
Induced Pluripotent Stem Cells and CRISPR-Cas9 Innovations for Treating Alpha-1 Antitrypsin Deficiency and Glycogen Storage Diseases


Abstract
:1. Introduction
2. Overview of A1AD and GSDI
2.1. Genetic Basis and Clinical Manifestations of A1AD
2.2. Genetic Basis and Clinical Manifestations of GSDI
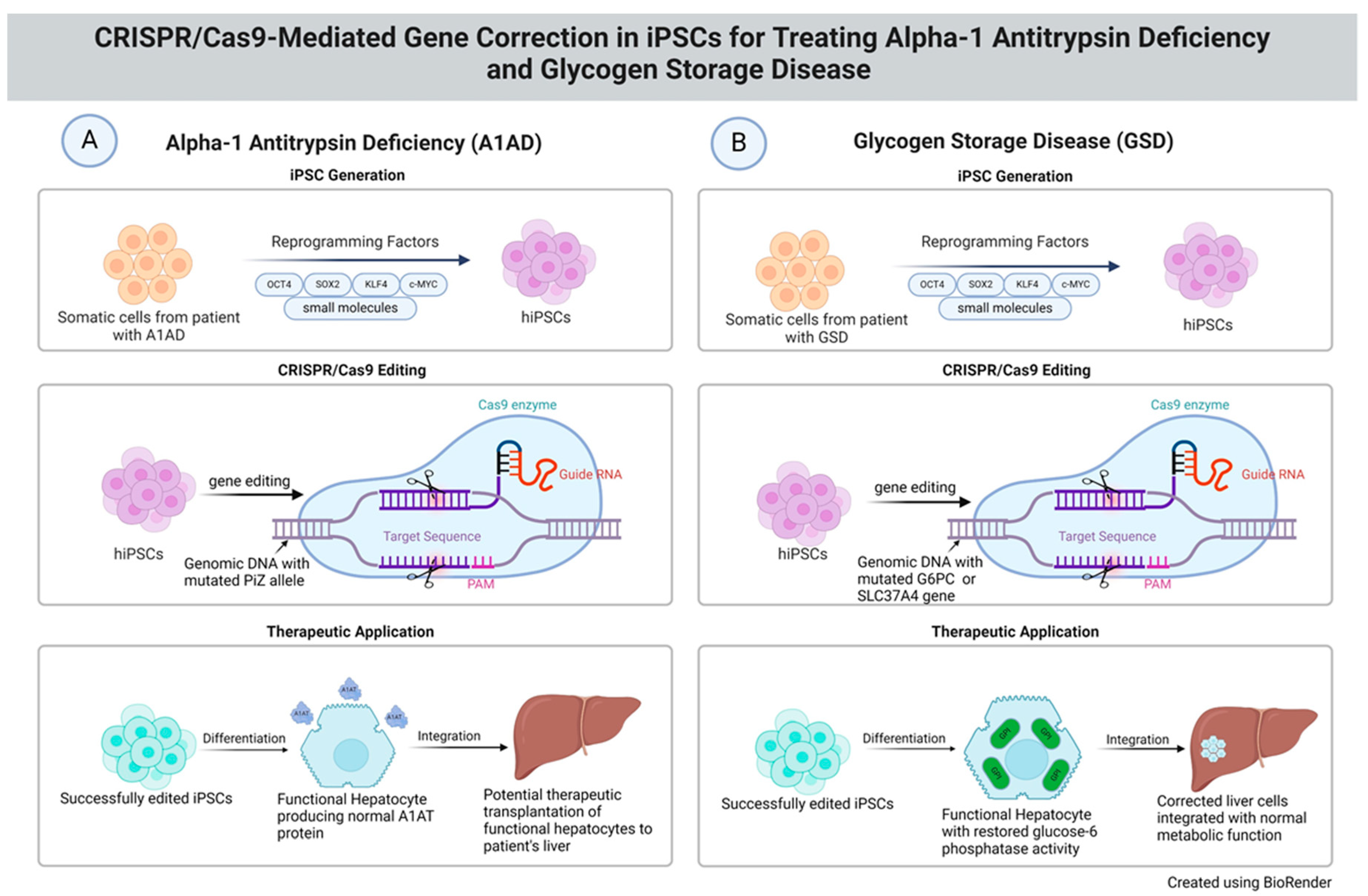
3. iPSCs for Modeling A1AD and GSDI
3.1. Discoveries about A1AD Disease Mechanisms and Potential Therapies
3.2. Advancements in Understanding Metabolic Impacts and Therapeutic Targets for GSDI

{kind=link}
| Aspect | A1AD Achievements | A1AD Challenges | GSDI Achievements | GSDI Challenges | References |
|---|---|---|---|---|---|
| Disease Modeling | Developed iPSC-derived hepatocyte models to study A1AT accumulation and liver fibrosis. | Difficulty in fully replicating the liver environment and its complex interactions in vitro. | Created detailed models of liver and kidney cells to study metabolic dysfunctions and glycogen accumulation. | Challenges in replicating exact physiological conditions of glucose metabolism in vitro. | [16,17,20,28] |
| Gene Therapy | Utilized CRISPR-Cas9 to correct SERPINA1 gene mutations directly in iPSCs. | Ensuring long-term stability and integration of corrected genes without off-target effects. | Tested gene-editing tools to correct genetic defects in the G6PC and SLC37A4 genes. | Managing off-target effects and ensuring precise gene correction in all affected cells. | [9,36,37,38,39,40] |
| Drug Testing | iPSC models used for screening potential therapeutic compounds that can alleviate liver fibrosis. | Variability in drug responses due to patient-specific iPSC differences. | Enabled preclinical testing of new pharmacological agents to manage glycogen storage and enhance glucose release. | Difficulty in predicting clinical efficacy based on iPSC-derived model results. | [1,11,16,18] |
| Pathophysiological Insights | Revealed mechanisms of inflammatory response and oxidative stress due to A1AT deficiency. | Requires more comprehensive models that include immune and other systemic interactions. | Provided insights into abnormal glucose-6-phosphatase activity and its systemic effects. | Requires deeper understanding of long-term disease progression and secondary complications. | [8,9,41,42] |
