
Excess Dally-like Induces Malformation of Drosophila Legs

 , ,
, ,
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Drosophila Stocks
2.2. Immunochemistry Staining
2.3. Measurements and Data Statistical Analysis
3. Results
3.1. Extra Dlp Expression Induces Leg Deformities
3.2. Excess Dlp Induces Morphology Modification of Leg Disc with No Apparent Cell Proliferation Alteration
3.3. Excess Dlp Induces Apoptotic Cell Death
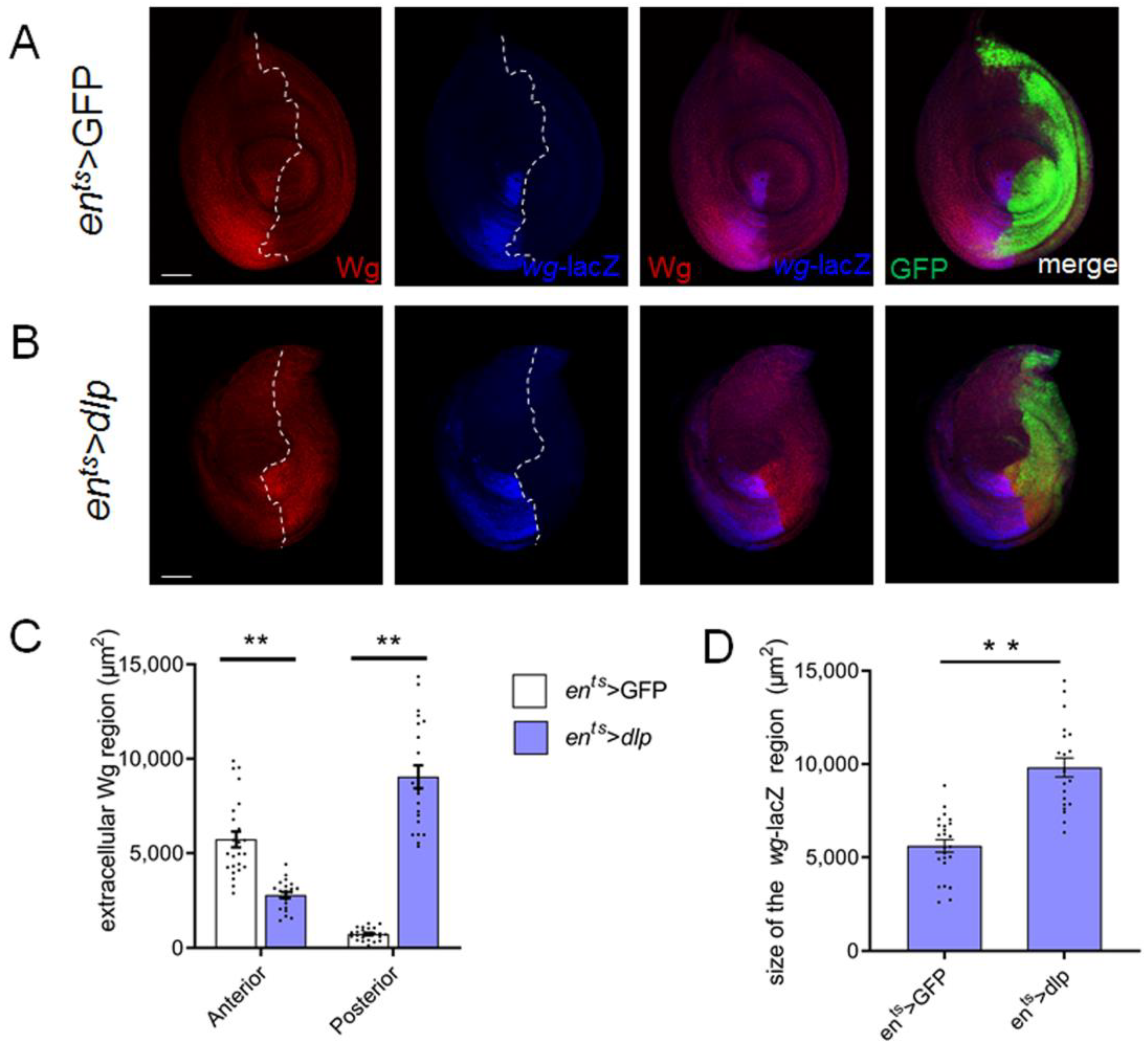
3.4. Extra Dlp Induces Wg Mis-Distribution
3.5. Excess Dlp Causes Mis-Expression of Leg Patterning Gene Dll
4. Discussion
4.1. Cell Death Is the Potential Factor Inducing Deformities of Leg Discs
4.2. Excess Dlp Results in Abnormal Distribution of Wg
4.3. Compared to Dac and Hth, Dll Is More Responsive to the Excess Dlp
4.4. The Limitations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jen, Y.H.L.; Musacchio, M.; Lander, A.D. Glypican-1 controls brain size through regulation of fibroblast growth factor signaling in early neurogenesis. Neural Dev. 2009, 4, 33. [Google Scholar] [CrossRef]
- Iozzo, R.V. Series Introduction: Heparan sulfate proteoglycans: Intricate molecules with intriguing functions. J. Clin. Investig. 2001, 108, 165–167. [Google Scholar] [CrossRef] [PubMed]
- Christianson, H.C.; Belting, M. Heparan sulfate proteoglycan as a cell-surface endocytosis receptor. Matrix Biol. 2014, 35, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Kirn-Safran, C.; Farach-Carson, M.C.; Carson, D.D. Multifunctionality of extracellular and cell surface heparan sulfate proteoglycans. Cell. Mol. Life Sci. 2009, 6, 3421–3434. [Google Scholar] [CrossRef] [PubMed]
- Xiong, A.; Kundu, S.; Forsberg-Nilsson, K. Heparan sulfate in the regulation of neural differentiation and glioma development. FEBS J. 2014, 281, 4993–5008. [Google Scholar] [CrossRef] [PubMed]
- Li, J.P.; Kusche-Gullberg, M. Heparan sulfate: Biosynthesis, Structure, and Function. Int. Rev. Cell Mol. Biol. 2016, 325, 215–273. [Google Scholar] [CrossRef] [PubMed]
- Dreyfuss, J.L.; Regatieri, C.V.; Jarrouge, T.R.; Cavalheiro, R.P.; Sampaio, L.O.; Nader, H.B. Heparan sulfate proteoglycans: Stru-cture, protein interactions and cell signaling. An. Acad. Bras. Cienc. 2009, 81, 409–429. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Lin, X. Shaping morphogen gradients by proteoglycans. Cold Spring Harb. Perspect. Biol. 2009, 1, a002493. [Google Scholar] [CrossRef]
- Xie, M.; Li, J. Heparan sulfate proteoglycan—A common receptor for diverse cytokines. Cell. Signal. 2019, 54, 115–121. [Google Scholar] [CrossRef]
