
Combined Insults of a MASH Diet and Alcohol Binges Activate Intercellular Communication and Neutrophil Recruitment via the NLRP3-IL-1β Axis in the Liver


 , , and
, , and
Abstract
1. Introduction
2. Material and Method
2.1. Animal Care and Treatments
2.2. Neutrophil Isolation
2.3. Neutrophil Extracellular Trap (NET) Formation Assay
2.4. Western Blot and ELISA
2.5. Flow Cytometry
2.6. RNA Extraction and qPCR
3. Results
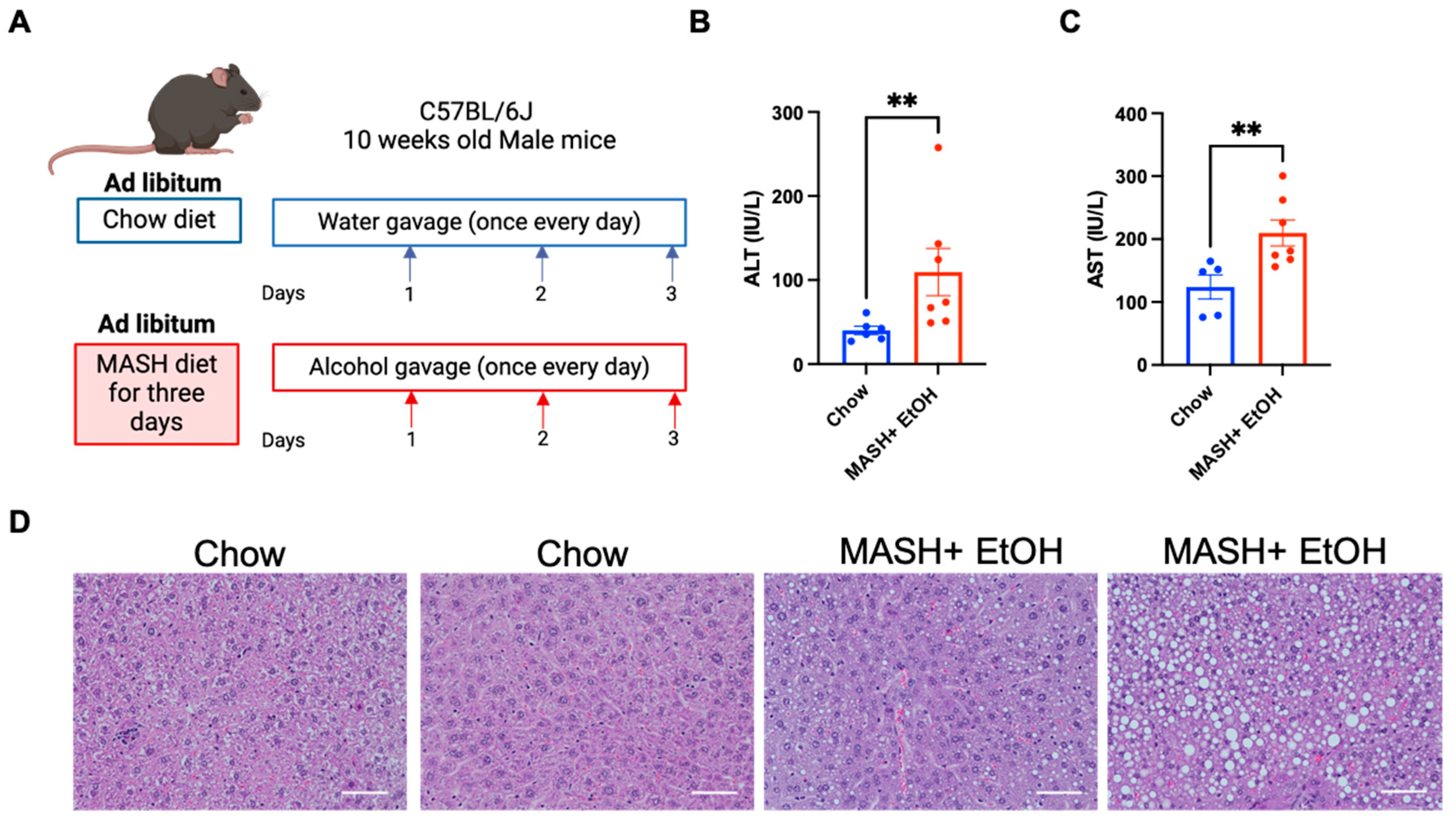
3.1. Short-Term Feeding of MASH Diet and Alcohol Binges Promote Liver Injury and Steatosis
3.2. Alcohol Binges and MASH Diet Leads to NLRP3 Inflammasome Activation
3.3. Short-Term Feeding of MASH Diet and Alcohol Binges Induce Hepatic Neutrophil Infiltration and NET Formation
3.4. Inhibition of NLRP3 Inflammasome by MCC950 Ameliorates Liver Damage
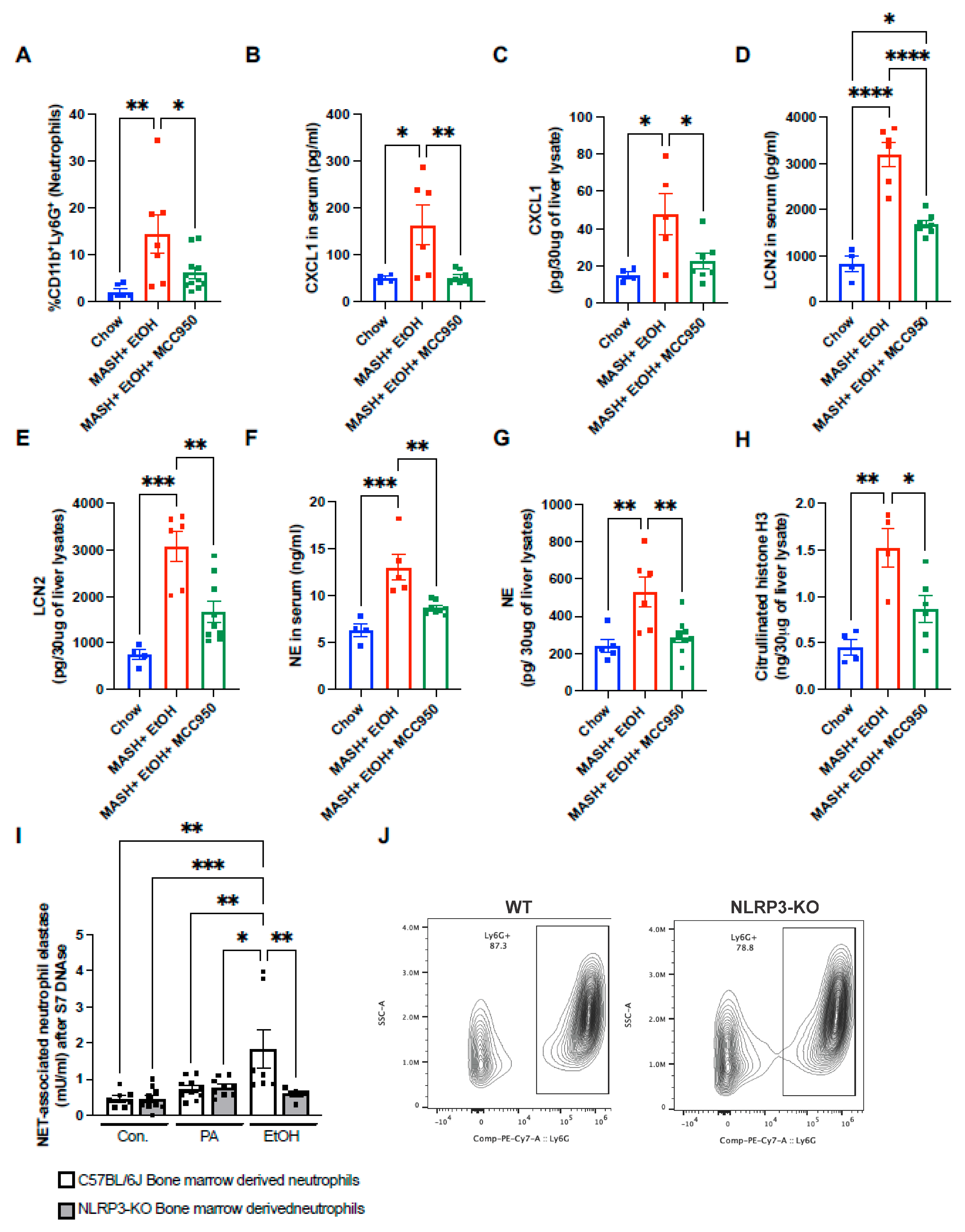
3.5. NLRP3 Inflammasome Inhibition Attenuates Neutrophil Infiltration and NET Formation
3.6. Exogenous IL-1β Regulates Production of CXCL1 and LCN2 from Primary Hepatocytes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AH | Alcoholic hepatitis/Alcohol-associated hepatitis |
| ALD | Alcoholic liver disease/Alcohol-associated liver disease |
| ALT | Alanine aminotransferase |
| ASC | Apoptosis-associated speck-like protein containing a caspase activating and recruitment domain |
| AST | Aspartate aminotransferase |
| CCL2/MCP-1 | chemokine (C-C motif) ligand 2/Monocyte chemoattractant protein 1 |
| Cit-H3 | Citrullinated histone 3 |
| CXCL1 | Chemokine (C-X-C motif) ligand 1 |
| CXCL2 | Chemokine (C-X-C motif) ligand 2 |
| CXCR1 | C-X-C Motif Chemokine Receptor 1 |
| CXCR2 | C-X-C Motif Chemokine Receptor 2 |
| DAMPs | Damage-associated molecular patterns |
| HF-HC-HS | High fat–cholesterol–sucrose diet |
| HFD | High-fat diet |
| HSC | Hepatic stellate cell |
| IL-1β | Interleukin 1 beta |
| IL-18 | Interleukin 18 |
| LCN2 | Lipocalin 2 |
| LMNCs | Liver mononuclear cells |
| MASH | Metabolic-dysfunction-associated steatohepatitis |
| MASLD: | Metabolic-dysfunction-associated steatotic liver disease |
| MFI | Mean fluorescent intensity |
| NE | Neutrophil elastase |
| NET(s) | Neutrophil extracellular trap(s) |
| NLRP3 | Nod-like receptor protein 3 |
| PAMPs | Pathogen-associated molecular pattern molecules |