| Therapeutic Development | Opened avenues for developing targeted therapies that can be personalized based on the patient’s genetic profile. | Development and regulatory challenges in transitioning from iPSC models to clinical treatments. | Facilitated exploration of enzyme replacement and other supportive therapies in a controlled setting. | Translating laboratory successes into viable clinical therapies remains slow and complex. | [13,18,39,43,44] |
4. CRISPR-Cas9 Gene-Editing Technology for A1AD Disease Modeling
5. CRISPR-Cas9 Gene-Editing Technology for Modeling GSDI
6. Technical Challenges in Disease Modeling and Gene Therapy
7. Ethical Dilemmas and Regulatory Landscape of Using iPSCs and CRISPR-Cas9
8. Future Directions
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cota-Coronado, A.; Ramírez-Rodríguez, P.B.; Padilla-Camberos, E.; Flores-Fernández, J.M.; Ávila-Gónzalez, D.; Diaz-Martinez, N.E. Implications of human induced pluripotent stem cells in metabolic disorders: From drug discovery toward precision medicine. Drug Discov. Today 2019, 24, 334–341. [Google Scholar] [CrossRef]
- Escribá, R.; Ferrer-Lorente, R.; Raya, Á. Inborn errors of metabolism: Lessons from iPSC models. Rev. Endocr. Metab. Disord. 2021, 22, 1189–1200. [Google Scholar] [CrossRef]
- Lopes, C.; Tang, Y.; Anjo, S.I.; Manadas, B.; Onofre, I.; de Almeida, L.P.; Daley, G.Q.; Schlaeger, T.M.; Rego, A.C.C. Mitochondrial and redox modifications in huntington disease induced pluripotent stem cells rescued by CRISPR/Cas9 CAGs targeting. Front. Cell Dev. Biol. 2020, 8, 576592. [Google Scholar] [CrossRef]
- Maldonado, R.; Jalil, S.; Keskinen, T.; Nieminen, A.I.; Hyvönen, M.E.; Lapatto, R.; Wartiovaara, K. CRISPR correction of the Finnish ornithine delta-aminotransferase mutation restores metabolic homeostasis in iPSC from patients with gyrate atrophy. Mol. Genet. Metab. Rep. 2022, 31, 100863. [Google Scholar] [CrossRef]
- Song, H.-Y.; Yang, Y.-P.; Chien, Y.; Lai, W.-Y.; Lin, Y.-Y.; Chou, S.-J.; Wang, M.-L.; Wang, C.-Y.; Leu, H.-B.; Yu, W.-C. Reversal of the inflammatory responses in Fabry patient IPSC-derived cardiovascular endothelial cells by CRISPR/Cas9-corrected mutation. Int. J. Mol. Sci. 2021, 22, 2381. [Google Scholar] [CrossRef]
- Li, T.; Yang, Y.; Qi, H.; Cui, W.; Zhang, L.; Fu, X.; He, X.; Liu, M.; Li, P.-F.; Yu, T. CRISPR/Cas9 therapeutics: Progress and prospects. Signal Transduct. Target. Ther. 2023, 8, 36. [Google Scholar] [CrossRef]
- Karatas, E.; Di-Tommaso, S.; Dugot-Senant, N.; Lachaux, A.; Bouchecareilh, M. Overview of alpha-1 antitrypsin deficiency-mediated liver disease. Eur. Med. J. 2019, 7, 65–79. [Google Scholar] [CrossRef]
