
circRNAs as Epigenetic Regulators of Integrity in Blood–Brain Barrier Architecture: Mechanisms and Therapeutic Strategies in Multiple Sclerosis
 , , , , ,
, , , , ,  , , and
, , and 
Abstract
:1. Introduction
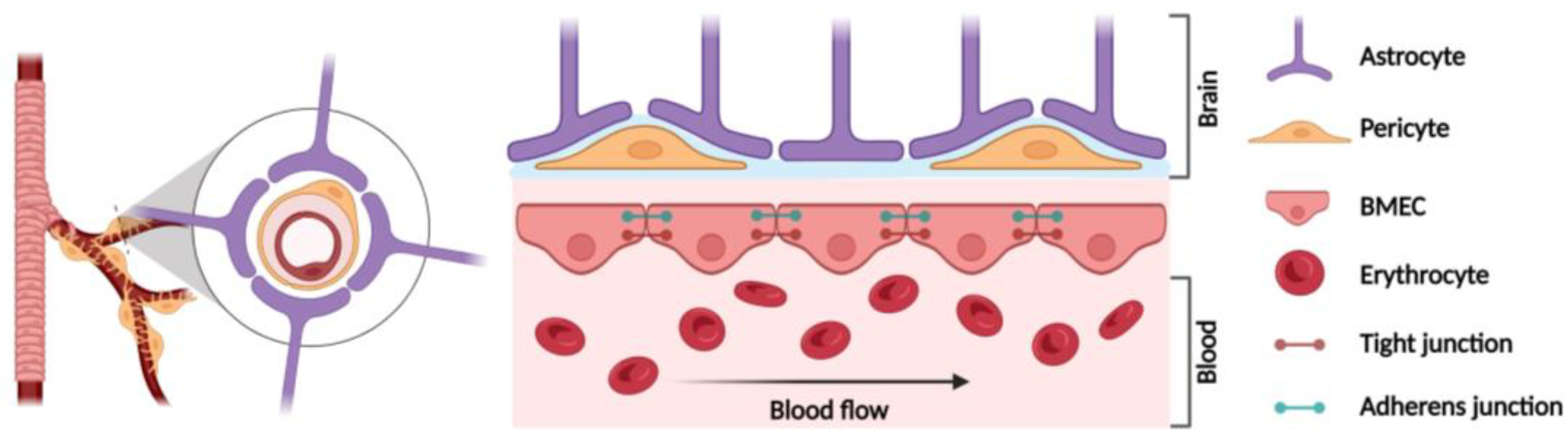
2. Blood–Brain Barrier (BBB) Structure
BMEC Cytoskeleton
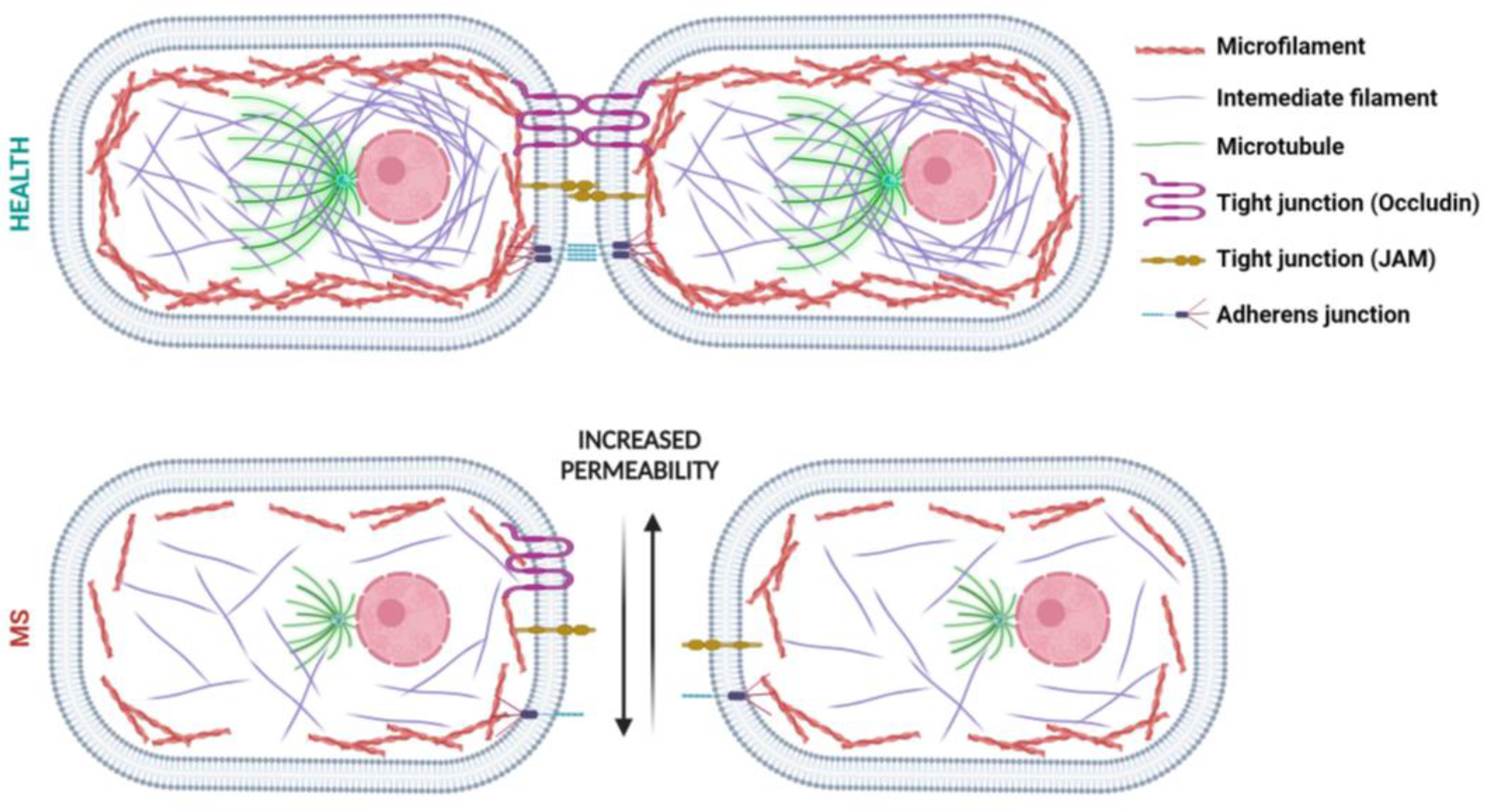
3. BBB Dysfunction in MS
Role of the Cytoskeleton on BBB Hyperpermeability in MS
4. Circular RNAs (circRNAs)
4.1. CircRNAs: Structure and Functions
4.2. CircRNAs as Potential Therapeutic Targets for Cytoskeleton Architecture in BBB Integrity
5. Conclusions, Remarks and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Compston, A.; Coles, A. Multiple Sclerosis. Lancet Lond. Engl. 2008, 372, 1502–1517. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, N.; Razavi, S.; Nikzad, E. Multiple Sclerosis: Pathogenesis, Symptoms, Diagnoses and Cell-Based Therapy. Cell J. 2017, 19, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Soelberg Sorensen, P.; Giovannoni, G.; Montalban, X.; Thalheim, C.; Zaratin, P.; Comi, G. The Multiple Sclerosis Care Unit. Mult. Scler. Houndmills Basingstoke Engl. 2019, 25, 627–636. [Google Scholar] [CrossRef]
- Oh, J.; Vidal-Jordana, A.; Montalban, X. Multiple Sclerosis: Clinical Aspects. Curr. Opin. Neurol. 2018, 31, 752–759. [Google Scholar] [CrossRef]
- Benedict, R.H.B.; Amato, M.P.; DeLuca, J.; Geurts, J.J.G. Cognitive Impairment in Multiple Sclerosis: Clinical Management, MRI, and Therapeutic Avenues. Lancet Neurol. 2020, 19, 860–871. [Google Scholar] [CrossRef] [PubMed]
- McGinley, M.P.; Goldschmidt, C.H.; Rae-Grant, A.D. Diagnosis and Treatment of Multiple Sclerosis: A Review. JAMA 2021, 325, 765–779. [Google Scholar] [CrossRef]
- Carotenuto, A.; Cacciaguerra, L.; Pagani, E.; Preziosa, P.; Filippi, M.; Rocca, M.A. Glymphatic System Impairment in Multiple Sclerosis: Relation with Brain Damage and Disability. Brain J. Neurol. 2022, 145, 2785–2795. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Zeng, M.Y. Sex-Biased Gut Dopamine Signaling in Multiple Sclerosis. Immunity 2023, 56, 2674–2676. [Google Scholar] [CrossRef]