| TLR | Toll-like receptor |
| TNF-α | Tumor necrosis factor alpha |
References
- Devarbhavi, H.; Asrani, S.K.; Arab, J.P.; Nartey, Y.A.; Pose, E.; Kamath, P.S. Global burden of liver disease: 2023 update. J. Hepatol. 2023, 79, 516–537. [Google Scholar] [CrossRef] [PubMed]
- Rehm, J.; Shield, K.D. Global Burden of Alcohol Use Disorders and Alcohol Liver Disease. Biomedicines 2019, 7, 99. [Google Scholar] [CrossRef] [PubMed]
- Teng, M.L.; Ng, C.H.; Huang, D.Q.; Chan, K.E.; Tan, D.J.; Lim, W.H.; Yang, J.D.; Tan, E.; Muthiah, M.D. Global incidence and prevalence of nonalcoholic fatty liver disease. Clin. Mol. Hepatol. 2023, 29, S32–S42. [Google Scholar] [CrossRef] [PubMed]
- Raza, S.A.; Sokale, I.O.; Thrift, A.P. Burden of high-risk phenotype of heavy alcohol consumption among obese U.S. population: Results from National Health and Nutrition Examination Survey, 1999–2020. Lancet Reg. Health Am. 2023, 23, 100525. [Google Scholar] [CrossRef] [PubMed]
- Boyle, M.; Masson, S.; Anstee, Q.M. The bidirectional impacts of alcohol consumption and the metabolic syndrome: Cofactors for progressive fatty liver disease. J. Hepatol. 2018, 68, 251–267. [Google Scholar] [CrossRef] [PubMed]
- Wandji, L.C.N.; Gnemmi, V.; Mathurin, P.; Louvet, A. Combined alcoholic and non-alcoholic steatohepatitis. JHEP Rep. 2020, 2, 100101. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Stepanova, M.; Ong, J.; Yilmaz, Y.; Duseja, A.; Eguchi, Y.; El Kassas, M.; Castellanos-Fernandez, M.; George, J.; Jacobson, I.M.; et al. Effects of Alcohol Consumption and Metabolic Syndrome on Mortality in Patients with Nonalcoholic and Alcohol-Related Fatty Liver Disease. Clin. Gastroenterol. Hepatol. 2019, 17, 1625–1633 e1. [Google Scholar] [CrossRef] [PubMed]
- Decraecker, M.; Dutartre, D.; Hiriart, J.; Irles-Depé, M.; Grottes, H.M.D.; Chermak, F.; Foucher, J.; Delamarre, A.; de Ledinghen, V. Long-term prognosis of patients with alcohol-related liver disease or non-alcoholic fatty liver disease according to metabolic syndrome or alcohol use. Liver Int. 2022, 42, 350–362. [Google Scholar] [CrossRef] [PubMed]
- Díaz, L.A.; Arab, J.P.; Louvet, A.; Bataller, R.; Arrese, M. The intersection between alcohol-related liver disease and nonalcoholic fatty liver disease. Nat. Rev. Gastroenterol. Hepatol. 2023, 20, 764–783. [Google Scholar] [CrossRef]