- Bouchecareilh, M. Alpha-1 antitrypsin deficiency-mediated liver toxicity: Why do some patients do poorly? What do we know so far? Chronic Obstr. Pulm. Dis. J. COPD Found. 2020, 7, 172. [Google Scholar] [CrossRef]
- Kishnani, P.S.; Sun, B.; Koeberl, D.D. Gene therapy for glycogen storage diseases. Hum. Mol. Genet. 2019, 28, R31–R41. [Google Scholar] [CrossRef]
- Adlat, S.; Salgado, A.M.V.; Lee, M.; Yin, D.; Wangensteen, K.J. Emerging and potential use of CRISPR in human liver disease. Hepatology, 2023; Online ahead of print. [Google Scholar]
- Corbett, J.L.; Duncan, S.A. iPSC-derived hepatocytes as a platform for disease modeling and drug discovery. Front. Med. 2019, 6, 265. [Google Scholar] [CrossRef]
- Heslop, J.A.; Duncan, S.A. The use of human pluripotent stem cells for modeling liver development and disease. Hepatology 2019, 69, 1306–1316. [Google Scholar] [CrossRef]
- Rowe, R.G.; Daley, G.Q. Induced pluripotent stem cells in disease modelling and drug discovery. Nat. Rev. Genet. 2019, 20, 377–388. [Google Scholar] [CrossRef]
- Yefroyev, D.A.; Jin, S. Induced pluripotent stem cells for treatment of Alzheimer’s and Parkinson’s diseases. Biomedicines 2022, 10, 208. [Google Scholar] [CrossRef]
- Ogi, D.A.; Jin, S. Transcriptome-Powered Pluripotent Stem Cell Differentiation for Regenerative Medicine. Cells 2023, 12, 1442. [Google Scholar] [CrossRef]
- Blaszkiewicz, J.; Duncan, S.A. Advancements in disease modeling and drug discovery using iPSC-derived hepatocyte-like cells. Genes 2022, 13, 573. [Google Scholar] [CrossRef]
- Caiazza, C.; Parisi, S.; Caiazzo, M. Liver organoids: Updates on disease modeling and biomedical applications. Biology 2021, 10, 835. [Google Scholar] [CrossRef]
- Gümüş, E.; Özen, H. Glycogen storage diseases: An update. World J. Gastroenterol. 2023, 29, 3932. [Google Scholar] [CrossRef]
- Rossiaud, L.; Pellier, E.; Benabides, M.; Nissan, X.; Ronzitti, G.; Hoch, L. Generation of three induced pluripotent stem cell lines from patients with glycogen storage disease type III. Stem Cell Res. 2023, 72, 103214. [Google Scholar] [CrossRef]
- Ignatius Irudayam, J.; Contreras, D.; Sivasubramaniam, S.; Arumugaswami, V. Modeling Liver Diseases Using Induced Pluripotent Stem Cell (iPSC)-Derived Hepatocytes. J. Stem Cell Res. Ther. 2014, 4, 2. [Google Scholar]
- Takahashi, K.; Yamanaka, S. A decade of transcription factor-mediated reprogramming to pluripotency. Nat. Rev. Mol. Cell Biol. 2016, 17, 183–193. [Google Scholar] [CrossRef]
