
Fat Phagocytosis Promotes Anti-Inflammatory Responses of Macrophages in a Mouse Model of Osteonecrosis


Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Human ONFH Samples
2.2. Animals
2.3. Histomorphometry
2.4. Cell Culture
2.5. Western Blot
2.6. RNA Isolation and RT-qPCR
2.7. Bulk RNA Sequencing
2.8. Statistical Analysis
3. Results
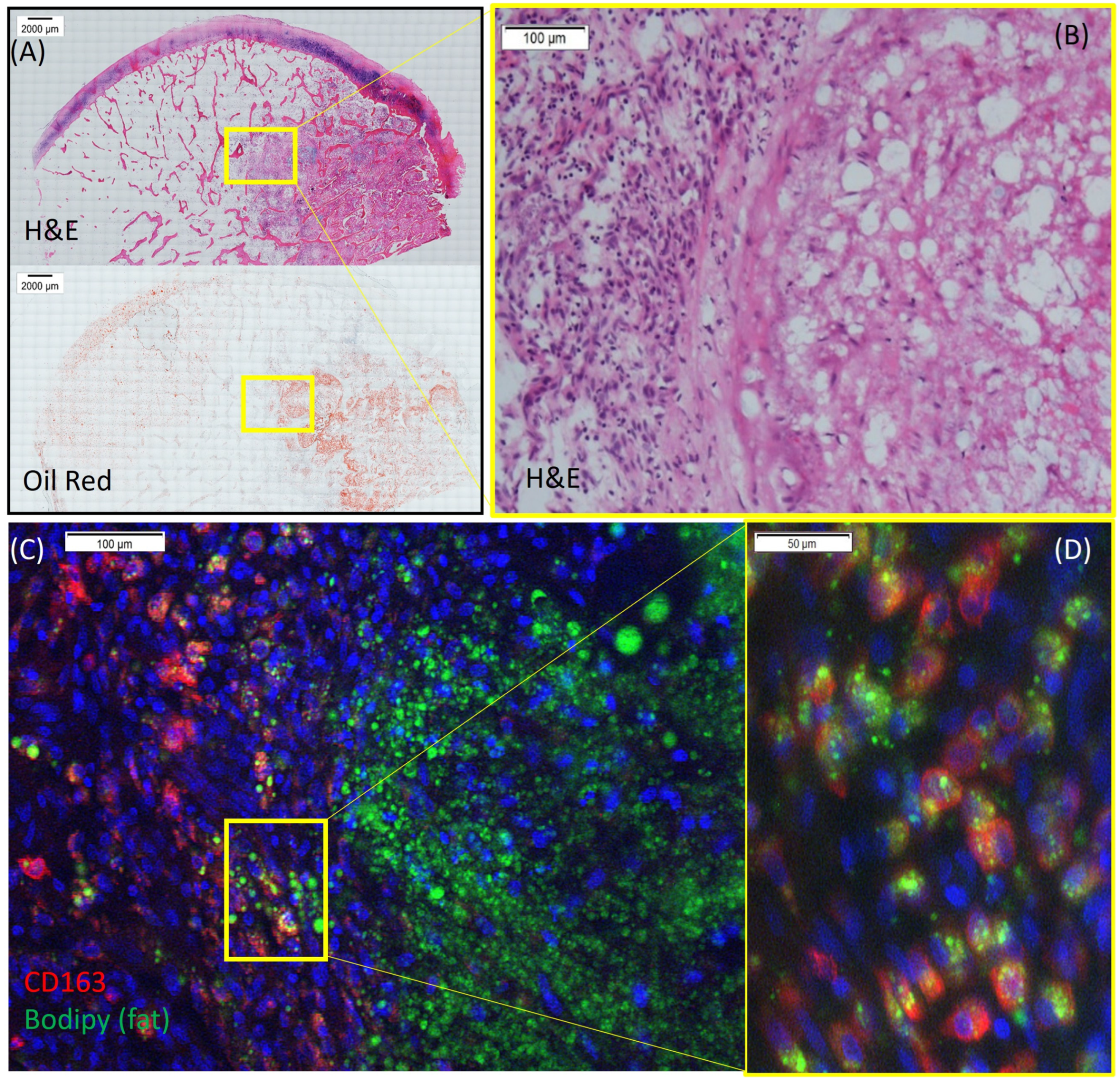
3.1. Phagocytosis of Fat by Macrophages in Human ONFH Sample
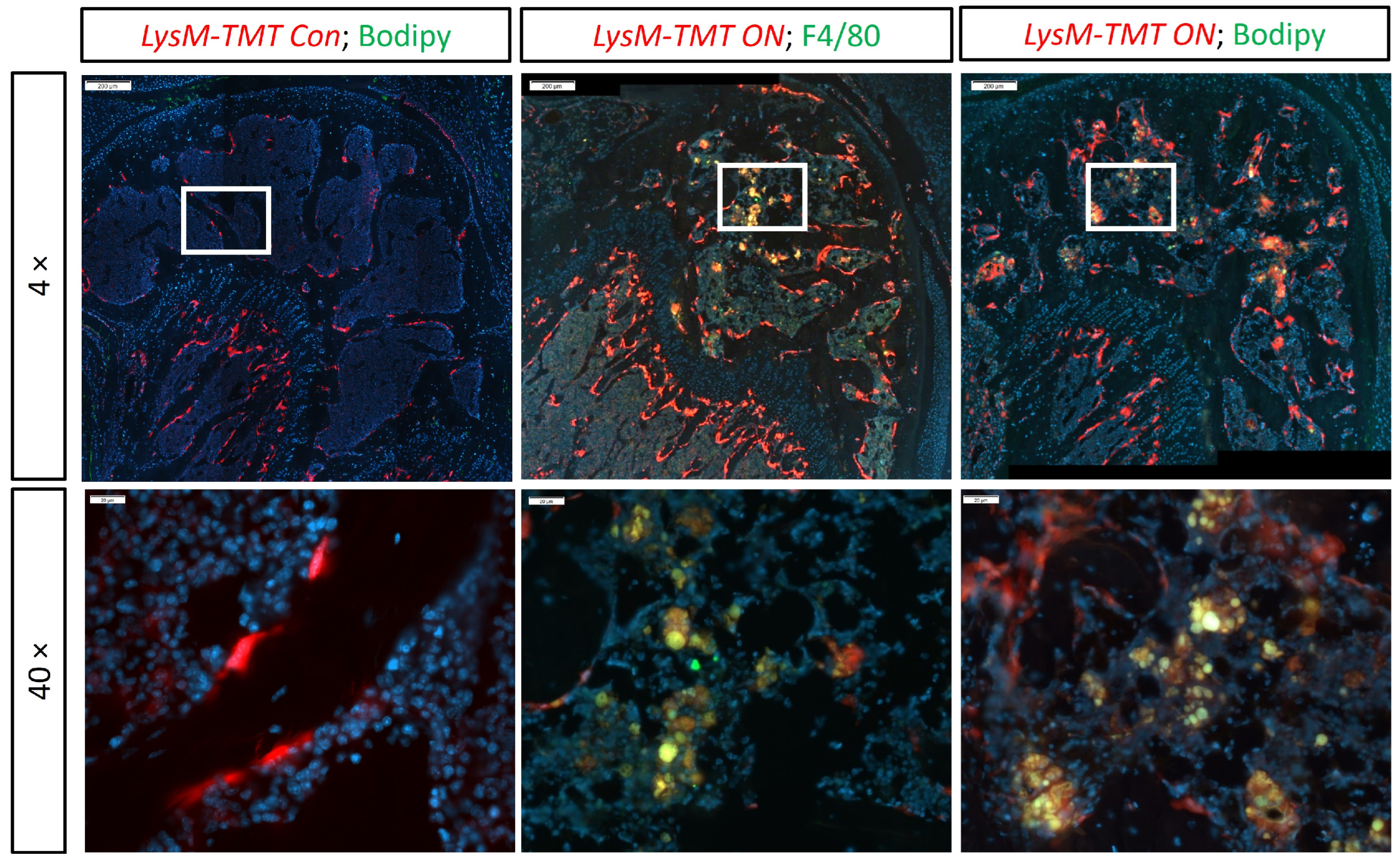
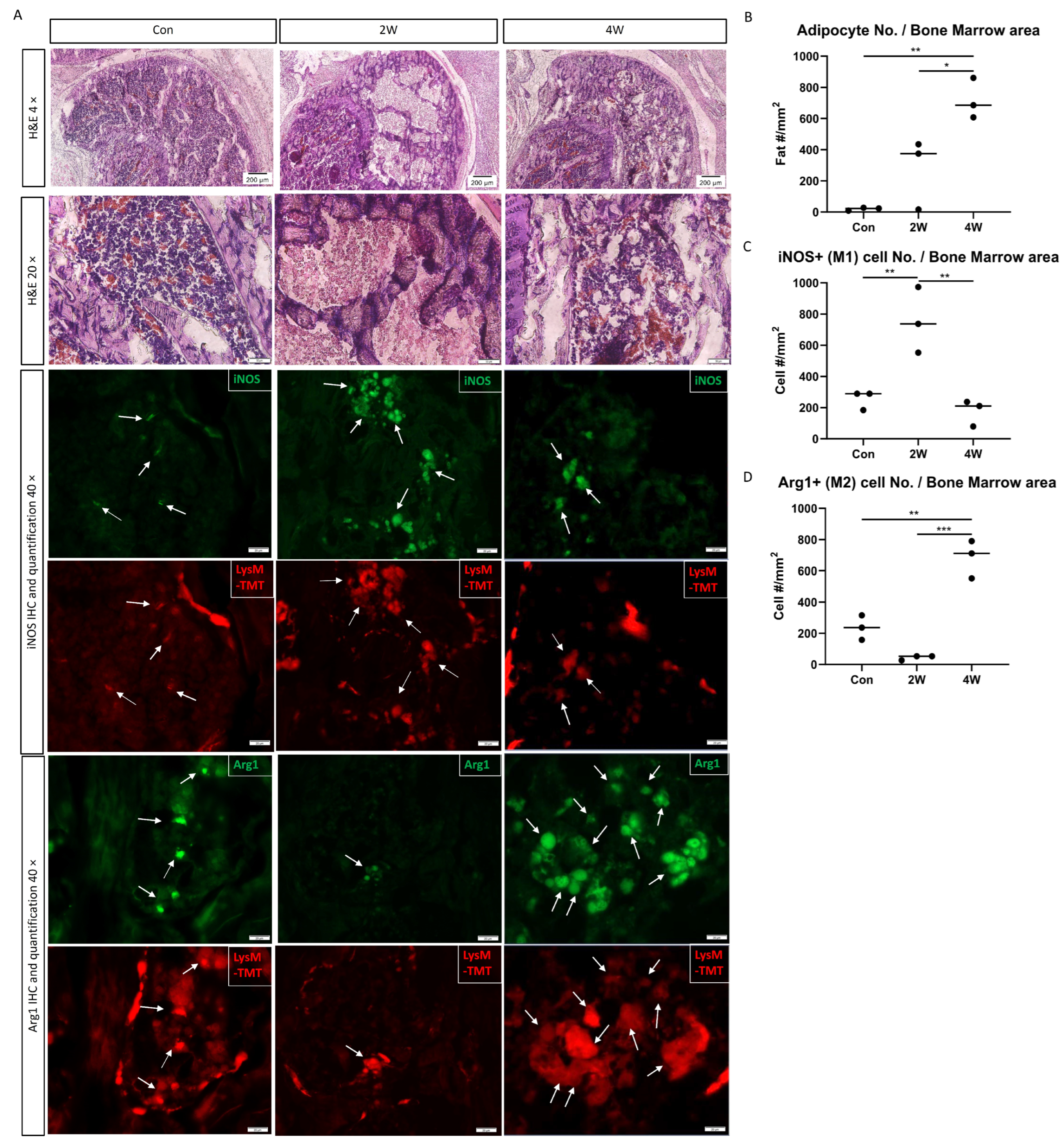
3.2. Phagocytosis of Fat by Macrophages in ON Mouse Model
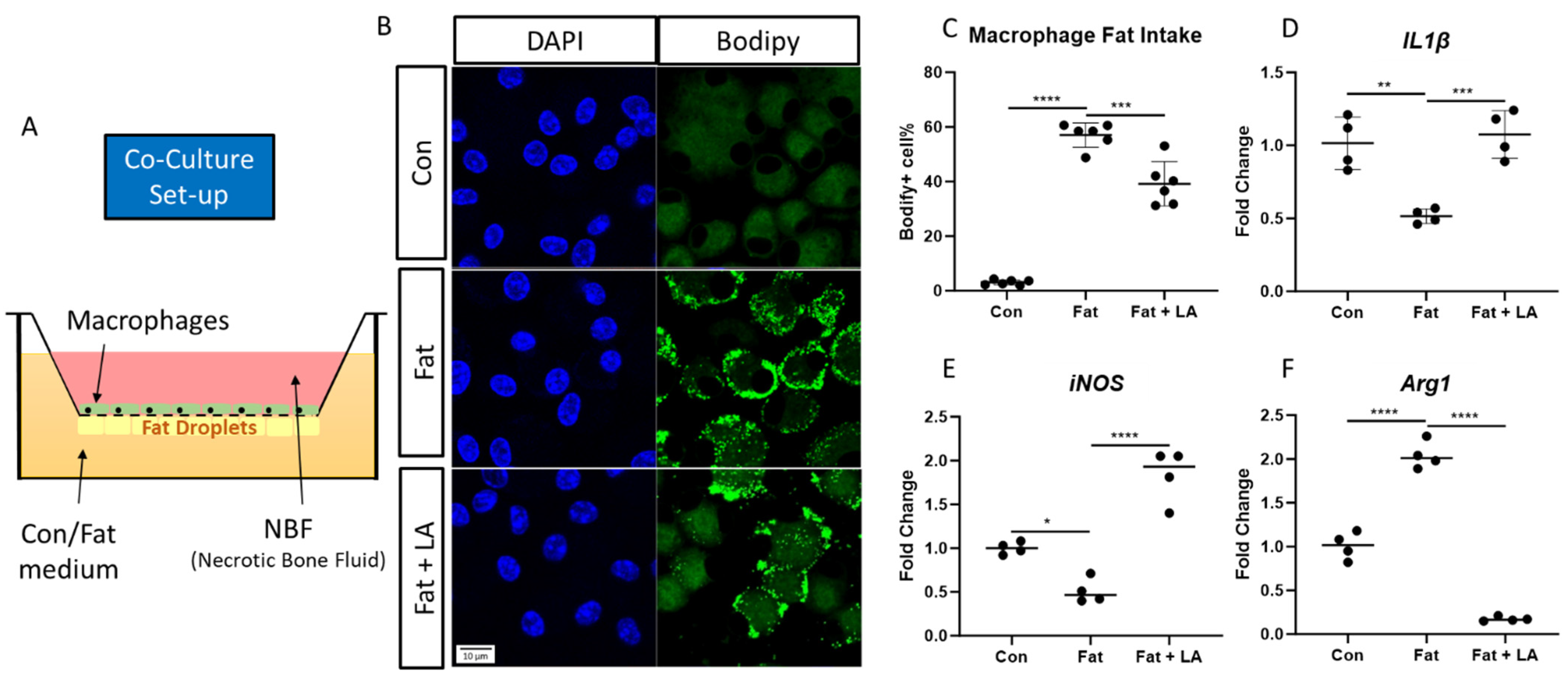
3.3. Macrophages and Necrotic Fat Co-Culture
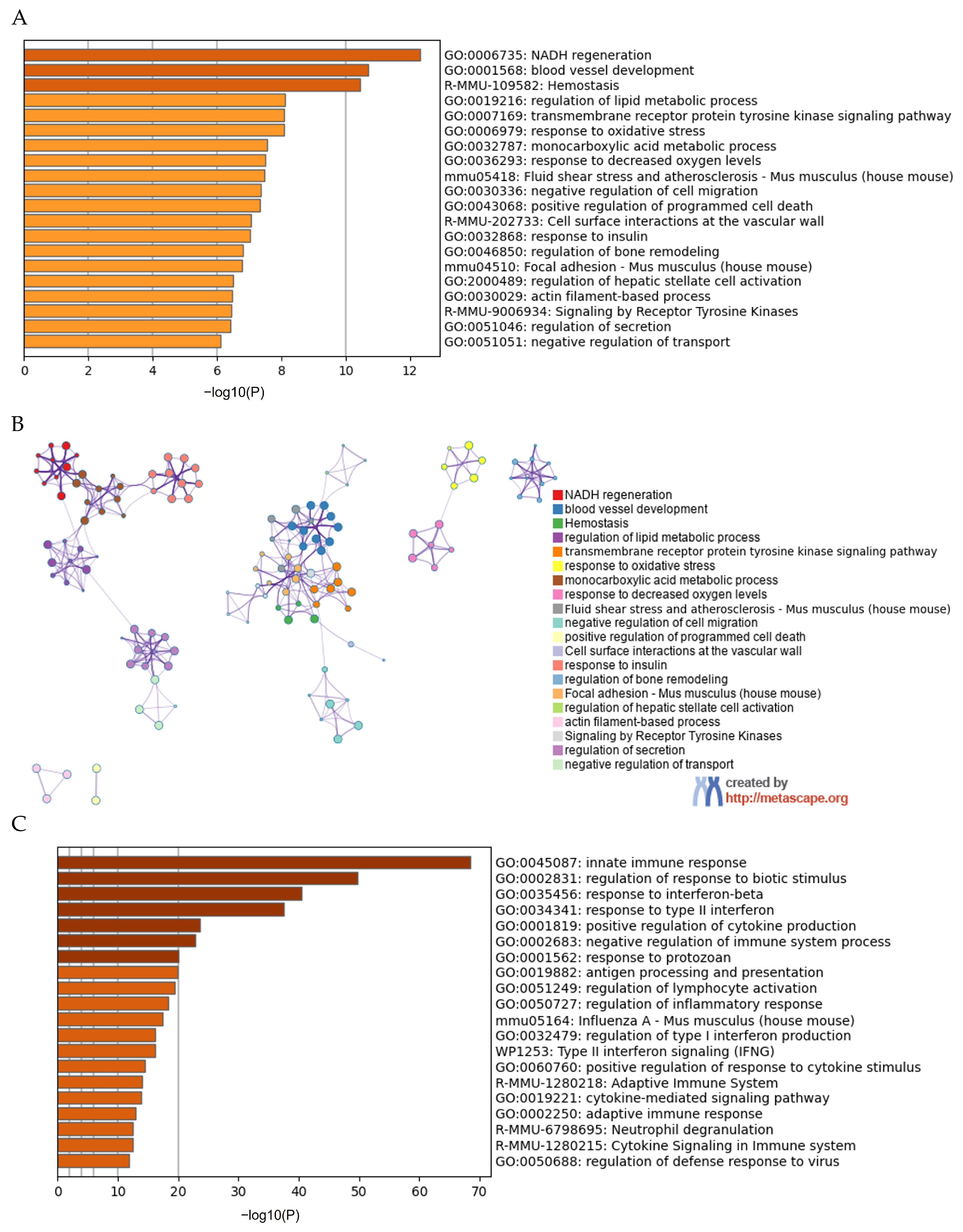
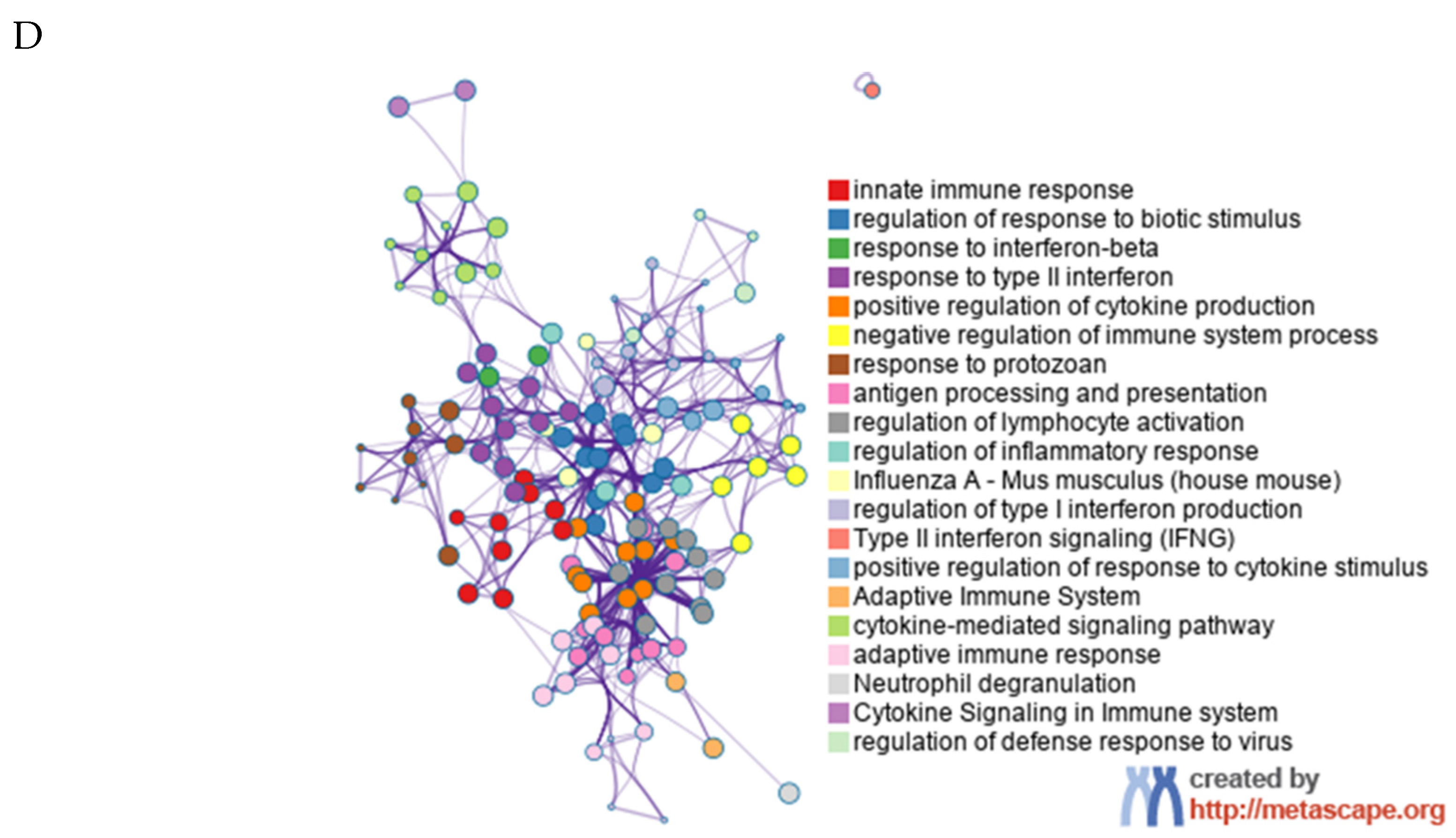
3.4. Effects of Fat Phagocytosis on Transcriptional Profile of Macrophages
3.5. Changes in Macrophage Markers after ON In Vivo
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Goodman, S.B.; Maruyama, M. Inflammation, Bone Healing and Osteonecrosis: From Bedside to Bench. J. Inflamm. Res. 2020, 13, 913–923. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Ren, Y.; Park, M.S.; Kim, H.K.W. Damage associated molecular patterns in necrotic femoral head inhibit osteogenesis and promote fibrogenesis of mesenchymal stem cells. Bone 2022, 154, 116215. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Yao, Z.; Xue, L.; Wang, D.; Tan, Z. The role of immune cells in modulating chronic inflammation and osteonecrosis. Front. Immunol. 2022, 13, 1064245. [Google Scholar] [CrossRef] [PubMed]
- Lavin, Y.; Merad, M. Macrophages: Gatekeepers of tissue integrity. Cancer Immunol. Res. 2013, 1, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T.A.; Chawla, A.; Pollard, J.W. Macrophage biology in development, homeostasis and disease. Nature 2013, 496, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.J. Macrophage Polarization. Annu. Rev. Physiol. 2017, 79, 541–566. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Xu, X.H.; Jin, L. Macrophage Polarization in Physiological and Pathological Pregnancy. Front. Immunol. 2019, 10, 792. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, R.; Kamiya, N.; Kuroyanagi, G.; Ren, Y.; Kim, H.K.W. Development of a murine model of ischemic osteonecrosis to study the effects of aging on bone repair. J. Orthop. Res. 2021, 39, 2663–2670. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.G.; Rao, V.M.; Dalinka, M.; Spritzer, C.E.; Axel, L.; Gefter, W.; Kricun, M.; Steinberg, M.E.; Kressel, H.Y. Hematopoietic and fatty bone marrow distribution in the normal and ischemic hip: New observations with 1.5-T MR imaging. Radiology 1986, 161, 199–202. [Google Scholar] [CrossRef]
- Deng, Z.; Aguirre-Flores, M.; Kim, H.K.W.; Ren, Y. Obesity impairs revascularization and bone healing in a mouse model of osteonecrosis. J. Orthop. Res. 2023, 42, 811–820. [Google Scholar] [CrossRef]
- Singh, A.; Mayengbam, S.S.; Yaduvanshi, H.; Wani, M.R.; Bhat, M.K. Obesity Programs Macrophages to Support Cancer Progression. Cancer Res. 2022, 82, 4303–4312. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.W.; Park, M.S.; Alves do Monte, F.; Gokani, V.; Aruwajoye, O.O.; Ren, Y. Minimally Invasive Necrotic Bone Washing Improves Bone Healing After Femoral Head Ischemic Osteonecrosis: An Experimental Investigation in Immature Pigs. J. Bone Jt. Surg. 2021, 103, 1193–1202. [Google Scholar] [CrossRef] [PubMed]