- Filmus, J.; Selleck, S.B. Glypicans: Proteoglycans with a surprise. J. Clin. Investig. 2001, 108, 497–501. [Google Scholar] [CrossRef]
- Waghmare, I.; Wang, X.; Page-McCaw, A. Dally-like protein sequesters multiple Wnt ligands in the Drosophila germarium. Dev. Biol. 2020, 464, 88–102. [Google Scholar] [CrossRef] [PubMed]
- Nakato, E.; Kamimura, K.; Knudsen, C.; Masutani, S.; Takemura, M.; Hayashi, Y.; Akiyama, T.; Nakato, H. Differential heparan sulfate dependency of the Drosophila glypicans. J. Biol. Chem. 2024, 300, 105544. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Gao, W.; Zhang, Y.F.; Ho, M. Glypicans as Cancer Therapeutic Targets. Trends Cancer 2018, 4, 741–754. [Google Scholar] [CrossRef]
- Han, C.; Yan, D.; Belenkaya, T.Y.; Lin, X. Drosophila glypicans Dally and Dally-like shape the extracellular Wingless morphogen gradient in the wing disc. Development 2005, 132, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Wu, Y.; Yang, Y.; Belenkaya, T.Y.; Tang, X.; Lin, X. The cell-surface proteins Dally-like and Ihog differentia-lly regulate Hedgehog signaling strength and range during development. Development 2010, 137, 2033–2044. [Google Scholar] [CrossRef]
- Williams, E.H.; Pappano, W.N.; Saunders, A.M.; Kim, M.S.; Leahy, D.J.; Beachy, P.A. Dally-like core protein and its mammalian homologues mediate stimulatory and inhibitory effects on Hedgehog signal response. Proc. Natl. Acad. Sci. USA 2010, 107, 5869–5874. [Google Scholar] [CrossRef]
- Kreuger, J.; Perez, L.; Giraldez, A.J.; Cohen, S.M. Opposing activities of Dally-like glypican at high and low levels of Wingless morphogen activity. Dev. Cell 2004, 7, 503–512. [Google Scholar] [CrossRef]
- Gallet, A.; Staccini-Lavenant, L.; Thérond, P.P. Cellular trafficking of the glypican Dally-like is required for full-strength Hedgehog signaling and wingless transcytosis. Dev. Cell 2008, 14, 712–725. [Google Scholar] [CrossRef]
- Tu, R.; Duan, B.; Song, X.; Xie, T. Dlp-mediated Hh and Wnt signaling interdependence is critical in the niche for germline stem cell progeny differentiation. Sci. Adv. 2020, 6, eaaz0480. [Google Scholar] [CrossRef]
- Cohen, B.; Wimmer, E.A.; Cohen, S.M. Early development of leg and wing primordia in the Drosophila embryo. Mech. Dev. 1991, 33, 229–240. [Google Scholar] [CrossRef]
- Kojima, T. The mechanism of Drosophila leg development along the proximodistal axis. Dev. Growth Differ. 2004, 46, 115–129. [Google Scholar] [CrossRef]
