
Cerebral Malaria Model Applying Human Brain Organoids
 ,
,  , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Malaria Parasite Culture
2.2. Endothelial Cell Line-Human Brain Microvascular Endothelial Cells
2.3. Flow Cytometry of HBMEC Cells
2.4. RNA Extraction of HBMEC Cells
2.5. RNA-Seq Analysis of P. falciparum Activated HBMEC Cells
2.6. HBMEC-P. falciparum Activated Secretome Analysis
2.7. iPSCs Culture and Human Brain Organoid Differentiation Media
2.8. Human Brain Organoids Stimulation with HBMEC Secretome
2.9. RNA Extraction of Human Brain Organoids
2.10. RNA-Seq Analysis of Human Brain Organoids
2.11. Human Brain Organoids Cryosection
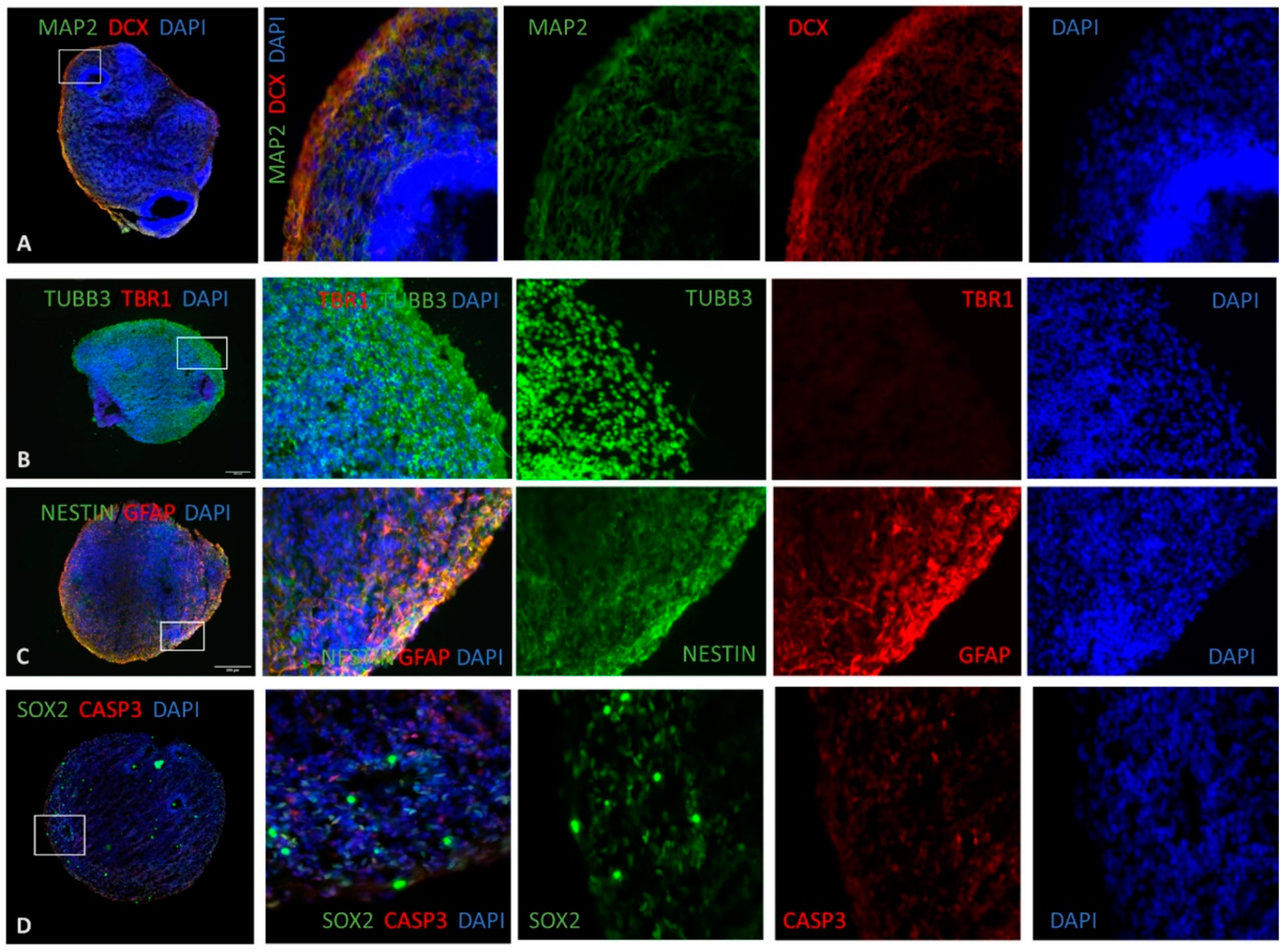
2.12. Immunofluorescence of Human Brain Organoids
3. Results
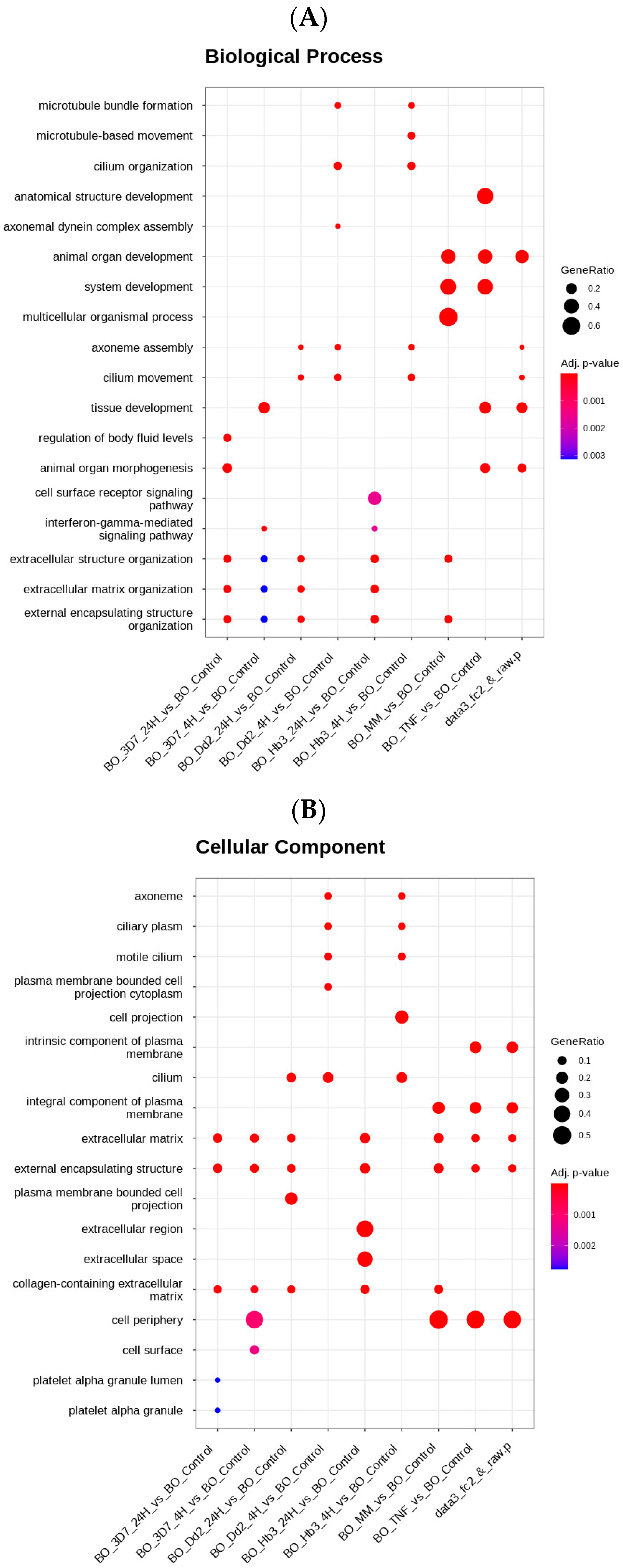
3.1. Transcriptomic Evaluation of Novel Receptor Genes Activated by P. falciparum
3.2. Receptors Expression of HBMEC by P. falciparum Strains
3.3. Impact of P. falciparum Activation in HBMEC Secretome
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- World Malaria Report 2022. Available online: https://www.who.int/publications/i/item/9789240064898 (accessed on 16 January 2023).