- Hou, P.; Li, Y.; Zhang, X.; Liu, C.; Guan, J.; Li, H.; Zhao, T.; Ye, J.; Yang, W.; Liu, K. Pluripotent stem cells induced from mouse somatic cells by small-molecule compounds. Science 2013, 341, 651–654. [Google Scholar] [CrossRef]
- Rais, Y.; Zviran, A.; Geula, S.; Gafni, O.; Chomsky, E.; Viukov, S.; Mansour, A.A.; Caspi, I.; Krupalnik, V.; Zerbib, M. Deterministic direct reprogramming of somatic cells to pluripotency. Nature 2013, 502, 65–70. [Google Scholar] [CrossRef]
- Shi, Y.; Do, J.T.; Desponts, C.; Hahm, H.S.; Schöler, H.R.; Ding, S. A combined chemical and genetic approach for the generation of induced pluripotent stem cells. Cell Stem Cell 2008, 2, 525–528. [Google Scholar] [CrossRef]
- Li, C.; Wang, L.; Cseke, L.J.; Vasconcelos, F.; Huguet-Tapia, J.C.; Gassmann, W.; Pauwels, L.; White, F.F.; Dong, H.; Yang, B. Efficient CRISPR-Cas9 based cytosine base editors for phytopathogenic bacteria. Commun. Biol. 2023, 6, 56. [Google Scholar] [CrossRef]
- Gaudelli, N.M.; Komor, A.C.; Rees, H.A.; Packer, M.S.; Badran, A.H.; Bryson, D.I.; Liu, D.R. Programmable base editing of A• T to G• C in genomic DNA without DNA cleavage. Nature 2017, 551, 464–471. [Google Scholar] [CrossRef]
- Werder, R.B.; Kaserman, J.E.; Packer, M.S.; Lindstrom-Vautrin, J.; Villacorta-Martin, C.; Young, L.E.; Aratyn-Schaus, Y.; Gregoire, F.; Wilson, A.A. Adenine base editing reduces misfolded protein accumulation and toxicity in alpha-1 antitrypsin deficient patient iPSC-hepatocytes. Mol. Ther. 2021, 29, 3219–3229. [Google Scholar] [CrossRef]
- Tafaleng, E.N.; Chakraborty, S.; Han, B.; Hale, P.; Wu, W.; Soto-Gutierrez, A.; Feghali-Bostwick, C.A.; Wilson, A.A.; Kotton, D.N.; Nagaya, M. Induced pluripotent stem cells model personalized variations in liver disease resulting from α1-antitrypsin deficiency. Hepatology 2015, 62, 147–157. [Google Scholar] [CrossRef]
- Kaserman, J.E.; Wilson, A.A. Patient-derived induced pluripotent stem cells for alpha-1 antitrypsin deficiency disease modeling and therapeutic discovery. Chronic Obstr. Pulm. Dis. J. COPD Found. 2018, 5, 258. [Google Scholar] [CrossRef]
- Wilson, A.A.; Ying, L.; Liesa, M.; Segeritz, C.-P.; Mills, J.A.; Shen, S.S.; Jean, J.; Lonza, G.C.; Liberti, D.C.; Lang, A.H. Emergence of a stage-dependent human liver disease signature with directed differentiation of alpha-1 antitrypsin-deficient iPS cells. Stem Cell Rep. 2015, 4, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Gieseck III, R.L.; Colquhoun, J.; Hannan, N.R. Disease modeling using human induced pluripotent stem cells: Lessons from the liver. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2015, 1851, 76–89. [Google Scholar] [CrossRef] [PubMed]
