
ScRNA-Seq Analyses Define the Role of GATA3 in iNKT Cell Effector Lineage Differentiation

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Material and Methods
2.1. Mice
2.2. Single-Cell RNA Sequencing and Data Processing
2.3. FACS Analysis and Antibodies
2.4. Bone Marrow Transfer
2.5. Statistical Analysis
3. Results
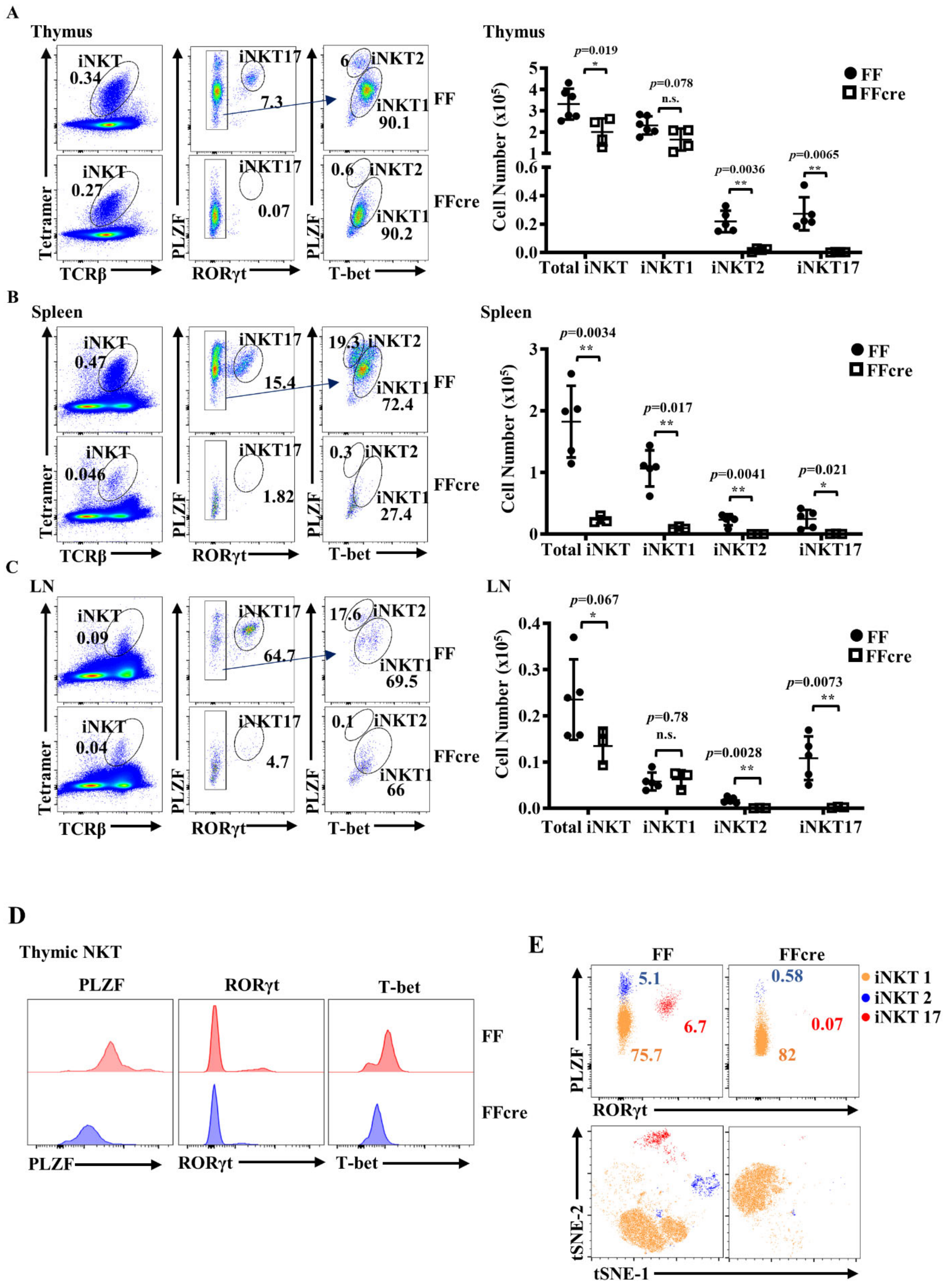
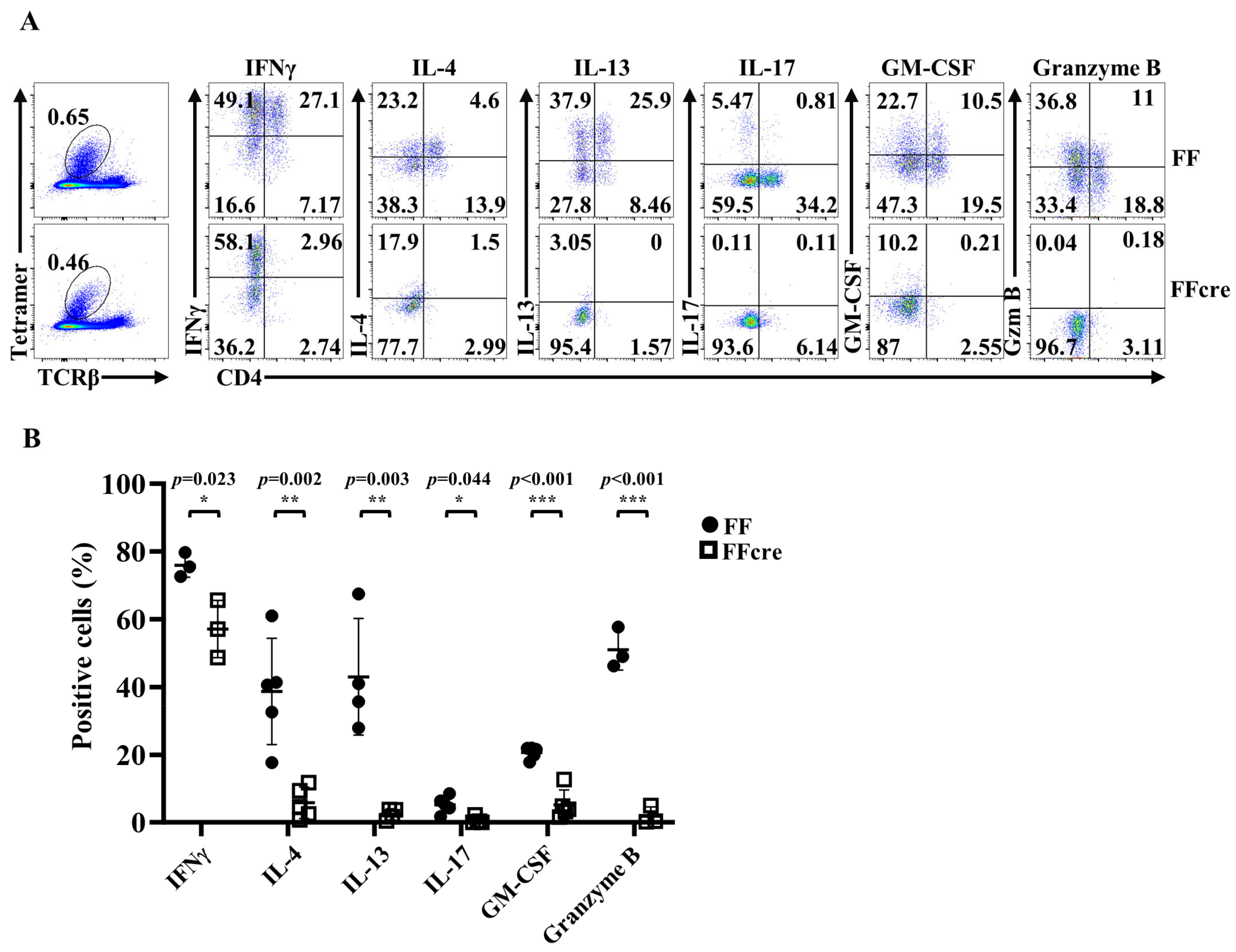
3.1. GATA-3 Deficiency Impairs the Generation of Both iNKT2 and iNKT17 Cells
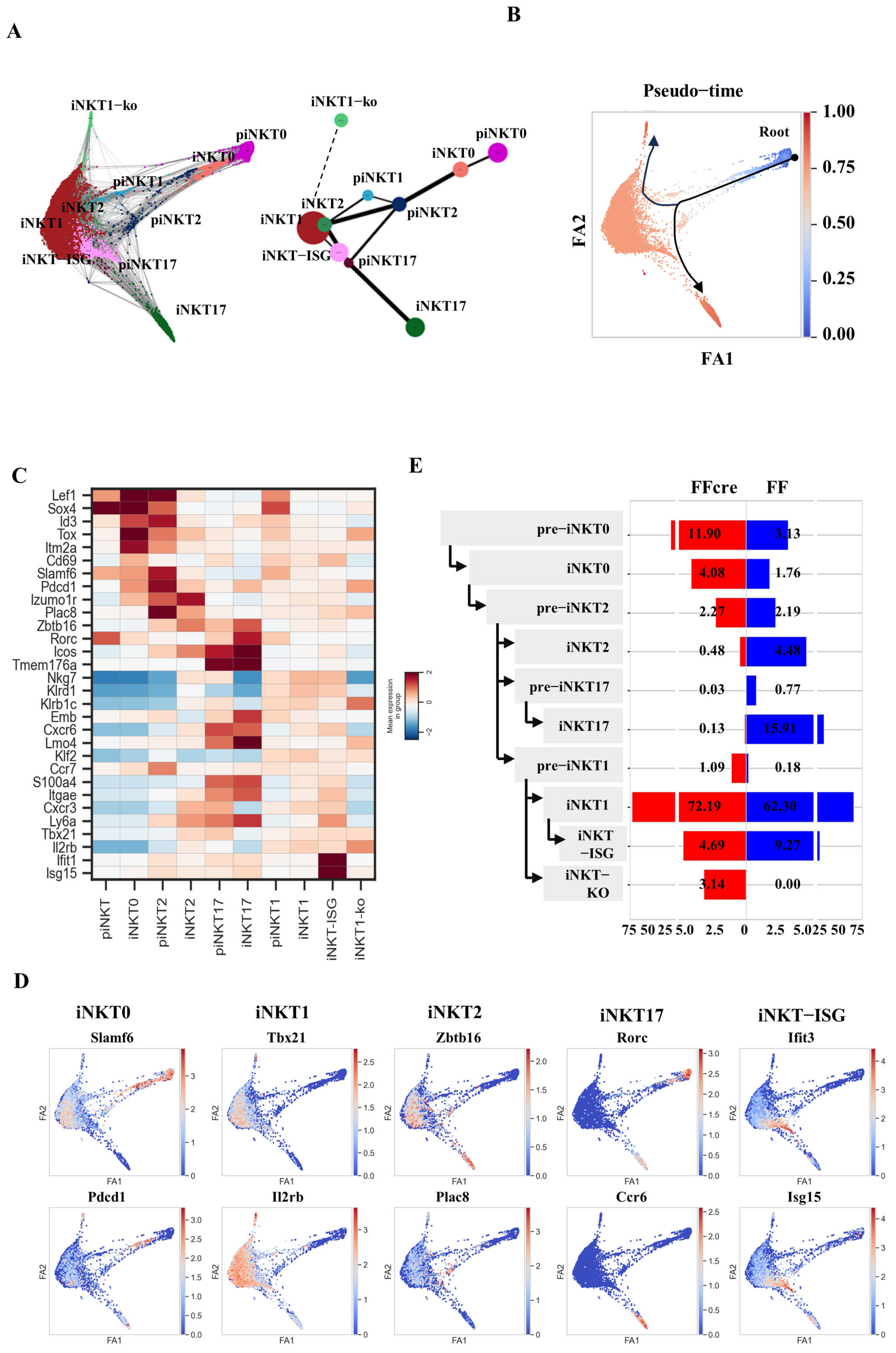
3.2. scRNA-seq Transcriptomic Profiles of the Thymic iNKT Cells from GATA-3 FF and FFcre Mice
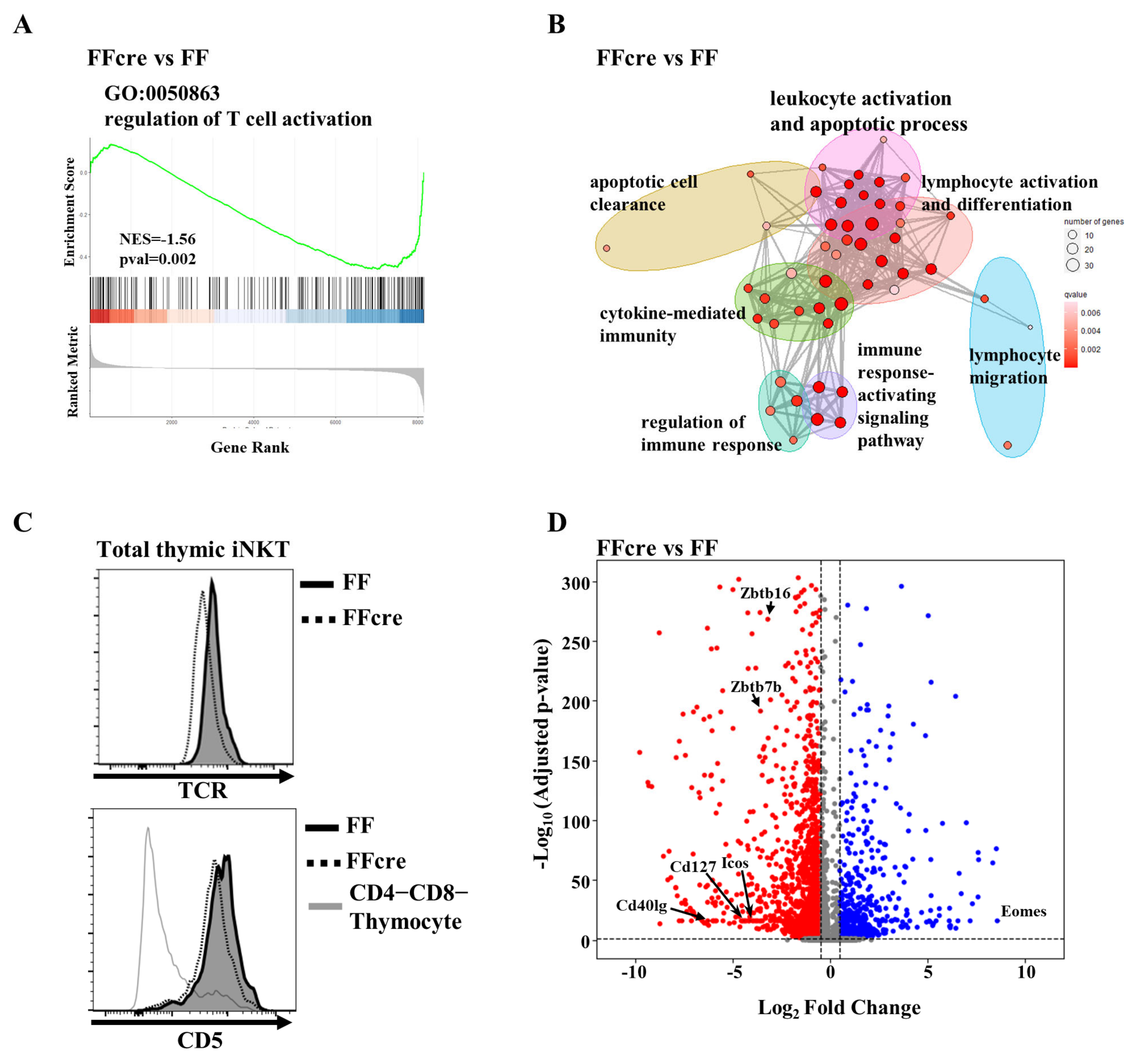
3.3. GATA-3 Controls iNKT Cell Activation Signal
3.4. GATA-3 Regulates Gene Expression in Various Subsets
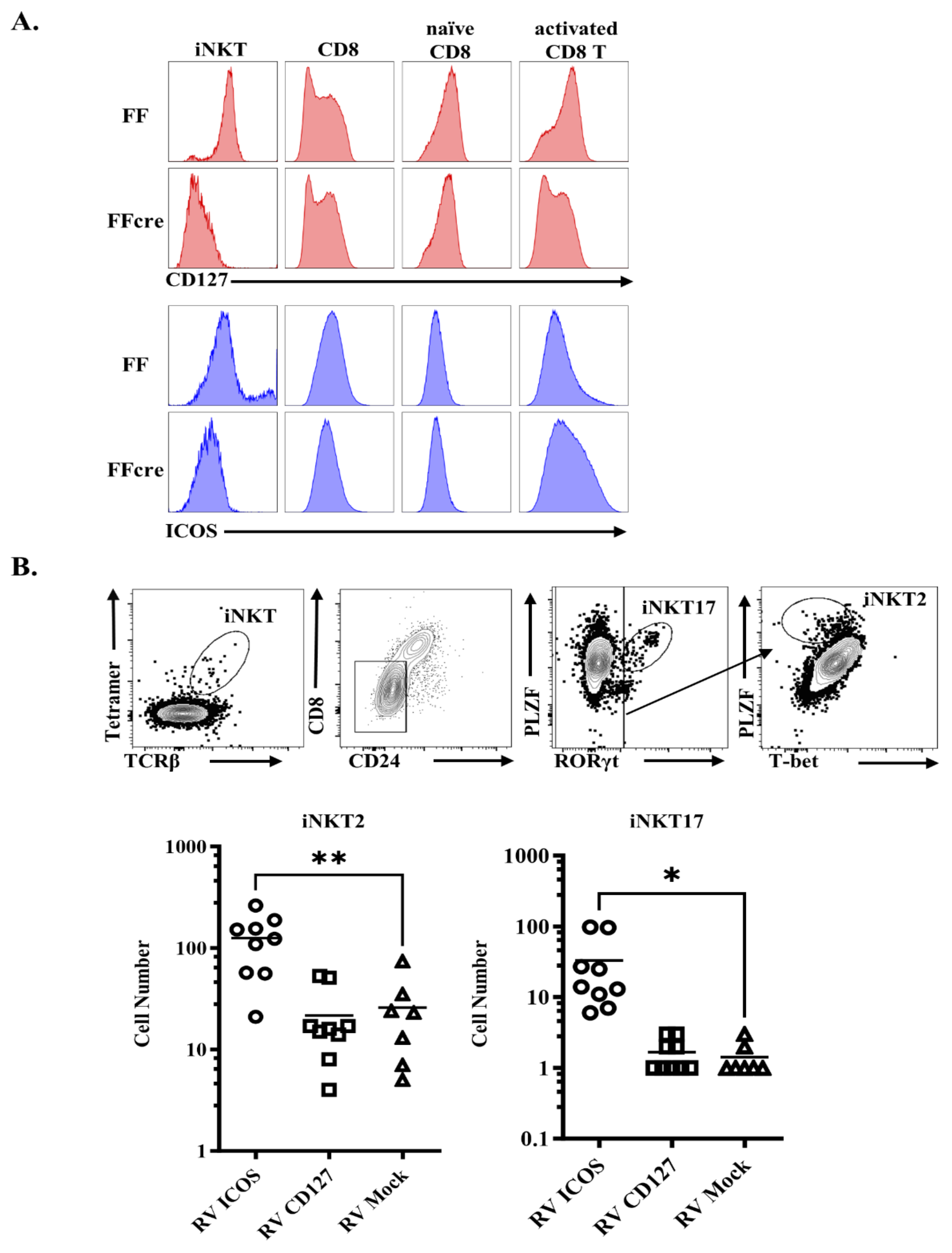
3.5. GATA-3 Affects iNKT2 and iNKT17 Differentiation by Regulating CD127 or ICOS
4. Discussion
5. Limitations of Current Study
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bendelac, A.; Savage, P.B.; Teyton, L. The Biology of Nkt Cells. Annu. Rev. Immunol. 2007, 25, 297–336. [Google Scholar] [CrossRef]
- Taniguchi, M.; Nakayama, T. Recognition and Function of Valpha14 NKT Cells. Semin. Immunol. 2000, 12, 543–550. [Google Scholar] [CrossRef]
- Kawano, T.; Nakayama, T.; Kamada, N.; Kaneko, Y.; Harada, M.; Ogura, N.; Akutsu, Y.; Motohashi, S.; Iizasa, T.; Endo, H.; et al. Antitumor Cytotoxicity Mediated by Ligand-Activated Human V Alpha24 NKT Cells. Cancer Res. 1999, 59, 5102–5105. [Google Scholar]