- Schiess, N.; Villabona-Rueda, A.; Cottier, K.E.; Huether, K.; Chipeta, J.; Stins, M.F. Pathophysiology and neurologic sequelae of cerebral malaria. Malar. J. 2020, 19, 266. [Google Scholar] [CrossRef]
- Sturtzel, C. Endothelial Cells. Adv. Exp. Med. Biol. 2017, 1003, 71–91. [Google Scholar] [PubMed]
- Nagyőszi, P.; Wilhelm, I.; Farkas, A.E.; Fazakas, C.; Dung, N.T.K.; Haskó, J.; Krizbai, I.A. Expression and regulation of toll-like receptors in cerebral endothelial cells. Neurochem. Int. 2010, 57, 556–564. [Google Scholar] [CrossRef] [PubMed]
- Nagyőszi, P.; Nyúl-Tóth, Á.; Fazakas, C.; Wilhelm, I.; Kozma, M.; Molnár, J.; Haskó, J.; Krizbai, I.A. Regulation of NOD-like receptors and inflammasome activation in cerebral endothelial cells. J. Neurochem. 2015, 135, 551–564. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, L.A.J.; Golenbock, D.; Bowie, A.G. The history of Toll-like receptors—Redefining innate immunity. Nat. Rev. Immunol. 2013, 13, 453–460. [Google Scholar] [CrossRef]
- Ouma, B.J.; Ssenkusu, J.M.; Shabani, E.; Datta, D.; Opoka, R.O.; Idro, R.; Bangirana, P.; Park, G.; Joloba, M.L.; Kain, K.C.; et al. Endothelial Activation, Acute Kidney Injury, and Cognitive Impairment in Pediatric Severe Malaria. Crit. Care Med. 2020, 48, e734–e743. [Google Scholar] [CrossRef]
- Cunningham, D.A.; Lin, J.-W.; Brugat, T.; Jarra, W.; Tumwine, I.; Kushinga, G.; Ramesar, J.; Franke-Fayard, B.; Langhorne, J. ICAM-1 is a key receptor mediating cytoadherence and pathology in the Plasmodium chabaudi malaria model. Malar. J. 2017, 16, 185. [Google Scholar] [CrossRef] [Green Version]
- Storm, J.; Jespersen, J.S.; Seydel, K.B.; Szestak, T.; Mbewe, M.; Chisala, N.V.; Phula, P.; Wang, C.W.; Taylor, T.E.; Moxon, C.; et al. Cerebral malaria is associated with differential cytoadherence to brain endothelial cells. EMBO Mol. Med. 2019, 11, e9164. [Google Scholar] [CrossRef]
- Ndam, N.T.; Moussiliou, A.; Lavstsen, T.; Kamaliddin, C.; Jensen, A.T.R.; Mama, A.; Tahar, R.; Wang, C.; Jespersen, J.S.; Alao, J.M.; et al. Parasites Causing Cerebral Falciparum Malaria Bind Multiple Endothelial Receptors and Express EPCR and ICAM-1-Binding PfEMP1. J. Infect. Dis. 2017, 215, 1918–1925. [Google Scholar] [CrossRef] [Green Version]
- Fleckenstein, H.; Portugal, S. Binding brain better—Matching var genes and endothelial receptors. EMBO Mol. Med. 2019, 11, e10137. [Google Scholar] [CrossRef]
- Kumar, S.; Trivedi, V. Extracellular methemoglobin promotes cyto-adherence of uninfected RBC to endothelial cells: Insight into cerebral malaria pathology. J. Cell. Biochem. 2019, 120, 11140–11149. [Google Scholar] [CrossRef]
- Viebig, N.K.; Wulbrand, U.; Förster, R.; Andrews, K.T.; Lanzer, M.; Knolle, P.A. Direct Activation of Human Endothelial Cells by Plasmodium falciparum-Infected Erythrocytes. Infect. Immun. 2005, 73, 3271–3277. [Google Scholar] [CrossRef] [Green Version]
- Utter, C.; Serrano, A.E.; Glod, J.W.; Leibowitz, M.J. Focus: Infectious Diseases: Association of Plasmodium falciparum with Human Endothelial Cells in vitro. Yale J. Biol. Med. 2017, 90, 183. [Google Scholar]
- Avril, M.; Bernabeu, M.; Benjamin, M.; Brazier, A.J.; Smith, J.D. Interaction between Endothelial Protein C Receptor and Intercellular Adhesion Molecule 1 to Mediate Binding of Plasmodium falciparum-Infected Erythrocytes to Endothelial Cells. mBio 2016, 7, e00615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, L.; Lavstsen, T.; Berger, S.S.; Wang, C.W.; Petersen, J.E.V.; Avril, M.; Brazier, A.J.; Freeth, J.; Jespersen, J.S.; Nielsen, M.A.; et al. Severe malaria is associated with parasite binding to endothelial protein C receptor. Nature 2013, 498, 502–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernabeu, M.; Smith, J.D. EPCR and Malaria Severity: The Center of a Perfect Storm. Trends Parasitol. 2017, 33, 295–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wassmer, S.C.; Taylor, T.; MacLennan, C.A.; Kanjala, M.; Mukaka, M.; Molyneux, M.E.; Grau, G.E. Platelet-Induced Clumping of Plasmodium falciparum—Infected Erythrocytes from Malawian Patients with Cerebral Malaria—Possible Modulation In Vivo by Thrombocytopenia. J. Infect. Dis. 2008, 197, 72–78. [Google Scholar] [CrossRef] [Green Version]
- White, N.J.; Turner, G.D.H.; Day, N.P.J.; Dondorp, A.M. Lethal Malaria: Marchiafava and Bignami Were Right. J. Infect. Dis. 2013, 208, 192–198. [Google Scholar] [CrossRef]
- Storm, J.; Craig, A.G. Pathogenesis of cerebral malaria—Inflammation and cytoadherence. Front. Cell. Infect. Microbiol. 2014, 4, 100. [Google Scholar]
- Riedl, J.; Mordmüller, B.; Koder, S.; Pabinger, I.; Kremsner, P.G.; Hoffman, S.L.; Ramharter, M.; Ay, C. Alterations of blood coagulation in controlled human malaria infection. Malar. J. 2016, 15, 15. [Google Scholar] [CrossRef] [Green Version]