- Katagami, Y.; Kondo, T.; Suga, M.; Yada, Y.; Imamura, K.; Shibukawa, R.; Sagara, Y.; Okanishi, Y.; Tsukita, K.; Hirayama, K. Generation of a human induced pluripotent stem cell line, BRCi009-A, derived from a patient with glycogen storage disease type 1a. Stem Cell Res. 2020, 49, 102095. [Google Scholar] [CrossRef] [PubMed]
- Satoh, D.; Maeda, T.; Ito, T.; Nakajima, Y.; Ohte, M.; Ukai, A.; Nakamura, K.; Enosawa, S.; Toyota, M.; Miyagawa, Y. Establishment and directed differentiation of induced pluripotent stem cells from glycogen storage disease type I b patient. Genes Cells 2013, 18, 1053–1069. [Google Scholar] [CrossRef] [PubMed]
- Rossiaud, L.; Fragner, P.; Barbon, E.; Gardin, A.; Benabides, M.; Pellier, E.; Cosette, J.; El Kassar, L.; Giraud-Triboult, K.; Nissan, X. Pathological modeling of glycogen storage disease type III with CRISPR/Cas9 edited human pluripotent stem cells. Front. Cell Dev. Biol. 2023, 11, 1163427. [Google Scholar] [CrossRef] [PubMed]
- Naito, C.; Kosar, K.; Kishimoto, E.; Pena, L.; Huang, Y.; Hao, K.; Bernieh, A.; Kasten, J.; Villa, C.; Kishnani, P. Induced pluripotent stem cell (iPSC) modeling validates reduced GBE1 enzyme activity due to a novel variant, p. Ile694Asn, found in a patient with suspected glycogen storage disease IV. Mol. Genet. Metab. Rep. 2024, 39, 101069. [Google Scholar] [PubMed]
- Koeberl, D.D.; Koch, R.L.; Lim, J.A.; Brooks, E.D.; Arnson, B.D.; Sun, B.; Kishnani, P.S. Gene therapy for glycogen storage diseases. J. Inherit. Metab. Dis. 2024, 47, 93–118. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, M.; Masi, G.; Palù, G. Genome editing technologies to treat rare liver diseases. Transl. Gastroenterol. Hepatol. 2020, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Arnaoutova, I.; Zhang, L.; Chen, H.-D.; Mansfield, B.C.; Chou, J.Y. Correction of metabolic abnormalities in a mouse model of glycogen storage disease type Ia by CRISPR/Cas9-based gene editing. Mol. Ther. 2021, 29, 1602–1610. [Google Scholar] [CrossRef] [PubMed]
- Chou, J.Y.; Mansfield, B.C. Gene therapy and genome editing for type I glycogen storage diseases. Front. Mol. Med. 2023, 3, 1167091. [Google Scholar] [CrossRef]
- Hannoun, Z.; Steichen, C.; Dianat, N.; Weber, A.; Dubart-Kupperschmitt, A. The potential of induced pluripotent stem cell derived hepatocytes. J. Hepatol. 2016, 65, 182–199. [Google Scholar] [CrossRef]
