz, R.M.; Alves-Lopes, R.; Rios, F.J.; Camargo, L.L.; Anagnostopoulou, A.; Arner, A.; Montezano, A.C. Vascular smooth muscle contraction in hypertension. Cardiovasc. Res. 2018, 114, 529–539. [Google Scholar] [CrossRef]
- Aroor, A.R.; Jia, G.; Sowers, J.R. Cellular mechanisms underlying obesity-induced arterial stiffness. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 314, R387–R398. [Google Scholar] [CrossRef]
- Rudner, X.L.; Berkowitz, D.E.; Booth, J.V.; Funk, B.L.; Cozart, K.L.; D’Amico, E.B.; El-Moalem, H.; Page, S.O.; Richardson, C.D.; Winters, B.; et al. Subtype Specific Regulation of Human Vascular α1 -Adrenergic Receptors by Vessel Bed and Age. Circulation 1999, 100, 2336–2343. [Google Scholar] [CrossRef] [PubMed]
- Kotchen, T.A. Obesity-Related Hypertension: Epidemiology, Pathophysiology, and Clinical Management. Am. J. Hypertens. 2010, 23, 1170–1178. [Google Scholar] [CrossRef] [PubMed]
- Baer, P.C.; Geiger, H. Adipose-Derived Mesenchymal Stromal/Stem Cells: Tissue Localization, Characterization, and Heterogeneity. Stem Cells Int. 2012, 2012, 812693. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.I.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage Cells from Human Adipose Tissue: Implications for Cell-Based Therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef]
- Kalinina, N.; Kharlampieva, D.; Loguinova, M.; Butenko, I.; Pobeguts, O.; Efimenko, A.; Ageeva, L.; Sharonov, G.; Ischenko, D.; Alekseev, D.; et al. Characterization of secretomes provides evidence for adipose-derived mesenchymal stromal cells subtypes. Stem Cell Res. Ther. 2015, 6, 221. [Google Scholar] [CrossRef]
- Ngo, P.; Ramalingam, P.; Phillips, J.A.; Furuta, G.T. Collagen Gel Contraction Assay. Cell-Cell Interact. Methods Protoc. 2006, 341, 103–109. [Google Scholar] [CrossRef]
- Longo, P.A.; Kavran, J.M.; Kim, M.-S.; Leahy, D.J. Transient Mammalian Cell Transfection with Polyethylenimine (PEI). Methods Enzymol. 2013, 529, 227–240. [Google Scholar] [CrossRef]
- Tyurin-Kuzmin, P.A.; Karagyaur, M.N.; Kulebyakin, K.Y.; Dyikanov, D.T.; Chechekhin, V.I.; Ivanova, A.M.; Skryabina, M.N.; Arbatskiy, M.S.; Sysoeva, V.Y.; Kalinina, N.I.; et al. Functional Heterogeneity of Protein Kinase A Activation in Multipotent Stromal Cells. Int. J. Mol. Sci. 2020, 21, 4442. [Google Scholar] [CrossRef]
- Hao, Y.; Hao, S.; Andersen-Nissen, E.; Mauck, W.M., 3rd; Zheng, S.; Butler, A.; Lee, M.J.; Wilk, A.J.; Darby, C.; Zager, M.; et al. Integrated analysis of multimodal single-cell data. Cell 2021, 184, 3573–3587.e29. [Google Scholar] [CrossRef]
- La Manno, G.; Soldatov, R.; Zeisel, A.; Braun, E.; Hochgerner, H.; Petukhov, V.; Lidschreiber, K.; Kastriti, M.E.; Lönnerberg, P.; Furlan, A.; et al. RNA velocity of single cells. Nature 2018, 560, 494–498. [Google Scholar] [CrossRef]
- Bergen, V.; Lange, M.; Peidli, S.; Wolf, F.A.; Theis, F.J. Generalizing RNA velocity to transient cell states through dynamical modeling. Nat. Biotechnol. 2020, 38, 1408–1414. [Google Scholar] [CrossRef]