- Inan-Eroglu, E.; Huang, B.-H.; Ahmadi, M.N.; Johnson, N.; El-Omar, E.M.; Stamatakis, E. Joint associations of adiposity and alcohol consumption with liver disease-related morbidity and mortality risk: Findings from the UK Biobank. Eur. J. Clin. Nutr. 2022, 76, 74–83. [Google Scholar] [CrossRef]
- Kuntsche, E.; Kuntsche, S.; Thrul, J.; Gmel, G. Binge drinking: Health impact, prevalence, correlates and interventions. Psychol. Health 2017, 32, 976–1017. [Google Scholar] [CrossRef] [PubMed]
- Lazarus, J.V.; Newsome, P.N.; Francque, S.M.; Kanwal, F.; Terrault, N.A.; Rinella, M.E. Reply: A multi-society Delphi consensus statement on new fatty liver disease nomenclature. Hepatology 2024, 79, E93–E94. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho Ribeiro, M.; Szabo, G. Role of the Inflammasome in Liver Disease. Annu. Rev. Pathol. 2022, 17, 345–365. [Google Scholar] [CrossRef] [PubMed]
- Mandrekar, P.; Szabo, G. Signalling pathways in alcohol-induced liver inflammation. J. Hepatol. 2009, 50, 1258–1266. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wu, M. Pattern recognition receptors in health and diseases. Signal. Transduct. Target. Ther. 2021, 6, 291. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Hong, W.; Lu, S.; Li, Y.; Guan, Y.; Weng, X.; Feng, Z. The NLRP3 Inflammasome in Non-Alcoholic Fatty Liver Disease and Steatohepatitis: Therapeutic Targets and Treatment. Front. Pharmacol. 2022, 13, 780496. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Dong, L.; Lin, X.; Li, J. Relevance of the NLRP3 Inflammasome in the Pathogenesis of Chronic Liver Disease. Front. Immunol. 2017, 8, 1728. [Google Scholar] [CrossRef] [PubMed]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int. J. Mol. Sci. 2019, 20, 3328. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho Ribeiro, M.; Iracheta-Vellve, A.; Babuta, M.; Calenda, C.D.; Copeland, C.; Zhuang, Y.; Lowe, P.P.; Hawryluk, D.; Catalano, D.; Cho, Y.; et al. Alcohol-induced extracellular ASC specks perpetuate liver inflammation and damage in alcohol-associated hepatitis even after alcohol cessation. Hepatology 2023, 78, 225–242. [Google Scholar] [CrossRef]
