
High-Throughput Quantitative Screening of Glucose-Stimulated Insulin Secretion and Insulin Content Using Automated MALDI-TOF Mass Spectrometry
 ,
,  , , ,
, , ,  and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Treatments
2.2. Automated Glucose-Stimulated Insulin Secretion (GSIS) in 384-Well Format
2.3. Automated siRNA Reverse Transfection in 384-Well Format
2.4. Automated Incubation with Chemical Compounds
2.5. Automated MALDI-TOF Mass Spectrometry Analysis
2.6. Measurement of Transfection Efficiency by Flow Cytometry
2.7. Immunofluorescence
2.8. RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.9. Immunoblotting Experiments
2.10. Statistical Analysis
3. Results
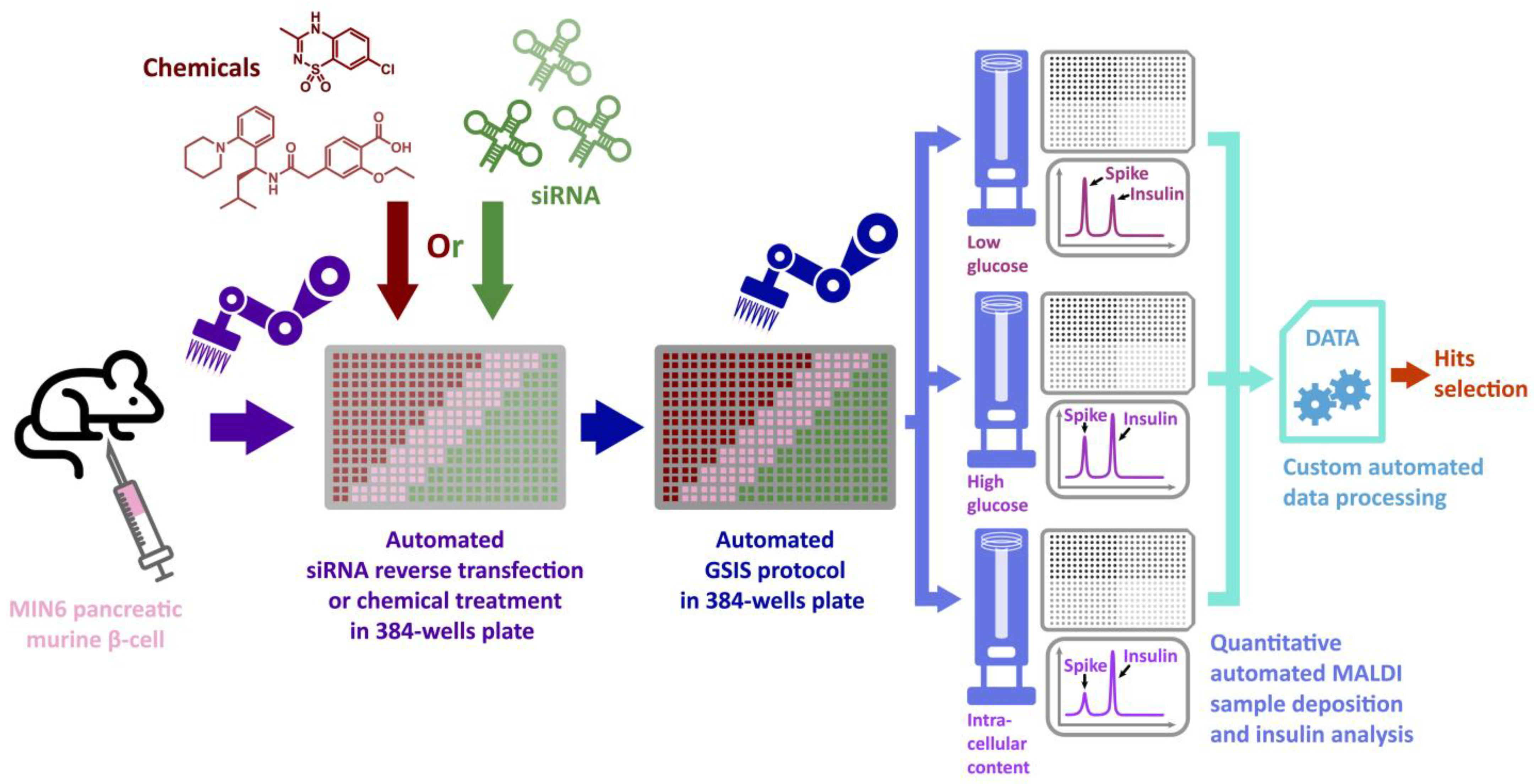
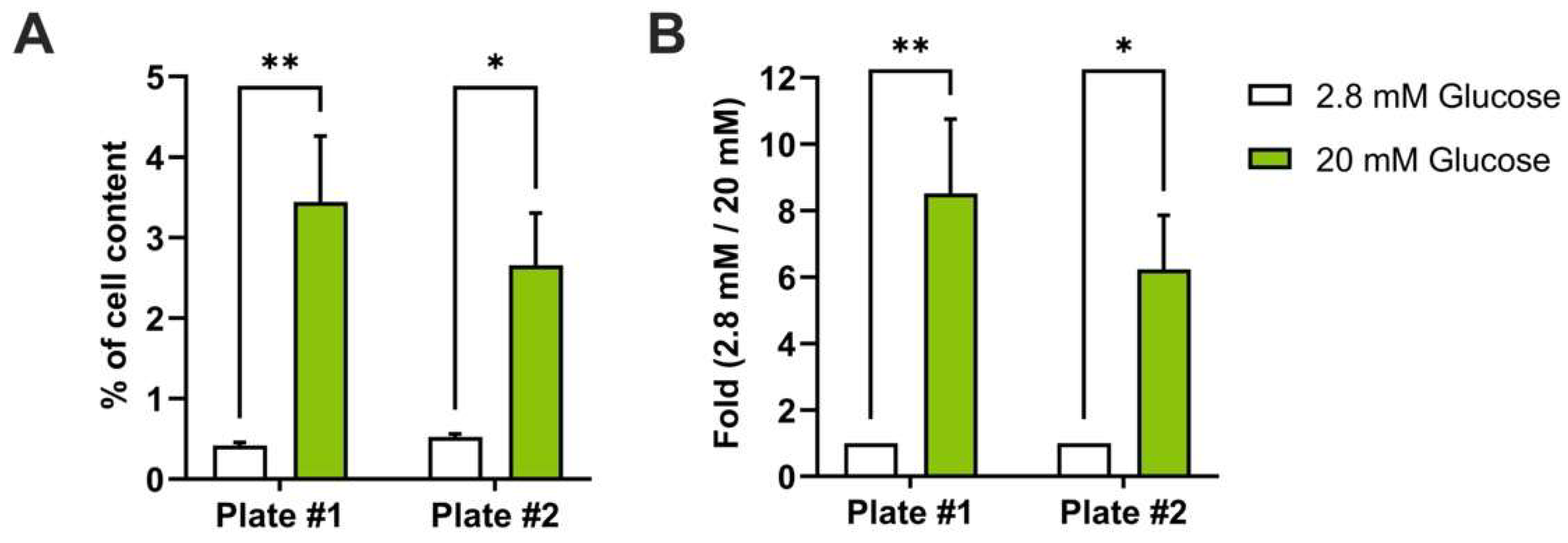
3.1. Automation of Glucose-Stimulated Insulin Secretion (GSIS) Assay
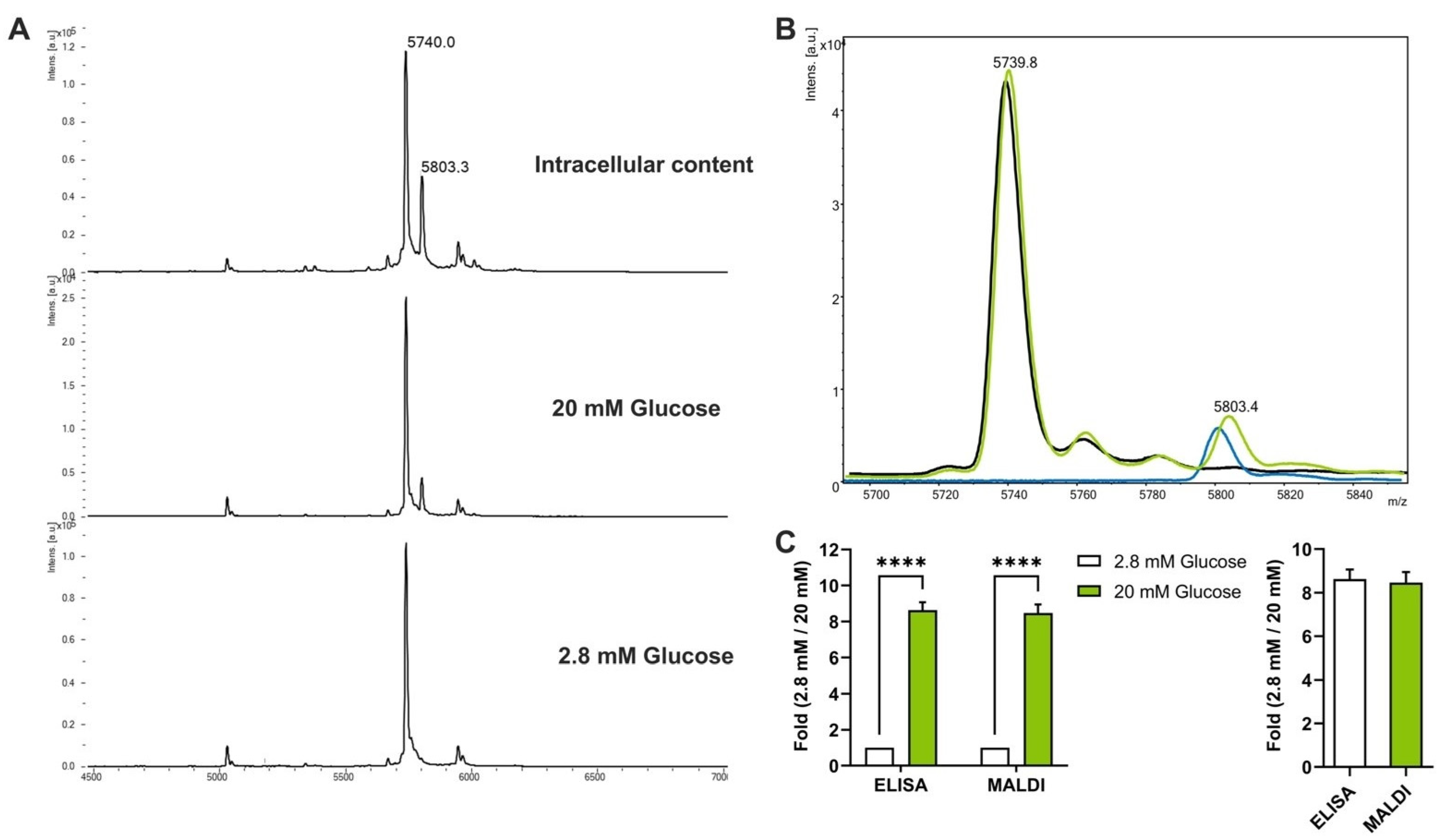
3.2. Automated Analysis of GSIS in 384-Well Plates through MALDI-TOF Mass Spectrometry
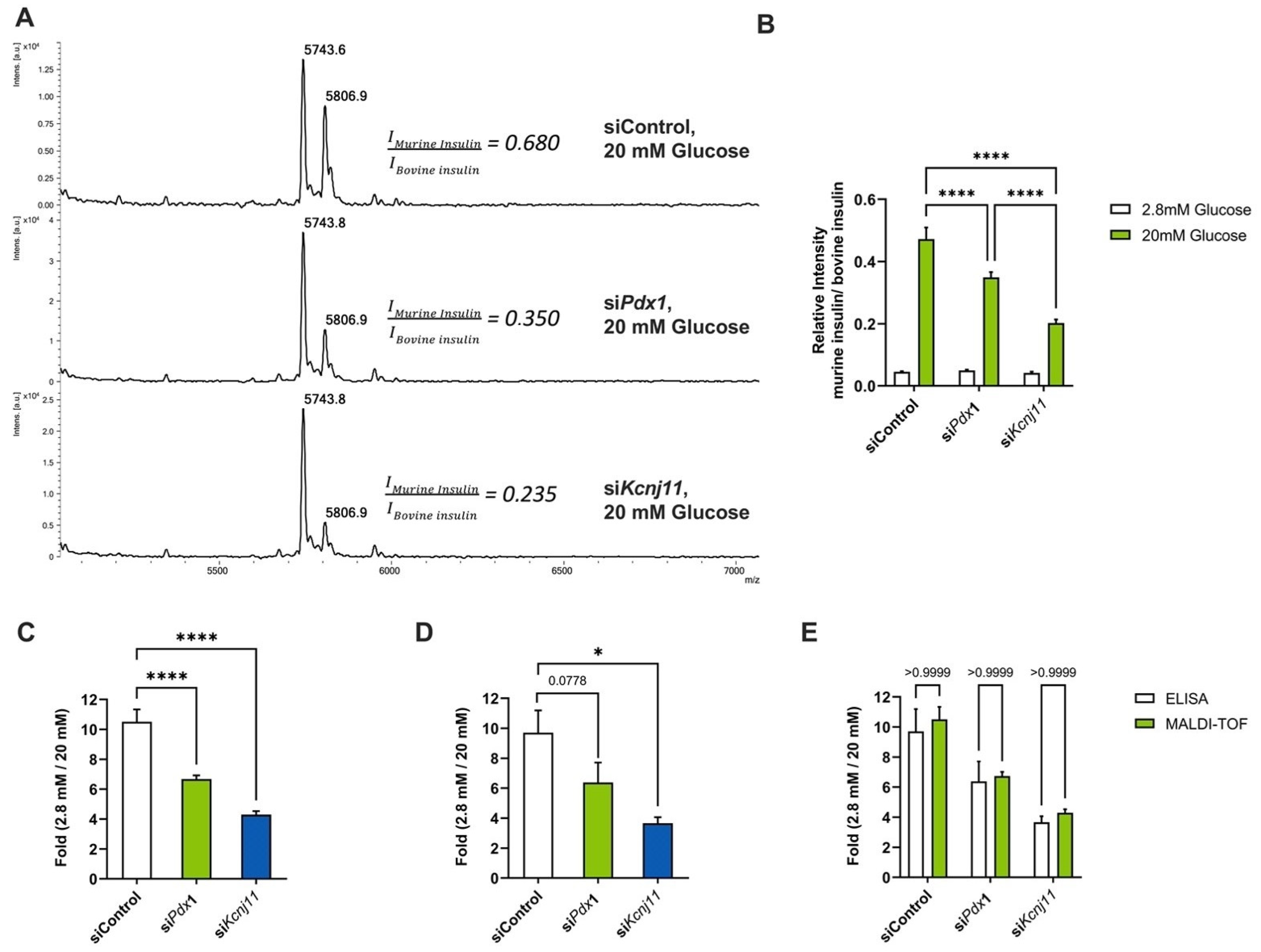
3.3. Automated siRNA Transfection Combined with GSIS
3.4. Automated Chemical Treatment Combined with GSIS