- Couso, J.P.; Bishop, S.A. Proximo-distal development in the legs of Drosophila. Int. J. Dev. Biol. 1998, 42, 345–352. [Google Scholar] [PubMed]
- Jiang, J.; Struhl, G. Complementary and mutually exclusive activities of decapentaplegic and wingless organize axial patterning during Drosophila leg development. Cell 1996, 86, 401–409. [Google Scholar] [CrossRef]
- Basler, K.; Struhl, G. Compartment boundaries and the control of Drosophila limb pattern by hedgehog protein. Nature 1994, 368, 208–214. [Google Scholar] [CrossRef]
- Johnston, L.A.; Schubiger, G. Ectopic expression of wingless in imaginal discs interferes with decapentaplegic expression and alters cell determination. Development 1996, 122, 3519–3529. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Cohen, S.M. Proximodistal axis formation in the Drosophila leg: Subdivision into proximal and distal domains by Homothorax and Distal-less. Development 1999, 126, 109–117. [Google Scholar] [CrossRef]
- Brook, W.J.; Cohen, S.M. Antagonistic interactions between wingless and decapentaplegic responsible for dorsal-ventral pattern in the Drosophila Leg. Science 1996, 273, 1373–1377. [Google Scholar] [CrossRef]
- Lecuit, T.; Cohen, S.M. Proximal–distal axis formation in the Drosophila leg. Nature 1997, 388, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Niwa, N.; Hiromi, Y.; Okabe, M. A conserved developmental program for sensory organ formation in Drosophila melanogaster. Nature genetics 2004, 36, 293–297. [Google Scholar] [CrossRef]
- Gonsalves, F.C.; DasGupta, R. Function of the wingless signaling pathway in Drosophila. Methods Mol. Biol. 2008, 469, 115–125. [Google Scholar] [CrossRef]
- Estella, C.; Mann, R.S. Logic of Wg and Dpp induction of distal and medial fates in the Drosophila leg. Development 2008, 135, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Gorfinkiel, N.; Morata, G.; Guerrero, I. The homeobox gene Distal-less induces ventral appendage development in Drosophila. Genes Dev. 1997, 11, 2259–2271. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Garijo, A.; Fuchs, Y.; Steller, H. Apoptotic cells can induce non-autonomous apoptosis through the TNF pathway. eLife 2013, 2, e01004. [Google Scholar] [CrossRef] [PubMed]
- Giraldez, A.J.; Cohen, S.M. Wingless and Notch signaling provide cell survival cues and control cell proliferation during wing development. Development 2003, 130, 6533–6543. [Google Scholar] [CrossRef] [PubMed]