- Metelitsa, L.S.; Naidenko, O.V.; Kant, A.; Wu, H.W.; Loza, M.J.; Perussia, B.; Kronenberg, M.; Seeger, R.C. Human NKT Cells Mediate Antitumor Cytotoxicity Directly by Recognizing Target Cell CD1d with Bound Ligand or Indirectly by Producing IL-2 to Activate NK Cells. J. Immunol. 2001, 167, 3114–3122. [Google Scholar] [CrossRef]
- Kim, C.H.; Butcher, E.C.; Johnston, B. Distinct Subsets of Human Valpha24-Invariant NKT Cells: Cytokine Responses and Chemokine Receptor Expression. Trends Immunol. 2002, 23, 516–519. [Google Scholar] [CrossRef]
- Gumperz, J.E.; Miyake, S.; Yamamura, T.; Brenner, M.B. Functionally Distinct Subsets of CD1d-Restricted Natural Killer T Cells Revealed by CD1d Tetramer Staining. J. Exp. Med. 2002, 195, 625–636. [Google Scholar] [CrossRef]
- Michel, M.L.; Keller, A.C.; Paget, C.; Fujio, M.; Trottein, F.; Savage, P.B.; Wong, C.H.; Schneider, E.; Dy, M.; Leite-de-Moraes, M.C. Identification of an IL-17-Producing NK1.1(Neg) iNKT Cell Population Involved in Airway Neutrophilia. J. Exp. Med. 2007, 204, 995–1001. [Google Scholar] [CrossRef]
- Goto, M.; Murakawa, M.; Kadoshima-Yamaoka, K.; Tanaka, Y.; Nagahira, K.; Fukuda, Y.; Nishimura, T. Murine NKT Cells Produce Th17 Cytokine Interleukin-22. Cell. Immunol. 2009, 254, 81–84. [Google Scholar] [CrossRef]
- Van Kaer, L.; Parekh, V.V.; Wu, L. Invariant Natural Killer T Cells as Sensors and Managers of Inflammation. Trends Immunol. 2013, 34, 50–58. [Google Scholar] [CrossRef]