- Angchaisuksiri, P. Coagulopathy in malaria. Thromb. Res. 2014, 133, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Francischetti, I.M.B. Does activation of the blood coagulation cascade have a role in malaria pathogenesis? Trends Parasitol. 2008, 24, 258–263. [Google Scholar] [PubMed] [Green Version]
- Moussa, E.M.; Huang, H.; Thézénas, M.L.; Fischer, R.; Ramaprasad, A.; Sisay-Joof, F.; Jallow, M.; Pain, A.; Kwiatkowski, D.; Kessler, B.M.; et al. Proteomic profiling of the plasma of Gambian children with cerebral malaria. Malar. J. 2018, 17, 337. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, J.M.; Preston, R.J.S.; O’Regan, N.; O’Donnell, J.S. Emerging roles for hemostatic dysfunction in malaria pathogenesis. Blood 2016, 127, 2281–2288. [Google Scholar] [CrossRef] [Green Version]
- Idro, R.; Kakooza-Mwesige, A.; Asea, B.; Ssebyala, K.; Bangirana, P.; Opoka, R.O.; Lubowa, S.K.; Semrud-Clikeman, M.; John, C.C.; Nalugya, J. Cerebral malaria is associated with long-term mental health disorders: A cross sectional survey of a long-term cohort. Malar. J. 2016, 15, 184. [Google Scholar] [CrossRef] [Green Version]
- Nassor, F.; Jarray, R.; Biard, D.S.F.; Maïza, A.; Papy-Garcia, D.; Pavoni, S.; Deslys, J.-P.; Yates, F. Long Term Gene Expression in Human Induced Pluripotent Stem Cells and Cerebral Organoids to Model a Neurodegenerative Disease. Front. Cell. Neurosci. 2020, 14, 14. [Google Scholar] [CrossRef] [PubMed]
- Muzio, L.; Consalez, G.G. Modeling human brain development with cerebral organoids. Stem Cell Res. Ther. 2013, 4, 154. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.-T.; Bendriem, R.M.; Wu, W.W.; Shen, R.-F. 3D brain Organoids derived from pluripotent stem cells: Promising experimental models for brain development and neurodegenerative disorders. J. Biomed. Sci. 2017, 24, 59. [Google Scholar] [CrossRef] [Green Version]
- Lancaster, M.A.; Knoblich, J.A. Generation of cerebral organoids from human pluripotent stem cells. Nat. Protoc. 2014, 9, 2329–2340. [Google Scholar] [CrossRef] [Green Version]
- Lancaster, M.A.; Knoblich, J.A. Organogenesis in a dish: Modeling development and disease using organoid technologies. Science 2014, 345, 1247125. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Renner, M.; Martin, C.-A.; Wenzel, D.; Bicknell, L.S.; Hurles, M.E.; Homfray, T.; Penninger, J.M.; Jackson, A.P.; Knoblich, J.A. Cerebral organoids model human brain development and microcephaly. Nature 2013, 501, 373–379. [Google Scholar] [CrossRef] [Green Version]
- Paşca, A.M.; Sloan, S.A.; Clarke, L.E.; Tian, Y.; Makinson, C.D.; Huber, N.; Kim, C.H.; Park, J.-Y.; O’Rourke, N.A.; Nguyen, K.D.; et al. Functional cortical neurons and astrocytes from human pluripotent stem cells in 3D culture. Nat. Methods 2015, 12, 671–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alia, C.; Terrigno, M.; Busti, I.; Cremisi, F.; Caleo, M. Pluripotent Stem Cells for Brain Repair: Protocols and Preclinical Applications in Cortical and Hippocampal Pathologies. Front. Neurosci. 2019, 13, 684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costamagna, G.; Andreoli, L.; Corti, S.; Faravelli, I. iPSCs-Based Neural 3D Systems: A Multidimensional Approach for Disease Modeling and Drug Discovery. Cells 2019, 8, 1438. [Google Scholar] [PubMed] [Green Version]
- Brown, J.; Quadrato, G.; Arlotta, P. Studying the Brain in a Dish: 3D Cell Culture Models of Human Brain Development and Disease. Curr. Top. Dev. Biol. 2018, 129, 99–122. [Google Scholar] [CrossRef]
- Wang, H. Modeling Neurological Diseases with Human Brain Organoids. Front. Synaptic Neurosci. 2018, 10, 15. [Google Scholar] [CrossRef]
- Kaindl, J.; Winner, B. Disease Modeling of Neuropsychiatric Brain Disorders Using Human Stem Cell-Based Neural Models. Behav. Neurogenomics 2019, 42, 159–183. [Google Scholar] [CrossRef]
- Harbuzariu, A.; Pitts, S.; Cespedes, J.C.; Harp, K.O.; Nti, A.; Shaw, A.P.; Liu, M.; Stiles, J.K. Modelling heme-mediated brain injury associated with cerebral malaria in human brain cortical organoids. Sci. Rep. 2019, 9, 19162. [Google Scholar]
- Klotz, C.; Aebischer, T.; Seeber, F. Stem cell-derived cell cultures and organoids for protozoan parasite propagation and studying host–parasite interaction. Int. J. Med. Microbiol. 2012, 302, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Eigenmann, D.E.; Xue, G.; Kim, K.S.; Moses, A.V.; Hamburger, M.; Oufir, M. Comparative study of four immortalized human brain capillary endothelial cell lines, hCMEC/D3, hBMEC, TY10, and BB19, and optimization of culture conditions, for an in vitro blood–brain barrier model for drug permeability studies. Fluids Barriers CNS 2013, 10, 33. [Google Scholar] [CrossRef] [Green Version]