- Nuciforo, S.; Heim, M.H. Organoids to model liver disease. JHEP Rep. 2021, 3, 100198. [Google Scholar] [CrossRef]
- Rahaghi, F.F. Alpha-1 antitrypsin deficiency research and emerging treatment strategies: What’s down the road? Ther. Adv. Chronic Dis. 2021, 12, 20406223211014025. [Google Scholar] [CrossRef] [PubMed]
- Zu, H.; Gao, D. Non-viral vectors in gene therapy: Recent development, challenges, and prospects. AAPS J. 2021, 23, 78. [Google Scholar] [CrossRef] [PubMed]
- Doudna, J.A. The promise and challenge of therapeutic genome editing. Nature 2020, 578, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Packer, M.S.; Chowdhary, V.; Lung, G.; Cheng, L.-I.; Aratyn-Schaus, Y.; Leboeuf, D.; Smith, S.; Shah, A.; Chen, D.; Zieger, M. Evaluation of cytosine base editing and adenine base editing as a potential treatment for alpha-1 antitrypsin deficiency. Mol. Ther. 2022, 30, 1396–1406. [Google Scholar] [CrossRef] [PubMed]
- Seixas, S.; Marques, P.I. Known mutations at the cause of alpha-1 antitrypsin deficiency an updated overview of SERPINA1 variation spectrum. Appl. Clin. Genet. 2021, 14, 173–194. [Google Scholar] [CrossRef]
- Smith, C.; Abalde-Atristain, L.; He, C.; Brodsky, B.R.; Braunstein, E.M.; Chaudhari, P.; Jang, Y.-Y.; Cheng, L.; Ye, Z. Efficient and allele-specific genome editing of disease loci in human iPSCs. Mol. Ther. 2015, 23, 570–577. [Google Scholar] [CrossRef] [PubMed]
- Skakic, A.; Andjelkovic, M.; Tosic, N.; Klaassen, K.; Djordjevic, M.; Pavlovic, S.; Stojiljkovic, M. CRISPR/Cas9 genome editing of SLC37A4 gene elucidates the role of molecular markers of endoplasmic reticulum stress and apoptosis in renal involvement in glycogen storage disease type Ib. Gene 2019, 703, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Rutten, M.G.; Derks, T.G.; Huijkman, N.C.; Bos, T.; Kloosterhuis, N.J.; van de Kolk, K.C.; Wolters, J.C.; Koster, M.H.; Bongiovanni, L.; Thomas, R.E. Modeling Phenotypic Heterogeneity of Glycogen Storage Disease Type 1a Liver Disease in Mice by Somatic CRISPR/CRISPR-associated protein 9–Mediated Gene Editing. Hepatology 2021, 74, 2491–2507. [Google Scholar] [CrossRef] [PubMed]
- Goswami, R.; Subramanian, G.; Silayeva, L.; Newkirk, I.; Doctor, D.; Chawla, K.; Chattopadhyay, S.; Chandra, D.; Chilukuri, N.; Betapudi, V. Gene therapy leaves a vicious cycle. Front. Oncol. 2019, 9, 297. [Google Scholar] [CrossRef]
- Doss, M.; Sachinidis, A. Current challenges of iPSC-based disease modeling and therapeutic implications. Cells 8: 403. 2019.