- Qiu, B.; Simon, M.C. BODIPY 493/503 Staining of Neutral Lipid Droplets for Microscopy and Quantification by Flow Cytometry. Bio-Protoc. 2016, 6, e1912. [Google Scholar] [CrossRef] [PubMed]
- Bowles, A.C.; Scruggs, B.A.; Bunnell, B.A. Mesenchymal stem cell-based therapy in a mouse model of experimental autoimmune encephalomyelitis (EAE). Methods Mol. Biol. 2014, 1213, 303–319. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, I.; Zweifel, M.E.; Courtemanche, N.; Pollard, T.D. Latrunculin A Accelerates Actin Filament Depolymerization in Addition to Sequestering Actin Monomers. Curr. Biol. 2018, 28, 3183–3192.e2. [Google Scholar] [CrossRef]
- Patel, N.R.; Bole, M.; Chen, C.; Hardin, C.C.; Kho, A.T.; Mih, J.; Deng, L.; Butler, J.; Tschumperlin, D.; Fredberg, J.J.; et al. Cell elasticity determines macrophage function. PLoS ONE 2012, 7, e41024. [Google Scholar] [CrossRef] [PubMed]
- Ruffell, D.; Mourkioti, F.; Gambardella, A.; Kirstetter, P.; Lopez, R.G.; Rosenthal, N.; Nerlov, C. A CREB-C/EBPbeta cascade induces M2 macrophage-specific gene expression and promotes muscle injury repair. Proc. Natl. Acad. Sci. USA 2009, 106, 17475–17480. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W., Jr. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investg. 2003, 112, 1796–1808. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Sheng, H.H.; Zhang, G.G.; Cheung, W.H.; Chan, C.W.; Wang, Y.X.; Lee, K.M.; Wang, H.F.; Leung, K.S.; Qin, L.L. Elevated adipogenesis of marrow mesenchymal stem cells during early steroid-associated osteonecrosis development. J. Orthop. Surg. Res. 2007, 2, 15. [Google Scholar] [CrossRef] [PubMed]
- Motomura, G.; Yamamoto, T.; Miyanishi, K.; Yamashita, A.; Sueishi, K.; Iwamoto, Y. Bone marrow fat-cell enlargement in early steroid-induced osteonecrosis--a histomorphometric study of autopsy cases. Pathol. Res. Pract. 2005, 200, 807–811. [Google Scholar] [CrossRef] [PubMed]
- Shin, E.; Koo, J.S. The Role of Adipokines and Bone Marrow Adipocytes in Breast Cancer Bone Metastasis. Int. J. Mol. Sci. 2020, 21, 4967. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, M. Marrow Fat-Secreted Factors as Biomarkers for Osteoporosis. Curr. Osteoporos. Rep. 2019, 17, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Rath, M.; Muller, I.; Kropf, P.; Closs, E.I.; Munder, M. Metabolism via Arginase or Nitric Oxide Synthase: Two Competing Arginine Pathways in Macrophages. Front. Immunol. 2014, 5, 532. [Google Scholar] [CrossRef] [PubMed]
- Su, P.; Wang, Q.; Bi, E.; Ma, X.; Liu, L.; Yang, M.; Qian, J.; Yi, Q. Enhanced Lipid Accumulation and Metabolism Are Required for the Differentiation and Activation of Tumor-Associated Macrophages. Cancer Res. 2020, 80, 1438–1450. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, M.E.; Sun, M.; Zhang, R.; Febbraio, M.; Silverstein, R.; Hazen, S.L. Oxidized phosphatidylserine-CD36 interactions play an essential role in macrophage-dependent phagocytosis of apoptotic cells. J. Exp. Med. 2006, 203, 2613–2625. [Google Scholar] [CrossRef] [PubMed]
- Isa, S.A.; Ruffino, J.S.; Ahluwalia, M.; Thomas, A.W.; Morris, K.; Webb, R. M2 macrophages exhibit higher sensitivity to oxLDL-induced lipotoxicity than other monocyte/macrophage subtypes. Lipids Health Dis. 2011, 10, 229. [Google Scholar] [CrossRef] [PubMed]