- Berzins, S.P.; Ritchie, D.S. Natural Killer T Cells: Drivers or Passengers in Preventing Human Disease? Nat. Rev. Immunol. 2014, 14, 640–646. [Google Scholar] [CrossRef]
- Terashima, A.; Watarai, H.; Inoue, S.; Sekine, E.; Nakagawa, R.; Hase, K.; Iwamura, C.; Nakajima, H.; Nakayama, T.; Taniguchi, M. A Novel Subset of Mouse NKT Cells Bearing the IL-17 Receptor B Responds to IL-25 and Contributes to Airway Hyperreactivity. J. Exp. Med. 2008, 205, 2727–2733. [Google Scholar] [CrossRef]
- Osman, Y.; Kawamura, T.; Naito, T.; Takeda, K.; Van Kaer, L.; Okumura, K.; Abo, T. Activation of Hepatic NKT Cells and Subsequent Liver Injury Following Administration of Alpha-Galactosylceramide. Eur. J. Immunol. 2000, 30, 1919–1928. [Google Scholar] [CrossRef]
- Sköld, M.; Behar, S.M. Role of CD1d-Restricted NKT Cells in Microbial Immunity. Infect. Immun. 2003, 71, 5447–5455. [Google Scholar] [CrossRef]
- Ronchi, F.; Falcone, M. Immune Regulation by Invariant NKT Cells in Autoimmunity. Front. Biosci. J. Virtual Libr. 2008, 13, 4827–4837. [Google Scholar] [CrossRef]
- Terabe, M.; Berzofsky, J.A. The Role of NKT Cells in Tumor Immunity. Adv. Cancer Res. 2008, 101, 277–348. [Google Scholar] [CrossRef]
- Umetsu, D.T.; DeKruyff, R.H. A Role for Natural Killer T Cells in Asthma. Nat. Rev. Immunol. 2006, 6, 953–958. [Google Scholar] [CrossRef]
- Lee, Y.J.; Holzapfel, K.L.; Zhu, J.; Jameson, S.C.; Hogquist, K.A. Steady-State Production of IL-4 Modulates Immunity in Mouse Strains and Is Determined by Lineage Diversity of iNKT Cells. Nat. Immunol. 2013, 14, 1146–1154. [Google Scholar] [CrossRef]