- Reshetnikov, V.V.; Chirinskaite, A.V.; Sopova, J.V.; Ivanov, R.A.; Leonova, E.I. Translational potential of base-editing tools for gene therapy of monogenic diseases. Front. Bioeng. Biotechnol. 2022, 10, 942440. [Google Scholar] [CrossRef] [PubMed]
- Shahryari, A.; Saghaeian Jazi, M.; Mohammadi, S.; Razavi Nikoo, H.; Nazari, Z.; Hosseini, E.S.; Burtscher, I.; Mowla, S.J.; Lickert, H. Development and clinical translation of approved gene therapy products for genetic disorders. Front. Genet. 2019, 10, 868. [Google Scholar] [CrossRef] [PubMed]
- Khoshandam, M.; Soltaninejad, H.; Mousazadeh, M.; Hamidieh, A.A.; Hosseinkhani, S. Clinical applications of the CRISPR/Cas9 genome-editing system: Delivery options and challenges in precision medicine. Genes Dis. 2024, 11, 268–282. [Google Scholar] [CrossRef] [PubMed]
- Richardson, C.; Kelsh, R.N.; Richardson, R.J. New advances in CRISPR/Cas-mediated precise gene-editing techniques. Dis. Models Mech. 2023, 16, dmm049874. [Google Scholar] [CrossRef] [PubMed]
- Tyumentseva, M.; Tyumentsev, A.; Akimkin, V. CRISPR/Cas9 landscape: Current state and future perspectives. Int. J. Mol. Sci. 2023, 24, 16077. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Yang, J.; Xin, X.; Liu, C.; Li, L.; Mei, X.; Li, M. Merits and challenges of iPSC-derived organoids for clinical applications. Front. Cell Dev. Biol. 2023, 11, 1188905. [Google Scholar] [CrossRef]
| Feature | Alpha-1 Antitrypsin Deficiency (A1AD) | Glycogen Storage Disease Type I (GSDI) |
|---|---|---|
| Genetic Basis | Mutation in the SERPINA1 gene leads to a defective production of alpha-1 antitrypsin. | Mutations in the G6PC (GSDIa) or SLC37A4 (GSDIb) genes affect glucose-6-phosphatase. |
| Pathophysiology | Accumulation of misfolded A1AT protein in the liver, impairing its release into the bloodstream and reducing its protease inhibitor activity. | Deficient activity of glucose-6-phosphatase disrupts glycogenolysis and gluconeogenesis, causing excessive glycogen and fat accumulation in the liver and kidneys. |
| Primary Organ Impact | Liver and lungs. | Liver and kidneys. |
| Clinical Manifestations | Pulmonary disorders such as early-onset emphysema and COPD; liver disease, ranging from mild enzyme elevations to cirrhosis; and hepatocellular carcinoma. | Severe hypoglycemia, hepatomegaly, growth retardation, metabolic acidosis, and progressive renal disease. |
| Common Symptoms | Shortness of breath, wheezing, and liver dysfunction. | Hypoglycemia symptoms (e.g., fatigue, irritability), enlarged liver, stunted growth. |
| Treatment Approaches | Augmentation therapy (infusion of A1AT) and liver transplantation in severe cases. | Dietary management (frequent carbohydrate-rich meals), medications to control metabolic symptoms, liver transplantation in severe cases. |
| Prognosis | Variable: depends on the degree of lung and liver disease. Life expectancy can be near normal with appropriate management. | Chronic and managed conditions; complications like kidney disease can impact life expectancy if not properly managed. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walsh, C.; Jin, S. Induced Pluripotent Stem Cells and CRISPR-Cas9 Innovations for Treating Alpha-1 Antitrypsin Deficiency and Glycogen Storage Diseases. Cells 2024, 13, 1052. https://doi.org/10.3390/cells13121052
Walsh C, Jin S. Induced Pluripotent Stem Cells and CRISPR-Cas9 Innovations for Treating Alpha-1 Antitrypsin Deficiency and Glycogen Storage Diseases. Cells. 2024; 13(12):1052. https://doi.org/10.3390/cells13121052
Chicago/Turabian StyleWalsh, Colin, and Sha Jin. 2024. "Induced Pluripotent Stem Cells and CRISPR-Cas9 Innovations for Treating Alpha-1 Antitrypsin Deficiency and Glycogen Storage Diseases" Cells 13, no. 12: 1052. https://doi.org/10.3390/cells13121052