- Lublin, F.D.; Häring, D.A.; Ganjgahi, H.; Ocampo, A.; Hatami, F.; Čuklina, J.; Aarden, P.; Dahlke, F.; Arnold, D.L.; Wiendl, H.; et al. How Patients with Multiple Sclerosis Acquire Disability. Brain J. Neurol. 2022, 145, 3147–3161. [Google Scholar] [CrossRef]
- Wengler, K.; Ha, J.; Syritsyna, O.; Bangiyev, L.; Coyle, P.K.; Duong, T.Q.; Schweitzer, M.E.; He, X. Abnormal Blood-Brain Barrier Water Exchange in Chronic Multiple Sclerosis Lesions: A Preliminary Study. Magn. Reson. Imaging 2020, 70, 126–133. [Google Scholar] [CrossRef]
- Zierfuss, B.; Larochelle, C.; Prat, A. Blood-Brain Barrier Dysfunction in Multiple Sclerosis: Causes, Consequences, and Potential Effects of Therapies. Lancet Neurol. 2024, 23, 95–109. [Google Scholar] [CrossRef]
- Bayir, E.; Sendemir, A. Role of Intermediate Filaments in Blood-Brain Barrier in Health and Disease. Cells 2021, 10, 1400. [Google Scholar] [CrossRef]
- Hohmann, T.; Dehghani, F. The Cytoskeleton-A Complex Interacting Meshwork. Cells 2019, 8, 362. [Google Scholar] [CrossRef]
- Stamatovic, S.M.; Johnson, A.M.; Keep, R.F.; Andjelkovic, A.V. Junctional Proteins of the Blood-Brain Barrier: New Insights into Function and Dysfunction. Tissue Barriers 2016, 4, e1154641. [Google Scholar] [CrossRef]
- Mentor, S.; Makhathini, K.B.; Fisher, D. The Role of Cytoskeletal Proteins in the Formation of a Functional In Vitro Blood-Brain Barrier Model. Int. J. Mol. Sci. 2022, 23, 742. [Google Scholar] [CrossRef] [PubMed]
- Haase, S.; Linker, R.A. Inflammation in Multiple Sclerosis. Ther. Adv. Neurol. Disord. 2021, 14, 17562864211007687. [Google Scholar] [CrossRef]
- Wang, K.; Song, F.; Fernandez-Escobar, A.; Luo, G.; Wang, J.-H.; Sun, Y. The Properties of Cytokines in Multiple Sclerosis: Pros and Cons. Am. J. Med. Sci. 2018, 356, 552–560. [Google Scholar] [CrossRef]
- Sweeney, M.D.; Sagare, A.P.; Zlokovic, B.V. Blood-Brain Barrier Breakdown in Alzheimer Disease and Other Neurodegenerative Disorders. Nat. Rev. Neurol. 2018, 14, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Löscher, W.; Friedman, A. Structural, Molecular, and Functional Alterations of the Blood-Brain Barrier during Epileptogenesis and Epilepsy: A Cause, Consequence, or Both? Int. J. Mol. Sci. 2020, 21, 591. [Google Scholar] [CrossRef]