- Saelens, W.; Cannoodt, R.; Todorov, H.; Saeys, Y. A comparison of single-cell trajectory inference methods. Nat. Biotechnol. 2019, 37, 547–554. [Google Scholar] [CrossRef]
- Wolf, F.A.; Hamey, F.K.; Plass, M.; Solana, J.; Dahlin, J.S.; Göttgens, B.; Rajewsky, N.; Simon, L.; Theis, F.J. PAGA: Graph abstraction reconciles clustering with trajectory inference through a topology preserving map of single cells. Genome Biol. 2019, 20, 59. [Google Scholar] [CrossRef]
- Hildreth, A.D.; Ma, F.; Wong, Y.Y.; Sun, R.; Pellegrini, M.; O’Sullivan, T.E. Single-cell sequencing of human white adipose tissue identifies new cell states in health and obesity. Nat. Immunol. 2021, 22, 639–653. [Google Scholar] [CrossRef]
- Muhl, L.; Genové, G.; Leptidis, S.; Liu, J.; He, L.; Mocci, G.; Sun, Y.; Gustafsson, S.; Buyandelger, B.; Chivukula, I.V.; et al. Single-cell analysis uncovers fibroblast heterogeneity and criteria for fibroblast and mural cell identification and discrimination. Nat. Commun. 2020, 11, 3953. [Google Scholar] [CrossRef]
- Tyurin-Kuzmin, P.A.; Fadeeva, J.I.; Kanareikina, M.A.; Kalinina, N.I.; Sysoeva, V.Y.; Dyikanov, D.T.; Stambolsky, D.V.; Tkachuk, V.A. Activation of β-adrenergic receptors is required for elevated α1A-adrenoreceptors expression and signaling in mesenchymal stromal cells. Sci. Rep. 2016, 6, 32835. [Google Scholar] [CrossRef]
- Yoshimura, K.; Suga, H.; Eto, H. Adipose-derived stem/progenitor cells: Roles in adipose tissue remodeling and potential use for soft tissue augmentation. Regen. Med. 2009, 4, 265–273. [Google Scholar] [CrossRef]
- Naji, A.; Eitoku, M.; Favier, B.; Deschaseaux, F.; Rouas-Freiss, N.; Suganuma, N. Biological functions of mesenchymal stem cells and clinical implications. Cell. Mol. Life Sci. 2019, 76, 3323–3348. [Google Scholar] [CrossRef] [PubMed]
- Allen, B.G.; Walsh, M.P. The biochemical basis of the regulation of smooth-muscle contraction. Trends Biochem. Sci. 1994, 19, 362–368. [Google Scholar] [CrossRef]
- Kalil, G.Z.; Haynes, W.G. Sympathetic nervous system in obesity-related hypertension: Mechanisms and clinical implications. Hypertens. Res. 2012, 35, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Grassi, G.; Dell’Oro, R.; Facchini, A.; Quarti Trevano, F.; Bolla, G.B.; Mancia, G. Effect of central and peripheral body fat distribution on sympathetic and baroreflex function in obese normotensives. J. Hypertens. 2004, 22, 2363–2369. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.; Zhu, Y.; Sun, Z.; Trzeciakowski, J.P.; Gansner, M.; Depre, C.; Resuello, R.R.; Natividad, F.F.; Hunter, W.C.; Genin, G.M.; et al. Short Communication: Vascular Smooth Muscle Cell Stiffness As a Mechanism for Increased Aortic Stiffness With Aging. Circ. Res. 2010, 107, 615–619. [Google Scholar] [CrossRef]
- Sun, K.; Tordjman, J.; Clement, K.; Scherer, P.E. Fibrosis and Adipose Tissue Dysfunction. Cell Metab. 2013, 18, 470–477. [Google Scholar] [CrossRef]