- Georgiev, H.; Ravens, I.; Benarafa, C.; Forster, R.; Bernhardt, G. Distinct Gene Expression Patterns Correlate with Developmental and Functional Traits of iNKT Subsets. Nat. Commun. 2016, 7, 13116. [Google Scholar] [CrossRef]
- Bennstein, S.B. Unraveling Natural Killer T-Cells Development. Front. Immunol. 2017, 8, 1950. [Google Scholar] [CrossRef]
- Wang, L.; Wildt, K.F.; Zhu, J.; Zhang, X.; Feigenbaum, L.; Tessarollo, L.; Paul, W.E.; Fowlkes, B.J.; Bosselut, R. Distinct Functions for the Transcription Factors GATA-3 and ThPOK during Intrathymic Differentiation of CD4(+) T Cells. Nat. Immunol. 2008, 9, 1122–1130. [Google Scholar] [CrossRef]
- Amsen, D.; Antov, A.; Jankovic, D.; Sher, A.; Radtke, F.; Souabni, A.; Busslinger, M.; McCright, B.; Gridley, T.; Flavell, R.A. Direct Regulation of Gata3 Expression Determines the T Helper Differentiation Potential of Notch. Immunity 2007, 27, 89–99. [Google Scholar] [CrossRef]
- Kim, P.J.; Pai, S.Y.; Brigl, M.; Besra, G.S.; Gumperz, J.; Ho, I.C. GATA-3 Regulates the Development and Function of Invariant NKT Cells. J. Immunol. 2006, 177, 6650–6659. [Google Scholar] [CrossRef]
- Engel, I.; Seumois, G.; Chavez, L.; Samaniego-Castruita, D.; White, B.; Chawla, A.; Mock, D.; Vijayanand, P.; Kronenberg, M. Innate-like Functions of Natural Killer T Cell Subsets Result from Highly Divergent Gene Programs. Nat. Immunol. 2016, 17, 728–739. [Google Scholar] [CrossRef]