- Steinruecke, M.; Lonergan, R.M.; Selvaraj, B.T.; Chandran, S.; Diaz-Castro, B.; Stavrou, M. Blood-CNS Barrier Dysfunction in Amyotrophic Lateral Sclerosis: Proposed Mechanisms and Clinical Implications. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2023, 43, 642–654. [Google Scholar] [CrossRef]
- Liu, K.-S.; Pan, F.; Mao, X.-D.; Liu, C.; Chen, Y.-J. Biological Functions of Circular RNAs and Their Roles in Occurrence of Reproduction and Gynecological Diseases. Am. J. Transl. Res. 2019, 11, 1–15. [Google Scholar] [PubMed]
- Cashion, J.M.; Young, K.M.; Sutherland, B.A. How Does Neurovascular Unit Dysfunction Contribute to Multiple Sclerosis? Neurobiol. Dis. 2023, 178, 106028. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Cai, Y.; Xu, J. Circular RNAs: Biogenesis, Mechanism, and Function in Human Cancers. Int. J. Mol. Sci. 2019, 20, 3926. [Google Scholar] [CrossRef] [PubMed]
- Lunde, H.M.B.; Assmus, J.; Myhr, K.-M.; Bø, L.; Grytten, N. Survival and Cause of Death in Multiple Sclerosis: A 60-Year Longitudinal Population Study. J. Neurol. Neurosurg. Psychiatry 2017, 88, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, G.G.; Pacheco-Moisés, F.P.; Macías-Islas, M.Á.; Flores-Alvarado, L.J.; Mireles-Ramírez, M.A.; González-Renovato, E.D.; Hernández-Navarro, V.E.; Sánchez-López, A.L.; Alatorre-Jiménez, M.A. Role of the Blood-Brain Barrier in Multiple Sclerosis. Arch. Med. Res. 2014, 45, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.H.; Miller, S.L.; Castillo-Melendez, M.; Malhotra, A. The Neurovascular Unit: Effects of Brain Insults During the Perinatal Period. Front. Neurosci. 2019, 13, 1452. [Google Scholar] [CrossRef] [PubMed]
- Persidsky, Y.; Ramirez, S.H.; Haorah, J.; Kanmogne, G.D. Blood-Brain Barrier: Structural Components and Function under Physiologic and Pathologic Conditions. J. Neuroimmune Pharmacol. Off. J. Soc. NeuroImmune Pharmacol. 2006, 1, 223–236. [Google Scholar] [CrossRef]
- Kadry, H.; Noorani, B.; Cucullo, L. A Blood-Brain Barrier Overview on Structure, Function, Impairment, and Biomarkers of Integrity. Fluids Barriers CNS 2020, 17, 69. [Google Scholar] [CrossRef]