- Marote, A.; Santos, D.; Mendes-Pinheiro, B.; Serre-Miranda, C.; Anjo, S.I.; Vieira, J.; Ferreira-Antunes, F.; Correia, J.S.; Borges-Pereira, C.; Pinho, A.G.; et al. Cellular Aging Secretes: A Comparison of Bone-Marrow-Derived and Induced Mesenchymal Stem Cells and Their Secretome Over Long-Term Culture. Stem Cell Rev. Rep. 2023, 19, 248–263. [Google Scholar] [CrossRef] [PubMed]
- Garmire, L.X.; Subramaniam, S. Evaluation of normalization methods in mammalian microRNA-Seq data. RNA 2012, 18, 1279. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Li, N.; Liu, S.; Wang, J.; Zhang, N.; Zheng, X.; Leung, K.-S.; Cheng, L. Normalization Methods for the Analysis of Unbalanced Transcriptome Data: A Review. Front. Bioeng. Biotechnol. 2019, 7, 358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, R.; Anantharam, V.; Kanthasamy, A.G.; Kanthasamy, A. Proteolytic activation of proapoptotic kinase protein kinase Cδ by tumor necrosis factor α death receptor signaling in dopaminergic neurons during neuroinflammation. J. Neuroinflamm. 2012, 9, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGuire, S.O.; Ling, Z.D.; Lipton, J.W.; Sortwell, C.E.; Collier, T.J.; Carvey, P.M. Tumor Necrosis Factor α Is Toxic to Embryonic Mesencephalic Dopamine Neurons. Exp. Neurol. 2001, 169, 219–230. [Google Scholar] [CrossRef]
- McCoy, M.K.; Martinez, T.N.; Ruhn, K.A.; Szymkowski, D.E.; Smith, C.G.; Botterman, B.R.; Tansey, K.E.; Tansey, M.G. Blocking Soluble Tumor Necrosis Factor Signaling with Dominant-Negative Tumor Necrosis Factor Inhibitor Attenuates Loss of Dopaminergic Neurons in Models of Parkinson’s Disease. J. Neurosci. 2006, 26, 9365–9375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarrant, J.M. Blood Cytokines as Biomarkers of In Vivo Toxicity in Preclinical Safety Assessment: Considerations for Their Use. Toxicol. Sci. 2010, 117, 4–16. [Google Scholar] [CrossRef] [Green Version]
- Avril, M.; Benjamin, M.; Dols, M.-M.; Smith, J.D. Interplay of Plasmodium falciparum and thrombin in brain endothelial barrier disruption. Sci. Rep. 2019, 9, 13142. [Google Scholar] [CrossRef] [Green Version]
- Pais, T.F.; Penha-Gonçalves, C. Brain endothelium: The ‘innate immunity response hypothesis’ in cerebral malaria path-ogenesis. Front. Immunol. 2019, 10, 3100. [Google Scholar] [CrossRef]
- Sun, A.X.; Yuan, Q.; Tan, S.; Xiao, Y.; Wang, D.; Khoo, A.T.T.; Sani, L.; Tran, H.-D.; Kim, P.; Chiew, Y.S.; et al. Direct Induction and Functional Maturation of Forebrain GABAergic Neurons from Human Pluripotent Stem Cells. Cell Rep. 2016, 16, 1942–1953. [Google Scholar] [CrossRef] [Green Version]
- Deininger, M.H.; Kremsner, P.G.; Meyermann, R.; Schluesener, H.J. Focal accumulation of cyclooxygenase-1 (COX-1) and COX-2 expressing cells in cerebral malaria. J. Neuroimmunol. 2000, 106, 198–205. [Google Scholar] [CrossRef]
- Ball, H.J.; MacDougall, H.; McGregor, I.S.; Hunt, N.H. Cyclooxygenase-2 in the Pathogenesis of Murine Cerebral Malaria. J. Infect. Dis. 2004, 189, 751–758. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, E.; Sadoughi, F.; Younesi, S.; Karimian, A.; Asemi, Z.; Farsad-Akhtar, N.; Jahanbakhshi, F.; Jamilian, H.; Yousefi, B. The molecular mechanism of nuclear signaling for degradation of cytoplasmic DNA: Importance in DNA damage response and cancer. DNA Repair 2021, 103, 103115. [Google Scholar] [CrossRef]
- Ye, Z.; Xue, A.; Huang, Y.; Wu, Q. Children with cyclic vomiting syndrome: Phenotypes, disease burden and mitochondrial DNA analysis. BMC Gastroenterol. 2018, 18, 104. [Google Scholar] [CrossRef] [PubMed]
- Tiihonen, J.; Koskuvi, M.; Lähteenvuo, M.; Virtanen, P.L.J.; Ojansuu, I.; Vaurio, O.; Gao, Y.; Hyötyläinen, I.; Puttonen, K.A.; Repo-Tiihonen, E.; et al. Neurobiological roots of psychopathy. Mol. Psychiatry 2020, 25, 3432–3441. [Google Scholar] [CrossRef] [PubMed]
- Osmanagic-Myers, S.; Rus, S.; Wolfram, M.; Brunner, D.; Goldmann, W.H.; Bonakdar, N.; Fischer, I.; Reipert, S.; Zuzuarregui, A.; Walko, G.; et al. Plectin reinforces vascular integrity by mediating crosstalk between the vimentin and the actin networks. Development 2015, 142, e1.1. [Google Scholar] [CrossRef]
- Suttitheptumrong, A.; Rawarak, N.; Reamtong, O.; Boonnak, K.; Pattanakitsakul, S.-N. Plectin is Required for Trans-Endothelial Permeability: A Model of Plectin Dysfunction in Human Endothelial Cells After TNF-α Treatment and Dengue Virus Infection. Proteomics 2018, 18, e1800215. [Google Scholar] [CrossRef]
- Potokar, M.; Morita, M.; Wiche, G.; Jorgačevski, J. The Diversity of Intermediate Filaments in Astrocytes. Cells 2020, 9, 1604. [Google Scholar] [CrossRef]
- Thierry, A.; Falilatou, A.; Covalic, B.; Elodie, D.; Mendinatou, A.; Didier, A.; Alphonse, N.; Joseph, A. Epilepsy and Malaria in Children Aged 1 to 15 Years in Parakou in 2018: Case-Control Study. Child Neurol. Open 2020, 7, 2329048X20954111. [Google Scholar] [CrossRef] [PubMed]