- Robinson, M.W.; Harmon, C.; O’Farrelly, C. Liver immunology and its role in inflammation and homeostasis. Cell. Mol. Immunol. 2016, 13, 267–276. [Google Scholar] [CrossRef]
- Zhou, Z.; Xu, M.J.; Gao, B. Hepatocytes: A key cell type for innate immunity. Cell. Mol. Immunol. 2016, 13, 301–315. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, M.J.; Radic, M. Neutrophil extracellular traps: Double-edged swords of innate immunity. J. Immunol. 2012, 189, 2689–2695. [Google Scholar] [CrossRef] [PubMed]
- Bukong, T.N.; Cho, Y.; Iracheta-Vellve, A.; Saha, B.; Lowe, P.; Adejumo, A.; Furi, I.; Ambade, A.; Gyongyosi, B.; Catalano, D.; et al. Abnormal neutrophil traps and impaired efferocytosis contribute to liver injury and sepsis severity after binge alcohol use. J. Hepatol. 2018, 69, 1145–1154. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Bukong, T.N.; Tornai, D.; Babuta, M.; Vlachos, I.S.; Kanata, E.; Catalano, D.; Szabo, G. Neutrophil extracellular traps contribute to liver damage and increase defective low-density neutrophils in alcohol-associated hepatitis. J. Hepatol. 2023, 78, 28–44. [Google Scholar] [CrossRef]
- Babuta, M.; Morel, C.; Ribeiro, M.d.C.; Calenda, C.; Ortega-Ribera, M.; Nagesh, P.T.; Copeland, C.; Zhuang, Y.; Wang, Y.; Cho, Y.; et al. Neutrophil Extracellular Traps activate hepatic stellate cells and monocytes via NLRP3 sensing in alcohol-induced acceleration of MASH fibrosis. Gut 2024. Online ahead of print. May 22:gutjnl-2023-331447. [Google Scholar] [CrossRef] [PubMed]
- Chang, B.; Xu, M.; Zhou, Z.; Cai, Y.; Li, M.; Wang, W.; Feng, D.; Bertola, A.; Wang, H.; Kunos, G.; et al. Short- or long-term high-fat diet feeding plus acute ethanol binge synergistically induce acute liver injury in mice: An important role for CXCL1. Hepatology 2015, 62, 1070–1085. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Sun, S.; Xia, S.; Yang, L.; Li, X.; Qi, L. Short term high fat diet challenge promotes alternative macrophage polarization in adipose tissue via natural killer T cells and interleukin-4. J. Biol. Chem. 2012, 287, 24378–24386. [Google Scholar] [CrossRef] [PubMed]
- Wiedemann, M.S.F.; Wueest, S.; Item, F.; Schoenle, E.J.; Konrad, D. Adipose tissue inflammation contributes to short-term high-fat diet-induced hepatic insulin resistance. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E388–E395. [Google Scholar] [CrossRef]