- Bazzoni, G.; Dejana, E. Endothelial Cell-to-Cell Junctions: Molecular Organization and Role in Vascular Homeostasis. Physiol. Rev. 2004, 84, 869–901. [Google Scholar] [CrossRef]
- Greene, C.; Hanley, N.; Campbell, M. Claudin-5: Gatekeeper of Neurological Function. Fluids Barriers CNS 2019, 16, 3. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Campbell, M.; Tachibana, K.; Okada, Y.; Kondoh, M. Claudin-5: A Pharmacological Target to Modify the Permeability of the Blood-Brain Barrier. Biol. Pharm. Bull. 2021, 44, 1380–1390. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Tsai, H.-H.; Hoi, K.K.; Huang, N.; Yu, G.; Kim, K.; Baranzini, S.E.; Xiao, L.; Chan, J.R.; Fancy, S.P.J. Aberrant Oligodendroglial-Vascular Interactions Disrupt the Blood Brain Barrier Triggering CNS Inflammation. Nat. Neurosci. 2019, 22, 709–718. [Google Scholar] [CrossRef]
- Feldman, G.J.; Mullin, J.M.; Ryan, M.P. Occludin: Structure, Function and Regulation. Adv. Drug Deliv. Rev. 2005, 57, 883–917. [Google Scholar] [CrossRef] [PubMed]
- Cummins, P.M. Occludin: One Protein, Many Forms. Mol. Cell. Biol. 2012, 32, 242–250. [Google Scholar] [CrossRef]
- Sugiyama, S.; Sasaki, T.; Tanaka, H.; Yan, H.; Ikegami, T.; Kanki, H.; Nishiyama, K.; Beck, G.; Gon, Y.; Okazaki, S.; et al. The Tight Junction Protein Occludin Modulates Blood-Brain Barrier Integrity and Neurological Function after Ischemic Stroke in Mice. Sci. Rep. 2023, 13, 2892. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, C.; Schwietzer, Y.A.; Otani, T.; Furuse, M.; Ebnet, K. Physiological Functions of Junctional Adhesion Molecules (JAMs) in Tight Junctions. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183299. [Google Scholar] [CrossRef]
- Duong, C.N.; Vestweber, D. Mechanisms Ensuring Endothelial Junction Integrity Beyond VE-Cadherin. Front. Physiol. 2020, 11, 519. [Google Scholar] [CrossRef]
- Ippolitov, D.; Arreza, L.; Munir, M.N.; Hombach-Klonisch, S. Brain Microvascular Pericytes-More than Bystanders in Breast Cancer Brain Metastasis. Cells 2022, 11, 1263. [Google Scholar] [CrossRef]