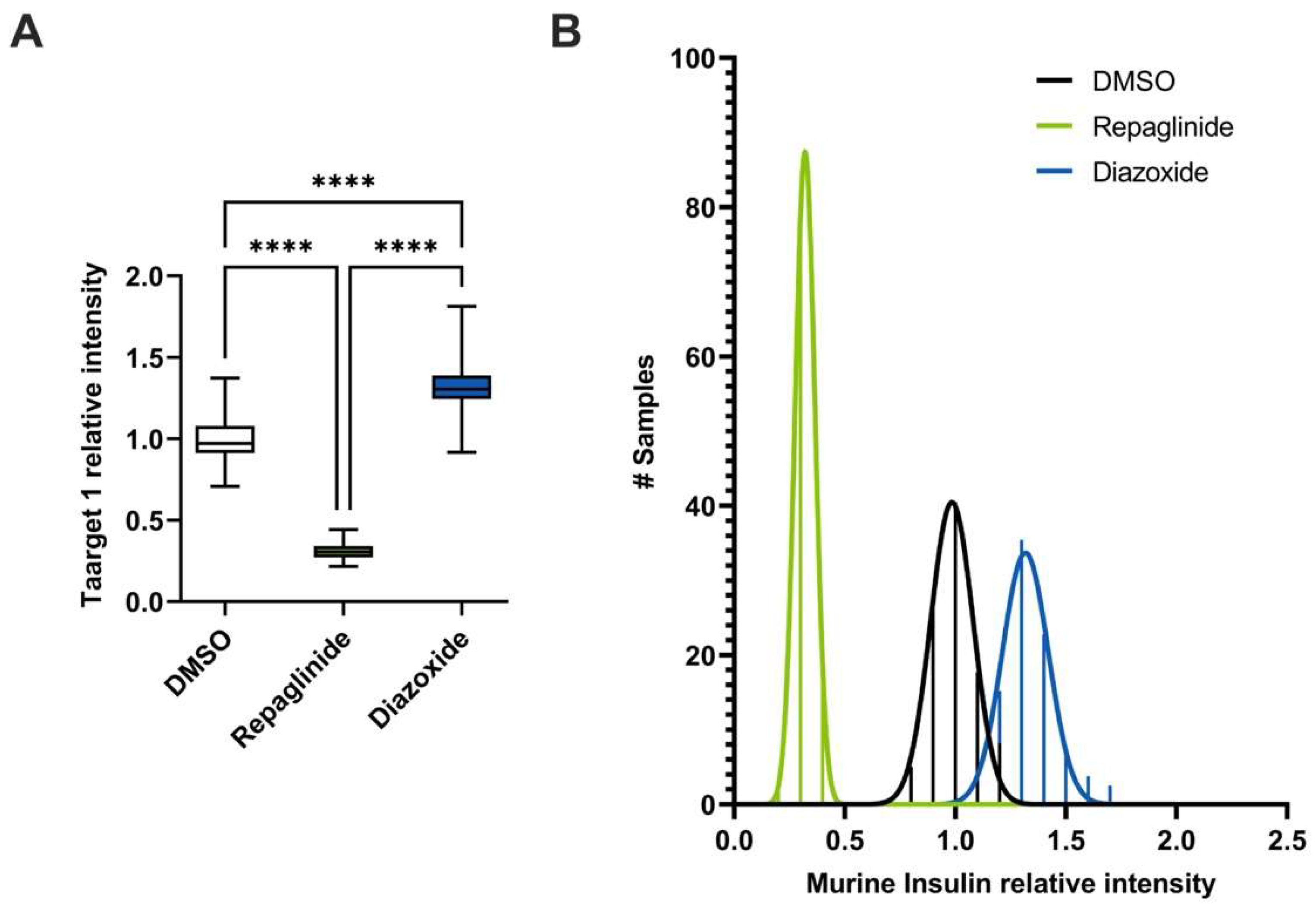
3.5. Automated Library Screening to Measure Insulin Intracellular Content after GSIS
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lin, X.; Xu, Y.; Pan, X.; Xu, J.; Ding, Y.; Sun, X.; Song, X.; Ren, Y.; Shan, P.-F. Global, Regional, and National Burden and Trend of Diabetes in 195 Countries and Territories: An Analysis from 1990 to 2025. Sci. Rep. 2020, 10, 14790. [Google Scholar] [CrossRef] [PubMed]
- Artasensi, A.; Pedretti, A.; Vistoli, G.; Fumagalli, L. Type 2 Diabetes Mellitus: A Review of Multi-Target Drugs. Molecules 2020, 25, 1987. [Google Scholar] [CrossRef] [PubMed]
- Nisal, K.; Kela, R.; Khunti, K.; Davies, M.J. Comparison of Efficacy between Incretin-Based Therapies for Type 2 Diabetes Mellitus. BMC Med. 2012, 10, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguayo-Mazzucato, C. Functional Changes in Beta Cells during Ageing and Senescence. Diabetologia 2020, 63, 2022–2029. [Google Scholar] [CrossRef] [PubMed]