- Li, G.; Cao, Z.; Wu, X.-W.; Wu, H.-K.; Ma, Y.; Wu, B.; Wang, W.-Q.; Cheng, J.; Zhou, Z.-H.; Tu, Y.-C. Autoantibodies against AT1 and α1-adrenergic receptors predict arterial stiffness progression in normotensive subjects over a 5-year period. Clin. Sci. 2017, 131, 2947–2957. [Google Scholar] [CrossRef]
- Méndez-Ferrer, S.; Michurina, T.V.; Ferraro, F.; Mazloom, A.R.; MacArthur, B.D.; Lira, S.A.; Scadden, D.T.; Ma’Ayan, A.; Enikolopov, G.N.; Frenette, P.S. Mesenchymal and haematopoietic stem cells form a unique bone marrow niche. Nature 2010, 466, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Pausova, Z.; Jomphe, M.; Houde, L.; Vézina, H.; Orlov, S.N.; Gossard, F.; Gaudet, D.; Tremblay, J.; Kotchen, T.A.; Cowley, A.W.; et al. A Genealogical Study of Essential Hypertension with and without Obesity in French Canadians. Obes. Res. 2002, 10, 463–470. [Google Scholar] [CrossRef] [Green Version]
- Dessy, C.; Balligand, J.-L. Beta3-Adrenergic Receptors in Cardiac and Vascular Tissues: Emerging Concepts and Therapeutic Perspectives. Adv. Pharmacol. 2010, 59, 135–163. [Google Scholar] [CrossRef] [PubMed]
- Michel, L.Y.M.; Farah, C.; Balligand, J.-L. The Beta3 Adrenergic Receptor in Healthy and Pathological Cardiovascular Tissues. Cells 2020, 9, 2584. [Google Scholar] [CrossRef] [PubMed]
- Wofford, M.R.; Anderson, D.C.; Brown, A.; Jones, D.W.; Miller, M.E.; Hall, J.E. Antihypertensive Effect of α-and β-Adrenergic Blockade in Obese and Lean Hypertensive Patients. Am. J. Hypertens. 2001, 14, 694–698. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence | Amplicon Length, bp | Tmelting, °C |
|---|---|---|---|
| hADRB2-test-f | GCAACTTCTGGTGCGAGTTT | 415 | 59.5 |
| hADRB2-test-r | AAGCGGCCCTCAGATTTGTC | ||
| hADRB3-test2-f | GCAGTAGATGAGCGGGTTGAA | 830 | 60 |
| hADRB3-test2-r | ACGTGTTCGTGACTTCGCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chechekhin, V.; Ivanova, A.; Kulebyakin, K.; Sysoeva, V.; Naida, D.; Arbatsky, M.; Basalova, N.; Karagyaur, M.; Skryabina, M.; Efimenko, A.; et al. Alpha1A- and Beta3-Adrenoceptors Interplay in Adipose Multipotent Mesenchymal Stromal Cells: A Novel Mechanism of Obesity-Driven Hypertension. Cells 2023, 12, 585. https://doi.org/10.3390/cells12040585
Chechekhin V, Ivanova A, Kulebyakin K, Sysoeva V, Naida D, Arbatsky M, Basalova N, Karagyaur M, Skryabina M, Efimenko A, et al. Alpha1A- and Beta3-Adrenoceptors Interplay in Adipose Multipotent Mesenchymal Stromal Cells: A Novel Mechanism of Obesity-Driven Hypertension. Cells. 2023; 12(4):585. https://doi.org/10.3390/cells12040585
Chicago/Turabian StyleChechekhin, Vadim, Anastasia Ivanova, Konstantin Kulebyakin, Veronika Sysoeva, Daria Naida, Mikhail Arbatsky, Nataliya Basalova, Maxim Karagyaur, Mariya Skryabina, Anastasia Efimenko, and et al. 2023. "Alpha1A- and Beta3-Adrenoceptors Interplay in Adipose Multipotent Mesenchymal Stromal Cells: A Novel Mechanism of Obesity-Driven Hypertension" Cells 12, no. 4: 585. https://doi.org/10.3390/cells12040585