- Baeg, G.H.; Lin, X.; Khare, N.; Baumgartner, S.; Perrimon, N. Heparan sulfate proteoglycans are critical for the organization of the extracellular distribution of Wingless. Development 2001, 128, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Wu, Y.; Feng, Y.; Lin, S.C.; Lin, X. The core protein of glypican Dally-like determines its biphasic activity in wingless morphogen signaling. Dev. Cell 2009, 17, 470–481. [Google Scholar] [CrossRef] [PubMed]
- Campbell, G.; Tomlinson, A. The roles of the homeobox genes aristaless and Distal-less in patterning the legs and wings of Drosophila. Development 1998, 125, 4483–4493. [Google Scholar] [CrossRef] [PubMed]
- Abu-Shaar, M.; Mann, R.S. Generation of multiple antagonistic domains along the proximodistal axis during Drosophila leg de-velopment. Development 1998, 125, 3821–3830. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Benjumea, F.J.; Cohen, B.; Cohen, S.M. Cell interaction between compartments establishes the proximal-distal axis of Drosophila legs. Nature 1994, 372, 175–179. [Google Scholar] [CrossRef]
- Zuniga, A.; Zeller, R.; Probst, S. The molecular basis of human congenital limb malformations. Wiley Interdiscip. Rev. Dev. Biol. 2012, 1, 803–822. [Google Scholar] [CrossRef]
- Talamllo, A.; Bastida, M.F.; Fernandez-Teran, M.; Ros, M.A. The developing limb and the control of the number of digits. Clin. Genet. 2005, 67, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Filmus, J. The contribution of in vivo manipulation of gene expression to the understanding of the function of glypicans. Glycoconj. J. 2002, 19, 319–323. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Wang, Y.; Zhao, W.; Yang, S.; Moussian, B.; Zhao, Z.; Zhang, J.; Dong, W. Excess Dally-like Induces Malformation of Drosophila Legs. Cells 2024, 13, 1199. https://doi.org/10.3390/cells13141199
Zhang X, Wang Y, Zhao W, Yang S, Moussian B, Zhao Z, Zhang J, Dong W. Excess Dally-like Induces Malformation of Drosophila Legs. Cells. 2024; 13(14):1199. https://doi.org/10.3390/cells13141199
Chicago/Turabian StyleZhang, Xubo, Yi Wang, Wenting Zhao, Shumin Yang, Bernard Moussian, Zhangwu Zhao, Jianzhen Zhang, and Wei Dong. 2024. "Excess Dally-like Induces Malformation of Drosophila Legs" Cells 13, no. 14: 1199. https://doi.org/10.3390/cells13141199
APA StyleZhang, X., Wang, Y., Zhao, W., Yang, S., Moussian, B., Zhao, Z., Zhang, J., & Dong, W. (2024). Excess Dally-like Induces Malformation of Drosophila Legs. Cells, 13(14), 1199. https://doi.org/10.3390/cells13141199