- Heyduk, E.; Moxley, M.M.; Salvatori, A.; Corbett, J.A.; Heyduk, T. Homogeneous Insulin and C-Peptide Sensors for Rapid Assessment of Insulin and C-Peptide Secretion by the Islets. Diabetes 2010, 59, 2360–2365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalwat, M.A.; Wichaidit, C.; Nava Garcia, A.Y.; McCoy, M.K.; McGlynn, K.; Hwang, I.H.; MacMillan, J.B.; Posner, B.A.; Cobb, M.H. Insulin Promoter-Driven Gaussia Luciferase-Based Insulin Secretion Biosensor Assay for Discovery of β-Cell Glucose-Sensing Pathways. ACS Sens. 2016, 1, 1208–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burns, S.M.; Vetere, A.; Walpita, D.; Dančík, V.; Khodier, C.; Perez, J.; Clemons, P.A.; Wagner, B.K.; Altshuler, D. High-Throughput Luminescent Reporter of Insulin Secretion for Discovering Regulators of Pancreatic Beta-Cell Function. Cell Metab. 2015, 21, 126–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pappalardo, Z.; Chopra, D.G.; Hennings, T.G.; Richards, H.; Choe, J.; Yang, K.; Baeyens, L.; Ang, K.; Chen, S.; Arkin, M.; et al. A Whole-Genome RNA Interference Screen Reveals a Role for Spry2 in Insulin Transcription and the Unfolded Protein Response. Diabetes 2017, 66, 