- del Valle-Pérez, B.; Martínez, V.G.; Lacasa-Salavert, C.; Figueras, A.; Shapiro, S.S.; Takafuta, T.; Casanovas, O.; Capellà, G.; Ventura, F.; Viñals, F. Filamin B Plays a Key Role in Vascular Endothelial Growth Factor-induced Endothelial Cell Motility through Its Interaction with Rac-1 and Vav-2. J. Biol. Chem. 2010, 285, 10748–10760. [Google Scholar] [CrossRef] [Green Version]
- Kanters, E.; van Rijssel, J.; Hensbergen, P.J.; Hondius, D.; Mul, F.P.; Deelder, A.M.; Sonnenberg, A.; van Buul, J.D.; Hordijk, P.L. Filamin B Mediates ICAM-1-driven Leukocyte Transendothelial Migration. J. Biol. Chem. 2008, 283, 31830–31839. [Google Scholar] [CrossRef] [Green Version]
- Bandaru, S.; Zhou, A.-X.; Rouhi, P.; Zhang, Y.; Bergo, M.O.; Cao, Y.; Akyürek, L.M. Targeting filamin B induces tumor growth and metastasis via enhanced activity of matrix metalloproteinase-9 and secretion of VEGF-A. Oncogenesis 2014, 3, e119. [Google Scholar] [CrossRef] [Green Version]
- Schaefer, A.; Riet, J.T.; Ritz, K.; Hoogenboezem, M.; Anthony, E.C.; Mul, F.P.J.; de Vries, C.J.; Daemen, M.; Figdor, C.; Van Buul, J.; et al. Actin-binding proteins differentially regulate endothelial cell stiffness, ICAM-1 function and neutrophil transmigration. J. Cell Sci. 2014, 127, 4470–4482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Liu, H.; Wu, J.; Zhao, Y. miR-125a Suppresses TrxR1 Expression and Is Involved in H2O2-Induced Oxidative Stress in Endothelial Cells. J. Immunol. Res. 2018, 2018, 6140320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakurai, A.; Yuasa, K.; Shoji, Y.; Himeno, S.; Tsujimoto, M.; Kunimoto, M.; Imura, N.; Hara, S. Overexpression of thioredoxin reductase 1 regulates NF-kappa B activation. J. Cell. Physiol. 2004, 198, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Kudin, A.P.; Baron, G.; Zsurka, G.; Hampel, K.G.; Elger, C.E.; Grote, A.; Weber, Y.; Lerche, H.; Thiele, H.; Nürnberg, P.; et al. Homozygous mutation in TXNRD1 is associated with genetic generalized epilepsy. Free Radic. Biol. Med. 2017, 106, 270–277. [Google Scholar] [CrossRef]
- Yu, J.-T.; Liu, Y.; Dong, P.; Cheng, R.-E.; Ke, S.-X.; Chen, K.-Q.; Wang, J.-J.; Shen, Z.-S.; Tang, Q.-Y.; Zhang, Z. Up-regulation of antioxidative proteins TRX1, TXNL1 and TXNRD1 in the cortex of PTZ kindling seizure model mice. PLoS ONE 2019, 14, e0210670. [Google Scholar] [CrossRef]
- Lennartz, F.; Smith, C.; Craig, A.G.; Higgins, M.K. Structural insights into diverse modes of ICAM-1 binding by Plasmodium falciparum-infected erythrocytes. Proc. Natl. Acad. Sci. USA 2019, 116, 20124–20134. [Google Scholar] [CrossRef] [Green Version]
- Bhalla, K.; Chugh, M.; Mehrotra, S.; Rathore, S.; Tousif, S.; Dwivedi, V.P.; Prakash, P.; Samuchiwal, S.; Kumar, S.; Singh, D.; et al. Host ICAMs play a role in cell invasion by Mycobacterium tuberculosis and Plasmodium falciparum. Nat. Commun. 2015, 6, 6049. [Google Scholar] [CrossRef] [Green Version]
- Gu, P.; Theiss, A.; Han, J.; Feagins, L.A. Increased Cell Adhesion Molecules, PECAM-1, ICAM-3, or VCAM-1, Predict Increased Risk for Flare in Patients with Quiescent Inflammatory Bowel Disease. J. Clin. Gastroenterol. 2017, 51, 522–527. [Google Scholar] [CrossRef]
- Mahamar, A.; Attaher, O.; Swihart, B.; Barry, A.; Diarra, B.S.; Kanoute, M.B.; Cisse, K.B.; Dembele, A.B.; Keita, S.; Gamain, B.; et al. Host factors that modify Plasmodium falciparum adhesion to endothelial receptors. Sci. Rep. 2017, 7, 13872. [Google Scholar] [CrossRef] [Green Version]
- Che, J.N.; Nmorsi, O.P.G.; Nkot, B.P.; Isaac, C.; Okonkwo, B.C. Chemokines responses to Plasmodium falciparum malaria and co-infections among rural Cameroonians. Parasitol. Int. 2015, 64, 139–144. [Google Scholar] [CrossRef]
- Feng, X.; Ma, B.-F.; Liu, B.; Ding, P.; Wei, J.-H.; Cheng, P.; Li, S.-Y.; Chen, D.-X.; Sun, Z.-J.; Li, Z. The Involvement of the Chemokine RANTES in Regulating Luminal Acidification in Rat Epididymis. Front. Immunol. 2020, 11, 583274. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, S.S.; Carret, C.; Grosso, A.R.; Tarun, A.S.; Peng, X.; Kappe, S.H.; Prudêncio, M.; Mota, M.M. Host cell transcriptional profiling during malaria liver stage infection reveals a coordinated and sequential set of biological events. BMC Genom. 2009, 10, 270. [Google Scholar] [CrossRef] [Green Version]
- Bando, H.; Pradipta, A.; Iwanaga, S.; Okamoto, T.; Okuzaki, D.; Tanaka, S.; Vega-Rodríguez, J.; Lee, Y.; Ma, J.S.; Sakaguchi, N.; et al. CXCR4 regulates Plasmodium development in mouse and human hepatocytes. J. Exp. Med. 2019, 216, 1733–1748. [Google Scholar] [CrossRef] [PubMed]
- Ioannidis, L.J.; Nie, C.Q.; Hansen, D.S. The role of chemokines in severe malaria: More than meets the eye. Parasitology 2014, 141, 602–613. [Google Scholar] [CrossRef] [PubMed]