- Iyoda, T.; Shimizu, K.; Endo, T.; Watanabe, T.; Taniuchi, I.; Aoshima, H.; Satoh, M.; Nakazato, H.; Yamasaki, S.; Fujii, S.I. Zeb2 Regulates Differentiation of Long-Lived Effector of Invariant Natural Killer T Cells. Commun. Biol. 2023, 6, 1070. [Google Scholar] [CrossRef]
- Patino-Lopez, G.; Hevezi, P.; Lee, J.; Willhite, D.; Verge, G.M.; Lechner, S.M.; Ortiz-Navarrete, V.; Zlotnik, A. Human Class-I Restricted T Cell Associated Molecule Is Highly Expressed in the Cerebellum and Is a Marker for Activated NKT and CD8+ T Lymphocytes. J. Neuroimmunol. 2006, 171, 145–155. [Google Scholar] [CrossRef]
- Wolf, F.A.; Hamey, F.K.; Plass, M.; Solana, J.; Dahlin, J.S.; Gottgens, B.; Rajewsky, N.; Simon, L.; Theis, F.J. PAGA: Graph Abstraction Reconciles Clustering with Trajectory Inference through a Topology Preserving Map of Single Cells. Genome Biol. 2019, 20, 59. [Google Scholar] [CrossRef]
- Wang, H.; Hogquist, K.A. CCR7 Defines a Precursor for Murine iNKT Cells in Thymus and Periphery. Elife 2018, 7, e34793. [Google Scholar] [CrossRef]
- Baranek, T.; Lebrigand, K.; de Amat Herbozo, C.; Gonzalez, L.; Bogard, G.; Dietrich, C.; Magnone, V.; Boisseau, C.; Jouan, Y.; Trottein, F.; et al. High Dimensional Single-Cell Analysis Reveals iNKT Cell Developmental Trajectories and Effector Fate Decision. Cell Rep. 2020, 32, 108116. [Google Scholar] [CrossRef]
- Lu, Y.; Zhong, M.C.; Qian, J.; Calderon, V.; Cruz Tleugabulova, M.; Mallevaey, T.; Veillette, A. SLAM Receptors Foster iNKT Cell Development by Reducing TCR Signal Strength after Positive Selection. Nat. Immunol. 2019, 20, 447–457. [Google Scholar] [CrossRef]