- Viola, A.; Munari, F.; Sanchez-Rodriguez, R.; Scolaro, T.; Castegna, A. The Metabolic Signature of Macrophage Responses. Front. Immunol. 2019, 10, 1462. [Google Scholar] [CrossRef]
- Olona, A.; Mukhopadhyay, S.; Hateley, C.; Martinez, F.O.; Gordon, S.; Behmoaras, J. Adipoclast: A multinucleated fat-eating macrophage. BMC Biol. 2021, 19, 246. [Google Scholar] [CrossRef]
- Jaggi, U.; Yang, M.; Matundan, H.H.; Hirose, S.; Shah, P.K.; Sharifi, B.G.; Ghiasi, H. Increased phagocytosis in the presence of enhanced M2-like macrophage responses correlates with increased primary and latent HSV-1 infection. PLoS Pathog. 2020, 16, e1008971. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Shang, Z.; Yang, X.; Zhang, Y.; Cao, L. Macrophage Polarization and the Regulation of Bone Immunity in Bone Homeostasis. J. Inflamm. Res. 2023, 16, 3563–3580. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Li, J.; Xie, X.; Gu, F.; Sui, Z.; Zhang, K.; Yu, T. Macrophage-Osteoclast Associations: Origin, Polarization, and Subgroups. Front. Immunol. 2021, 12, 778078. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Ni, S.; Liu, C.; He, M.; Pan, Y.; Cui, P.; Wang, C.; Ni, X. Neglected immunoregulation: M2 polarization of macrophages triggered by low-dose irradiation plays an important role in bone regeneration. J. Cell. Mol. Med. 2023, 27, 1095–1109. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, Z.; Kim, H.K.W.; Hernandez, P.A.; Ren, Y. Fat Phagocytosis Promotes Anti-Inflammatory Responses of Macrophages in a Mouse Model of Osteonecrosis. Cells 2024, 13, 1227. https://doi.org/10.3390/cells13141227
Deng Z, Kim HKW, Hernandez PA, Ren Y. Fat Phagocytosis Promotes Anti-Inflammatory Responses of Macrophages in a Mouse Model of Osteonecrosis. Cells. 2024; 13(14):1227. https://doi.org/10.3390/cells13141227
Chicago/Turabian StyleDeng, Zhuo, Harry K. W. Kim, Paula A. Hernandez, and Yinshi Ren. 2024. "Fat Phagocytosis Promotes Anti-Inflammatory Responses of Macrophages in a Mouse Model of Osteonecrosis" Cells 13, no. 14: 1227. https://doi.org/10.3390/cells13141227
APA StyleDeng, Z., Kim, H. K. W., Hernandez, P. A., & Ren, Y. (2024). Fat Phagocytosis Promotes Anti-Inflammatory Responses of Macrophages in a Mouse Model of Osteonecrosis. Cells, 13(14), 1227. https://doi.org/10.3390/cells13141227