- Sercundes, M.K.; Ortolan, L.S.; Debone, D.; Soeiro-Pereira, P.V.; Gomes, E.; Aitken, E.H.; Neto, A.C.; Russo, M.; Lima, M.R.D.I.; Alvarez, J.M.; et al. Targeting Neutrophils to Prevent Malaria-Associated Acute Lung Injury/Acute Respiratory Distress Syndrome in Mice. PLoS Pathog. 2016, 12, e1006054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, D.S.A.; Rodrigues, D.; Prestes, E.B.; de Souza Silva, L.; Pinheiro, A.A.S.; Ribeiro, J.M.C.; Dicko, A.; Duffy, P.E.; Fried, M.; Francischetti, I.M.B.; et al. CXCR4 and MIF are required for neutrophil extracellular trap release triggered by Plasmodium-infected erythrocytes. bioRxiv 2019, 16, 852574. [Google Scholar] [CrossRef] [Green Version]
- Dunst, J.; Kamena, F.; Matuschewski, K. Cytokines and Chemokines in Cerebral Malaria Pathogenesis. Front. Cell. Infect. Microbiol. 2017, 7, 324. [Google Scholar] [CrossRef] [Green Version]
- Wangala, B.; Vovor, A.; Gantin, R.; Agbeko, Y.; Lechner, C.; Huang, X.; Soboslay, P.; Köhler, C. Chemokine levels and parasite- and allergen-specific antibodyresponses in children and adults with severe or uncomplicated Plasmodium falciparum malaria. Eur. J. Microbiol. Immunol. 2015, 5, 131. [Google Scholar] [CrossRef] [Green Version]
- Abrams, E.T.; Brown, H.; Chensue, S.W.; Turner, G.D.H.; Tadesse, E.; Lema, V.M.; Molyneux, M.E.; Rochford, R.; Meshnick, S.R.; Rogerson, S.J. Host Response to Malaria During Pregnancy: Placental Monocyte Recruitment Is Associated with Elevated β Chemokine Expression. J. Immunol. 2003, 170, 2759–2764. [Google Scholar] [CrossRef] [Green Version]
- Hojo-Souza, N.S.; Pereira, D.B.; de Souza, F.S.H.; Mendes, T.A.D.O.; Cardoso, M.S.; Tada, M.S.; Zanini, G.M.; Bartholomeu, D.C.; Fujiwara, R.T.; Bueno, L.L. On the cytokine/chemokine network during Plasmodium vivax malaria: New insights to understand the disease. Malar. J. 2017, 16, 42. [Google Scholar] [CrossRef] [Green Version]
- Raacke, M.; Kerr, A.; Dörpinghaus, M.; Brehmer, J.; Wu, Y.; Lorenzen, S.; Fink, C.; Jacobs, T.; Roeder, T.; Sellau, J.; et al. Altered Cytokine Response of Human Brain Endothelial Cells after Stimulation with Malaria Patient Plasma. Cells 2021, 10, 1656. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Haus, J.M.; Chen, L.; Wu, S.C.; Urao, N.; Koh, T.J.; Minshall, R.D. CCL28-induced CCR10/eNOS Interaction in Angiogenesis and Skin Wound Healing. FASEB J. 2020, 34, 5838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, F.S.; Desruisseaux, M.S.; Kennan, R.P.; Hetherington, H.P.; Wittner, M.; Weiss, L.M.; Lee, S.C.; Scherer, P.E.; Tsuji, M.; Tanowitz, H.B.; et al. Endothelin in a murine model of cerebral malaria. Exp. Biol. Med. 2017, 231, 1176–1181. [Google Scholar]
- Freeman, B.D.; Martins, Y.C.; Akide-Ndunge, O.B.; Bruno, F.P.; Wang, H.; Tanowitz, H.B.; Spray, D.C.; Desruisseaux, M.S. Endothelin-1 Mediates Brain Microvascular Dysfunction Leading to Long-Term Cognitive Impairment in a Model of Experimental Cerebral Malaria. PLoS Pathog. 2016, 12, e1005477. [Google Scholar] [CrossRef]
- Martins, Y.C.; Freeman, B.D.; Ndunge, O.B.A.; Weiss, L.M.; Tanowitz, H.B.; Desruisseaux, M.S. Endothelin-1 Treatment Induces an Experimental Cerebral Malaria-Like Syndrome in C57BL/6 Mice Infected with Plasmodium berghei NK65. Am. J. Pathol. 2016, 186, 2957–2969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenisch, C.; Wenisch, H.; Wilairatana, P.; Looareesuwan, S.; Vannaphan, S.; Wagner, O.; Graninger, W.; Schönthal, E.; Rumpold, H. Big Endothelin in Patients with Complicated Plasmodium falciparum Malaria. J. Infect. Dis. 1996, 173, 1281–1284. [Google Scholar] [CrossRef] [Green Version]
- Dietmann, A.; Lackner, P.; Helbok, R.; Spora, K.; Issifou, S.; Lell, B.; Reindl, M.; Kremsner, P.; Schmutzhard, E. Opposed circulating plasma levels of endothelin-1 and C-type natriuretic peptide in children with Plasmodium falciparum malaria. Malar. J. 2008, 7, 253. [Google Scholar] [CrossRef] [Green Version]
- Colborn, J.M.; Ylöstalo, J.H.; Koita, O.A.; Cissé, O.H.; Krogstad, D.J. Human Gene Expression in Uncomplicated Plasmodium falciparum Malaria. J. Immunol. Res. 2015, 2015, 162639. [Google Scholar] [CrossRef] [Green Version]
- Graham, S.M.; Chen, J.; Chung, D.W.; Barker, K.R.; Conroy, A.L.; Hawkes, M.T.; Namasopo, S.; Kain, K.C.; López, J.A.; Liles, W.C. Endothelial activation, haemostasis and thrombosis biomarkers in Ugandan children with severe malaria participating in a clinical trial. Malar. J. 2016, 15, 56. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, A.S.; Fazavana, J.; O’Donnell, J.S. The von Willebrand factor—ADAMTS-13 axis in malaria. Res. Pract. Thromb. Haemost. 2022, 6, e12641. [Google Scholar] [CrossRef] [PubMed]
- Mbagwu, S.I.; Filgueira, L. Differential Expression of CD31 and Von Willebrand Factor on Endothelial Cells in Different Regions of the Human Brain: Potential Implications for Cerebral Malaria Pathogenesis. Brain Sci. 2020, 10, 31. [Google Scholar] [CrossRef] [Green Version]