- Harsha Krovi, S.; Zhang, J.; Michaels-Foster, M.J.; Brunetti, T.; Loh, L.; Scott-Browne, J.; Gapin, L. Thymic iNKT Single Cell Analyses Unmask the Common Developmental Program of Mouse Innate T Cells. Nat. Commun. 2020, 11, 6238. [Google Scholar] [CrossRef]
- Tuttle, K.D.; Krovi, S.H.; Zhang, J.; Bedel, R.; Harmacek, L.; Peterson, L.K.; Dragone, L.L.; Lefferts, A.; Halluszczak, C.; Riemondy, K.; et al. TCR Signal Strength Controls Thymic Differentiation of iNKT Cell Subsets. Nat. Commun. 2018, 9, 2650. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Svensson, M.N.D.; Venken, K.; Chawla, A.; Liang, S.; Engel, I.; Mydel, P.; Day, J.; Elewaut, D.; Bottini, N.; et al. Altered Thymic Differentiation and Modulation of Arthritis by Invariant NKT Cells Expressing Mutant ZAP70. Nat. Commun. 2018, 9, 2627. [Google Scholar] [CrossRef] [PubMed]
- Mandl, J.N.; Monteiro, J.P.; Vrisekoop, N.; Germain, R.N. T cell-positive selection uses self-ligand binding strength to optimize repertoire recognition of foreign antigens. Immunity 2013, 38, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Azzam, H.S.; Grinberg, A.; Lui, K.; Shen, H.; Shores, E.W.; Love, P.E. CD5 expression is developmentally regulated by T cell receptor (TCR) signals and TCR avidity. J. Exp. Med. 1998, 188, 2301–2311. [Google Scholar] [CrossRef] [PubMed]