- Lee, Y.S.; Li, P.; Huh, J.Y.; Hwang, I.J.; Lu, M.; Kim, J.I.; Ham, M.; Talukdar, S.; Chen, A.; Lu, W.J.; et al. Inflammation is necessary for long-term but not short-term high-fat diet-induced insulin resistance. Diabetes 2011, 60, 2474–2483. [Google Scholar] [CrossRef]
- Ganz, M.; Csak, T.; Szabo, G. High fat diet feeding results in gender specific steatohepatitis and inflammasome activation. World J. Gastroenterol. 2014, 20, 8525–8534. [Google Scholar] [CrossRef]
- Ganz, M.; Bukong, T.N.; Csak, T.; Saha, B.; Park, J.-K.; Ambade, A.; Kodys, K.; Szabo, G. Progression of non-alcoholic steatosis to steatohepatitis and fibrosis parallels cumulative accumulation of danger signals that promote inflammation and liver tumors in a high fat-cholesterol-sugar diet model in mice. J. Transl. Med. 2015, 13, 193. [Google Scholar] [CrossRef] [PubMed]
- Omary, M.B.; E Cohen, D.; El-Omar, E.; Jalan, R.; Low, M.; Nathanson, M.; Peek, R.; Turner, J. Not All Mice Are the Same: Standardization of Animal Research Data Presentation. Gut 2016, 65, 894–895. [Google Scholar] [CrossRef]
- Swamydas, M.; Lionakis, M.S. Isolation, purification and labeling of mouse bone marrow neutrophils for functional studies and adoptive transfer experiments. J. Vis. Exp. 2013, 10, e50586. [Google Scholar] [CrossRef] [PubMed]
- Babuta, M.; Furi, I.; Bala, S.; Bukong, T.N.; Lowe, P.; Catalano, D.; Calenda, C.; Kodys, K.; Szabo, G. Dysregulated Autophagy and Lysosome Function Are Linked to Exosome Production by Micro-RNA 155 in Alcoholic Liver Disease. Hepatology 2019, 70, 2123–2141. [Google Scholar] [CrossRef]
- Phillips, B.E.; Lantier, L.; Engman, C.; Garciafigueroa, Y.; Singhi, A.; Trucco, M.; Mantzoros, C.; Wasserman, D.; Giannoukakis, N. Improvement in insulin sensitivity and prevention of high fat diet-induced liver pathology using a CXCR2 antagonist. Cardiovasc. Diabetol. 2022, 21, 130. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sun, K.; Zhou, Y.; Wang, H.; Zhou, Y.; Liu, S.; Nie, Y.; Li, Y. NLRP3 inflammasome inhibitor CY-09 reduces hepatic steatosis in experimental NAFLD mice. Biochem. Biophys. Res. Commun. 2021, 534, 734–739. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Liu, Y.; Li, Q.; Wang, F. Inhibition of hepatic NLRP3 inflammasome ameliorates non-alcoholic steatohepatitis/hepatitis B—Induced hepatic injury. Clin. Res. Hepatol. Gastroenterol. 2023, 47, 102056. [Google Scholar] [CrossRef]