1703–1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalwat, M.A. High-Throughput Screening for Insulin Secretion Modulators. In Exocytosis and Endocytosis: Methods and Protocols; Niedergang, F., Vitale, N., Gasman, S., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2021; pp. 131–138. ISBN 978-1-07-161044-2. [Google Scholar]
- Bourouh, C.; Courty, E.; Rolland, L.; Pasquetti, G.; Gromada, X.; Rabhi, N.; Carney, C.; Moreno, M.; Boutry, R.; Caron, E.; et al. The Transcription Factor E2F1 Controls the GLP-1 Receptor Pathway in Pancreatic β Cells. Cell Rep. 2022, 40, 111170. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Shang, J.; Feng, Y.; Thompson, C.M.; Horwitz, S.; Thompson, J.R.; Macintyre, E.D.; Thornberry, N.A.; Chapman, K.; Zhou, Y.-P.; et al. Identification of Glucose-Dependent Insulin Secretion Targets in Pancreatic β Cells by Combining Defined-Mechanism Compound Library Screening and SiRNA Gene Silencing. J. Biomol. Screen 2008, 13, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Aamodt, K.I.; Aramandla, R.; Brown, J.J.; Fiaschi-Taesch, N.; Wang, P.; Stewart, A.F.; Brissova, M.; Powers, A.C. Development of a Reliable Automated Screening System to Identify Small Molecules and Biologics That Promote Human β-Cell Regeneration. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E859–E868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szczerbinska, I.; Tessitore, A.; Hansson, L.K.; Agrawal, A.; Lopez, A.R.; Helenius, M.; Malinowski, A.R.; Gilboa, B.; Ruby, M.A.; Gupta, R.; et al. Large-Scale Functional Genomics Screen to Identify Modulators of Human β-Cell Insulin Secretion. Biomedicines 2022, 10, 103. [Google Scholar] [CrossRef] [PubMed]
- Bevacqua, R.J.; Dai, X.; Lam, J.Y.; Gu, X.; Friedlander, M.S.H.; Tellez, K.; Miguel-Escalada, I.; Bonàs-Guarch, S.; Atla, G.; Zhao, W.; et al. CRISPR-Based Genome Editing in Primary Human Pancreatic Islet Cells. Nat. Commun. 2021, 12, 2397. [Google Scholar] [CrossRef] [PubMed]
- Grotz, A.K.; Abaitua, F.; Navarro-Guerrero, E.; Hastoy, B.; Ebner, D.; Gloyn, A.L. A CRISPR/Cas9 Genome Editing Pipeline in the EndoC-ΒH1 Cell Line to Study Genes Implicated in Beta Cell Function. Wellcome Open Res. 2020, 4, 150. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delannoy, C.P.; Heuson, E.; Herledan, A.; Oger, F.; Thiroux, B.; Chevalier, M.; Gromada, X.; Rolland, L.; Froguel, P.; Deprez, B.; et al. High-Throughput Quantitative Screening of Glucose-Stimulated Insulin Secretion and Insulin Content Using Automated MALDI-TOF Mass Spectrometry. Cells 2023, 12, 849. https://doi.org/10.3390/cells12060849
Delannoy CP, Heuson E, Herledan A, Oger F, Thiroux B, Chevalier M, Gromada X, Rolland L, Froguel P, Deprez B, et al. High-Throughput Quantitative Screening of Glucose-Stimulated Insulin Secretion and Insulin Content Using Automated MALDI-TOF Mass Spectrometry. Cells. 2023; 12(6):849. https://doi.org/10.3390/cells12060849
Chicago/Turabian StyleDelannoy, Clément Philippe, Egon Heuson, Adrien Herledan, Frederik Oger, Bryan Thiroux, Mickaël Chevalier, Xavier Gromada, Laure Rolland, Philippe Froguel, Benoit Deprez, and et al. 2023. "High-Throughput Quantitative Screening of Glucose-Stimulated Insulin Secretion and Insulin Content Using Automated MALDI-TOF Mass Spectrometry" Cells 12, no. 6: 849. https://doi.org/10.3390/cells12060849