- Yagi, R.; Zhong, C.; Northrup, D.L.; Yu, F.; Bouladoux, N.; Spencer, S.; Hu, G.; Barron, L.; Sharma, S.; Nakayama, T.; et al. The Transcription Factor GATA3 Is Critical for the Development of All IL-7Rα-Expressing Innate Lymphoid Cells. Immunity 2014, 40, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Cui, K.; Wilhelm, C.; Hu, G.; Mao, K.; Belkaid, Y.; Zhao, K.; Zhu, J. Group 3 Innate Lymphoid Cells Continuously Require the Transcription Factor GATA-3 after Commitment. Nat. Immunol. 2016, 17, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Webster, K.E.; Kim, H.O.; Kyparissoudis, K.; Corpuz, T.M.; Pinget, G.V.; Uldrich, A.P.; Brink, R.; Belz, G.T.; Cho, J.H.; Godfrey, D.I.; et al. IL-17-Producing NKT Cells Depend Exclusively on IL-7 for Homeostasis and Survival. Mucosal Immunol. 2014, 7, 1058–1067. [Google Scholar] [CrossRef] [PubMed]
- Akbari, O.; Stock, P.; Meyer, E.H.; Freeman, G.J.; Sharpe, A.H.; Umetsu, D.T.; DeKruyff, R.H. ICOS/ICOSL Interaction Is Required for CD4+ Invariant NKT Cell Function and Homeostatic Survival. J. Immunol. 2008, 180, 5448–5456. [Google Scholar] [CrossRef] [PubMed]
- Zietara, N.; Lyszkiewicz, M.; Krueger, A.; Weiss, S. ICOS-Dependent Stimulation of NKT Cells by Marginal Zone B Cells. Eur. J. Immunol. 2011, 41, 3125–3134. [Google Scholar] [CrossRef]
- Look, A.; Burns, D.; Tews, I.; Roghanian, A.; Mansour, S. Towards a better understanding of human iNKT cell subpopulations for improved clinical outcomes. Front. Immunol. 2023, 14, 1176724. [Google Scholar] [CrossRef]