- O’Regan, N.; Gegenbauer, K.; O’Sullivan, J.M.; Maleki, S.; Brophy, T.M.; Dalton, N.; Chion, A.; Fallon, P.G.; Grau, G.; Budde, U.; et al. A novel role for von Willebrand factor in the pathogenesis of experimental cerebral malaria. Blood 2016, 127, 1192–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraisin, S.; Martinod, K.; Desender, L.; Pareyn, I.; Verhenne, S.; Deckmyn, H.; Vanhoorelbeke, K.; Van den Steen, P.E.; De Meyer, S.F. von Willebrand factor increases in experimental cerebral malaria but is not essential for late-stage patho-genesis in mice. J. Thromb. Haemost. 2020, 18, 2377–2390. [Google Scholar] [CrossRef]
- Hollestelle, M.J.; Donkor, C.; Mantey, E.A.; Chakravorty, S.J.; Craig, A.; Akoto, A.O.; O’Donnell, J.; van Mourik, J.A.; Bunn, J. von Willebrand factor propeptide in malaria: Evidence of acute endothelial cell activation. Br. J. Haematol. 2006, 133, 562–569. [Google Scholar] [CrossRef]
- Cox, P.R.; Fowler, V.; Xu, B.; Sweatt, J.; Paylor, R.; Zoghbi, H.Y. Mice lacking tropomodulin-2 show enhanced long-term potentiation, hyperactivity, and deficits in learning and memory. Mol. Cell. Neurosci. 2003, 23, 1–12. [Google Scholar] [CrossRef]
- Armah, H.B.; Wilson, N.O.; Sarfo, B.Y.; Powell, M.D.; Bond, V.C.; Anderson, W.; Adjei, A.A.; Gyasi, R.K.; Tettey, Y.; Wiredu, E.K.; et al. Cerebrospinal fluid and serum biomarkers of cerebral malaria mortality in Ghanaian children. Malar. J. 2007, 6, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polimeni, M.; Prato, M. Host matrix metalloproteinases in cerebral malaria: New kids on the block against blood–brain barrier integrity? Fluids Barriers CNS 2014, 11, 1. [Google Scholar] [CrossRef]
- Prato, M.; Giribaldi, G. Matrix Metalloproteinase-9 and Haemozoin: Wedding Rings for Human Host and Plasmodium falciparum Parasite in Complicated Malaria. J. Trop. Med. 2011, 2011, 628435. [Google Scholar] [CrossRef] [Green Version]
- Polimeni, M.; Valente, E.; Ulliers, D.; Opdenakker, G.; Van den Steen, P.E.; Giribaldi, G.; Prato, M. Natural haemozoin induces expression and release of human monocyte tissue inhibitor of metallopro-teinase-1. PLoS ONE 2013, 8, e71468. [Google Scholar] [CrossRef] [Green Version]
- Prato, M.; D’Alessandro, S.; Van den Steen, P.E.; Opdenakker, G.; Arese, P.; Taramelli, D.; Basilico, N. Natural haemozoin modulates matrix metalloproteinases and induces morphological changes in human microvascular endothelium. Cell. Microbiol. 2011, 13, 1275–1285. [Google Scholar] [CrossRef]
- Mandala, W.L.; Msefula, C.L.; Gondwe, E.N.; Drayson, M.T.; Molyneux, M.E.; MacLennan, C.A. Cytokine Profiles in Malawian Children Presenting with Uncomplicated Malaria, Severe Malarial Anemia, and Cerebral Malaria. Clin. Vaccine Immunol. 2017, 24, e00533-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bujarbaruah, D.; Kalita, M.P.; Baruah, V.; Basumatary, T.K.; Hazarika, S.; Begum, R.H.; Medhi, S.; Bose, S. RANTES levels as a determinant of falciparum malaria severity or recovery. Parasite Immunol. 2017, 39, e12452. [Google Scholar] [CrossRef]
- Ochiel, D.O.; Awandare, G.A.; Keller, C.C.; Hittner, J.B.; Kremsner, P.G.; Weinberg, J.B.; Perkins, D.J. Differential Regulation of β-Chemokines in Children with Plasmodium falciparum Malaria. Infect. Immun. 2005, 73, 4190–4197. [Google Scholar] [CrossRef] [Green Version]
- Camp, J.G.; Badsha, F.; Florio, M.; Kanton, S.; Gerber, T.; Wilsch-Bräuninger, M.; Lewitus, E.; Sykes, A.; Hevers, W.; Lancaster, M.A.; et al. Human cerebral organoids recapitulate gene expression programs of fetal neocortex development. Proc. Natl. Acad. Sci. USA 2015, 112, 15672–15677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciarpella, F.; Zamfir, R.G.; Campanelli, A.; Ren, E.; Pedrotti, G.; Bottani, E.; Borioli, A.; Caron, D.; Di Chio, M.; Dolci, S.; et al. Murine cerebral organoids develop network of functional neurons and hippocampal brain region identity. iScience 2021, 24, 103438. [Google Scholar] [CrossRef]
- Dezonne, R.S.; Sartore, R.C.; Nascimento, J.M.; Saia-Cereda, V.M.; Romão, L.F.; Alves-Leon, S.V.; de Souza, J.M.; Martins-De-Souza, D.; Rehen, S.K.; Gomes, F.C.A. Derivation of Functional Human Astrocytes from Cerebral Organoids. Sci. Rep. 2017, 7, srep45091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nascimento, J.M.; Saia-Cereda, V.M.; Sartore, R.C.; da Costa, R.M.; Schitine, C.S.; Freitas, H.R.; Murgu, M.; Reis, R.A.D.M.; Rehen, S.K.; Martins-De-Souza, D. Human Cerebral Organoids and Fetal Brain Tissue Share Proteomic Similarities. Front. Cell Dev. Biol. 2019, 7, 303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, D.; Xiang, A.P.; Mao, F.F.; Zhang, L.; Di, C.-G.; Liu, X.-M.; Shao, Y.; Ma, B.-F.; Lee, J.-H.; Ha, K.-S.; et al. Nestin Is Required for the Proper Self-Renewal of Neural Stem Cells. Stem Cells 2010, 28, 2162–2171. [Google Scholar] [CrossRef]