- Cho, Y.; Szabo, G. Two Faces of Neutrophils in Liver Disease Development and Progression. Hepatology 2021, 74, 503–512. [Google Scholar] [CrossRef]
- Aroca-Crevillen, A.; Vicanolo, T.; Ovadia, S.; Hidalgo, A. Neutrophils in Physiology and Pathology. Annu. Rev. Pathol. 2024, 19, 227–259. [Google Scholar] [CrossRef]
- Petrasek, J.; Iracheta-Vellve, A.; Saha, B.; Satishchandran, A.; Kodys, K.; A Fitzgerald, K.; A Kurt-Jones, E.; Szabo, G. Metabolic danger signals, uric acid and ATP, mediate inflammatory cross-talk between hepatocytes and immune cells in alcoholic liver disease. J. Leukoc. Biol. 2015, 98, 249–256. [Google Scholar] [CrossRef]
- Carpino, G.; Del Ben, M.; Pastori, D.; Carnevale, R.; Baratta, F.; Overi, D.; Francis, H.; Cardinale, V.; Onori, P.; Safarikia, S.; et al. Increased Liver Localization of Lipopolysaccharides in Human and Experimental NAFLD. Hepatology 2020, 72, 470–485. [Google Scholar] [CrossRef] [PubMed]
- Männistö, V.; Färkkilä, M.; Pussinen, P.; Jula, A.; Männistö, S.; Lundqvist, A.; Valsta, L.; Salomaa, V.; Perola, M.; Åberg, F. Serum lipopolysaccharides predict advanced liver disease in the general population. JHEP Rep. 2019, 1, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Gaul, S.; Leszczynska, A.; Alegre, F.; Kaufmann, B.; Johnson, C.D.; Adams, L.A.; Wree, A.; Damm, G.; Seehofer, D.; Calvente, C.J.; et al. Hepatocyte pyroptosis and release of inflammasome particles induce stellate cell activation and liver fibrosis. J. Hepatol. 2021, 74, 156–167. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Shirasuna, K.; Kimura, H.; Usui, F.; Kawashima, A.; Karasawa, T.; Tago, K.; Dezaki, K.; Nishimura, S.; Sagara, J.; et al. NLRP3 regulates neutrophil functions and contributes to hepatic ischemia-reperfusion injury independently of inflammasomes. J. Immunol. 2014, 192, 4342–4351. [Google Scholar] [CrossRef] [PubMed]
- Burke, S.J.; Lu, D.; Sparer, T.E.; Masi, T.; Goff, M.R.; Karlstad, M.D.; Collier, J.J. NF-kappaB and STAT1 control CXCL1 and CXCL2 gene transcription. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E131–E149. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Sequence (5′-3′) | Reverse Sequence (5′-3′) |