- Li, Y.R.; Zhou, Y.; Kim, Y.J.; Zhu, Y.; Ma, F.; Yu, J.; Wang, Y.-C.; Chen, X.; Li, Z.; Zeng, S.; et al. Development of allogeneic HSC-engineered iNKT cells for off-the-shelf cancer immunotherapy. Cell Rep. Med. 2021, 2, 100449. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.R.; Zhou, Y.; Yu, J.; Zhu, Y.; Lee, D.; Zhu, E.; Li, Z.; Kim, Y.J.; Zhou, K.; Fang, Y.; et al. Engineering allorejection-resistant CAR-NKT cells from hematopoietic stem cells for off-the-shelf cancer immunotherapy. Mol. Ther. 2024, 32, 1849–1874. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tai, T.-S.; Yang, H.-Y.; Chuang, W.-C.; Huang, Y.-W.; Ho, I.-C.; Tsai, C.-C.; Chuang, Y.-T. ScRNA-Seq Analyses Define the Role of GATA3 in iNKT Cell Effector Lineage Differentiation. Cells 2024, 13, 1073. https://doi.org/10.3390/cells13121073
Tai T-S, Yang H-Y, Chuang W-C, Huang Y-W, Ho I-C, Tsai C-C, Chuang Y-T. ScRNA-Seq Analyses Define the Role of GATA3 in iNKT Cell Effector Lineage Differentiation. Cells. 2024; 13(12):1073. https://doi.org/10.3390/cells13121073
Chicago/Turabian StyleTai, Tzong-Shyuan, Huang-Yu Yang, Wan-Chu Chuang, Yu-Wen Huang, I-Cheng Ho, Ching-Chung Tsai, and Ya-Ting Chuang. 2024. "ScRNA-Seq Analyses Define the Role of GATA3 in iNKT Cell Effector Lineage Differentiation" Cells 13, no. 12: 1073. https://doi.org/10.3390/cells13121073
APA StyleTai, T.-S., Yang, H.-Y., Chuang, W.-C., Huang, Y.-W., Ho, I.-C., Tsai, C.-C., & Chuang, Y.-T. (2024). ScRNA-Seq Analyses Define the Role of GATA3 in iNKT Cell Effector Lineage Differentiation. Cells, 13(12), 1073. https://doi.org/10.3390/cells13121073