- Huang, S.; Zhang, Z.; Cao, J.; Yu, Y.; Pei, G. Chimeric cerebral organoids reveal the essentials of neuronal and astrocytic APOE4 for Alzheimer’s tau pathology. Signal Transduct. Target. Ther. 2022, 7, 176. [Google Scholar] [CrossRef] [PubMed]
- Yakoub, A.M. Cerebral organoids exhibit mature neurons and astrocytes and recapitulate electrophysiological activity of the human brain. Neural Regen. Res. 2019, 14, 757–761. [Google Scholar] [CrossRef]
- Renner, M.; Lancaster, M.A.; Bian, S.; Choi, H.; Ku, T.; Peer, A.; Chung, K.; Knoblich, J.A. Self-organized developmental patterning and differentiation in cerebral organoids. EMBO J. 2017, 36, 1316–1329. [Google Scholar] [CrossRef]
- Glushakova, O.Y.; Glushakov, A.A.; Wijesinghe, D.S.; Valadka, A.B.; Hayes, R.L.; Glushakov, A.V. Prospective clinical biomarkers of caspase-mediated apoptosis associated with neuronal and neuro-vascular damage following stroke and other severe brain injuries: Implications for chronic neurodegeneration. Brain Circ. 2017, 3, 87. [Google Scholar] [CrossRef]
- Ramirez, S.; Mukherjee, A.; Sepulveda, S.; Becerra-Calixto, A.; Bravo-Vasquez, N.; Gherardelli, C.; Chavez, M.; Soto, C. Modeling Traumatic Brain Injury in Human Cerebral Organoids. Cells 2021, 10, 2683. [Google Scholar] [CrossRef] [PubMed]
- An, H.L.; Kuo HCTang, T.K. Modeling Human Primary Microcephaly With hiPSC-Derived Brain Organoids Carrying CPAP-E1235V Disease-Associated Mutant Protein. Front. Cell Dev. Biol. 2022, 10, 451. [Google Scholar]
- Hyland, R.M.; Brody, S.L. Impact of Motile Ciliopathies on Human Development and Clinical Consequences in the Newborn. Cells 2021, 11, 125. [Google Scholar] [CrossRef]
- Ki, S.M.; Jeong, H.S.; Lee, J.E. Primary Cilia in Glial Cells: An Oasis in the Journey to Overcoming Neurodegenerative Diseases. Front. Neurosci. 2021, 15, 736888. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Gleeson, J.G. Cilia in the nervous system: Linking cilia function and neurodevelopmental disorders. Curr. Opin. Neurol. 2011, 24, 98–105. [Google Scholar] [CrossRef] [Green Version]
- Ringers, C.; Olstad, E.W.; Jurisch-Yaksi, N. The role of motile cilia in the development and physiology of the nervous system. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190156. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Yang, S.-L.; Yang, M.; Herrlinger, S.; Shao, Q.; Collar, J.L.; Fierro, E.; Shi, Y.; Liu, A.; Lu, H.; et al. Modeling microcephaly with cerebral organoids reveals a WDR62–CEP170–KIF2A pathway promoting cilium disassembly in neural progenitors. Nat. Commun. 2019, 10, 2612. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P. falciparum Strain & Time-Point | Pro-Inflammatory | Anti-Inflammatory | p-Value | Fold-Change | Increase/Decrease |
|---|---|---|---|---|---|
| 3D7_4h | RANTES | 0.0001 | 0.2 | ↓ | |
| IL-6 | IL-6 | 0.0002 | 2.1 | ↑ | |
| MCP-1 | 0.0001 | 2.1 | ↑ | ||
| IL-8 | 0.0006 | 2.7 | ↑ | ||
| HB3_4h | RANTES | 0.0001 | 0.2 | ↓ | |
| IL-6 | IL-6 | 0.0001 | 2.3 | ↑ | |
| Dd2_4h | RANTES | 0.0001 | 0.3 | ↓ | |
| IL-6 | IL-6 | 0.0001 | 2.0 | ↑ | |
| 3D7_24h | RANTES | 0.0001 | 0.7 | ↓ | |
| IL-6 | IL-6 | 0.0001 | 2.5 | ↑ | |
| MCP-1 | 0.0006 | 2.6 | ↑ | ||
| PDGF-BB | 0.0022 | 2.6 | ↑ | ||
| GM-CSF | 0.0001 | 4.4 | ↑ | ||
| TIMP-2 | 0.0005 | 2.4 | ↑ | ||
| IL-8 | 0.0101 | 2.7 | ↑ | ||
| HB3_24h | RANTES | 0.0105 | 0.7 | ↓ | |
| IL-6 | IL-6 | 0.0001 | 2.0 | ↑ | |
| MCP-1 | 0.0006 | 2.4 | ↑ | ||
| PDGF-BB | 0.0209 | 2.3 | ↑ | ||
| GM-CSF | 0.0224 | 0.7 | ↑ | ||
| TIMP-2 | 0.0001 | 2.3 | ↑ | ||
| Dd2_24h | IL-6 | IL-6 | 0.0001 | 2.6 | ↑ |
| MCP-1 | 0.0001 | 2.8 | ↑ | ||
| PDGF-BB | 0.0044 | 2.5 | ↑ | ||
| GM-CSF | 0.0042 | 3.2 | ↑ | ||
| TIMP-2 | 0.0016 | 2.5 | ↑ | ||
| IL-8 | 0.0097 | 2.7 | ↑ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva-Pedrosa, R.; Campos, J.; Fernandes, A.M.; Silva, M.; Calçada, C.; Marote, A.; Martinho, O.; Veiga, M.I.; Rodrigues, L.R.; Salgado, A.J.; et al. Cerebral Malaria Model Applying Human Brain Organoids. Cells 2023, 12, 984. https://doi.org/10.3390/cells12070984
Silva-Pedrosa R, Campos J, Fernandes AM, Silva M, Calçada C, Marote A, Martinho O, Veiga MI, Rodrigues LR, Salgado AJ, et al. Cerebral Malaria Model Applying Human Brain Organoids. Cells. 2023; 12(7):984. https://doi.org/10.3390/cells12070984
Chicago/Turabian StyleSilva-Pedrosa, Rita, Jonas Campos, Aline Marie Fernandes, Miguel Silva, Carla Calçada, Ana Marote, Olga Martinho, Maria Isabel Veiga, Ligia R. Rodrigues, António José Salgado, and et al. 2023. "Cerebral Malaria Model Applying Human Brain Organoids" Cells 12, no. 7: 984. https://doi.org/10.3390/cells12070984