|---|---|---|
| Casp1 | GGCACATTTCCAGGACTGACTG | GCAAGACGTGTACGAGTGGTTG |
| Ccl2 | TCTTGGTTCCCTGGCGTACTCT | GTGAGTGTCACTCTCCAGTTTGC |
| Cxcl1 | CAAGGCTGGTCCATGCTCC | TGCTATCACTTCCTTTCTGTTGC |
| Cxcl2 | CCAACCACCAGGCTACAGG | GCGTCACACTCAAGCTCTG |
| Cxcr1 | TCTGGACTAATCCTGAGGGTG | GCCTGTTGGTTATTGGAACTCTC |
| Cxcr2 | TGTCTGGGCTGCATCTAAAGT | AGGTAACCTCCTTCACGTATGAG |
| Il18 | CCTACTTCAGCATCCTCTACTGG | AGGGTTTCTTGAGAAGGGGAC |
| Il1b | TGGACCTTCCAGGATGAGGACA | GTTCATCTCGGAGCCTGTAGTG |
| Itgam | ATGGACGCTGATGGCAATACC | TCCCCATTCACGTCTCCCA |
| Lcn2 | TGGCCCTGAGTGTCATGTG | CTCTTGTAGCTCATAGATGGTGC |
| Nlrp3 | TCACAACTCGCCCAAGGAGGAA | AAGAGACCACGGCAGAAGCTAG |
| Pycard | CGGAAAGTGGAATCCTTGCAGG | AGCAGTGAGGTCAGGCTTGGAA |
| Tnf | CTGGATGTCAATCAACAATGGGA | ACTAGGGTGTGAGTGTTTTCTGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Babuta, M.; Nagesh, P.T.; Datta, A.A.; Remotti, V.; Zhuang, Y.; Mehta, J.; Lami, F.; Wang, Y.; Szabo, G. Combined Insults of a MASH Diet and Alcohol Binges Activate Intercellular Communication and Neutrophil Recruitment via the NLRP3-IL-1β Axis in the Liver. Cells 2024, 13, 960. https://doi.org/10.3390/cells13110960
Babuta M, Nagesh PT, Datta AA, Remotti V, Zhuang Y, Mehta J, Lami F, Wang Y, Szabo G. Combined Insults of a MASH Diet and Alcohol Binges Activate Intercellular Communication and Neutrophil Recruitment via the NLRP3-IL-1β Axis in the Liver. Cells. 2024; 13(11):960. https://doi.org/10.3390/cells13110960
Chicago/Turabian StyleBabuta, Mrigya, Prashanth Thevkar Nagesh, Aditi Ashish Datta, Victoria Remotti, Yuan Zhuang, Jeeval Mehta, Francesca Lami, Yanbo Wang, and Gyongyi Szabo. 2024. "Combined Insults of a MASH Diet and Alcohol Binges Activate Intercellular Communication and Neutrophil Recruitment via the NLRP3-IL-1β Axis in the Liver" Cells 13, no. 11: 960. https://doi.org/10.3390/cells13110960
APA StyleBabuta, M., Nagesh, P. T., Datta, A. A., Remotti, V., Zhuang, Y., Mehta, J., Lami, F., Wang, Y., & Szabo, G. (2024). Combined Insults of a MASH Diet and Alcohol Binges Activate Intercellular Communication and Neutrophil Recruitment via the NLRP3-IL-1β Axis in the Liver. Cells, 13(11), 960. https://doi.org/10.3390/cells13110960