- Kim, Y.; Park, J.; Choi, Y.K. The Role of Astrocytes in the Central Nervous System Focused on BK Channel and Heme Oxygenase Metabolites: A Review. Antioxidants 2019, 8, 121. [Google Scholar] [CrossRef]
- Cabezas, R.; Avila, M.; Gonzalez, J.; El-Bachá, R.S.; Báez, E.; García-Segura, L.M.; Jurado Coronel, J.C.; Capani, F.; Cardona-Gomez, G.P.; Barreto, G.E. Astrocytic Modulation of Blood Brain Barrier: Perspectives on Parkinson’s Disease. Front. Cell. Neurosci. 2014, 8, 211. [Google Scholar] [CrossRef]
- Dominguez, R.; Holmes, K.C. Actin Structure and Function. Annu. Rev. Biophys. 2011, 40, 169–186. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Dominguez, R. Regulation of Actin Cytoskeleton Dynamics in Cells. Mol. Cells 2010, 29, 311–325. [Google Scholar] [CrossRef] [PubMed]
- Belvitch, P.; Htwe, Y.M.; Brown, M.E.; Dudek, S. Cortical Actin Dynamics in Endothelial Permeability. Curr. Top. Membr. 2018, 82, 141–195. [Google Scholar] [CrossRef] [PubMed]
- Lampugnani, M.G.; Zanetti, A.; Breviario, F.; Balconi, G.; Orsenigo, F.; Corada, M.; Spagnuolo, R.; Betson, M.; Braga, V.; Dejana, E. VE-Cadherin Regulates Endothelial Actin Activating Rac and Increasing Membrane Association of Tiam. Mol. Biol. Cell 2002, 13, 1175–1189. [Google Scholar] [CrossRef] [PubMed]
- Alieva, I.B. Role of Microtubule Cytoskeleton in Regulation of Endothelial Barrier Function. Biochem. Biokhimiia 2014, 79, 964–975. [Google Scholar] [CrossRef] [PubMed]
- Karki, P.; Birukova, A.A. Microtubules as Major Regulators of Endothelial Function: Implication for Lung Injury. Front. Physiol. 2021, 12, 758313. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, H.; Aebi, U. Intermediate Filaments: Structure and Assembly. Cold Spring Harb. Perspect. Biol. 2016, 8, a018242. [Google Scholar] [CrossRef] [PubMed]
- Pogoda, K.; Janmey, P.A. Transmit and Protect: The Mechanical Functions of Intermediate Filaments. Curr. Opin. Cell Biol. 2023, 85, 102281. [Google Scholar] [CrossRef] [PubMed]
- Ridge, K.M.; Eriksson, J.E.; Pekny, M.; Goldman, R.D. Roles of Vimentin in Health and Disease. Genes Dev. 2022, 36, 391–407. [Google Scholar] [CrossRef]
- Alahmari, A. Blood-Brain Barrier Overview: Structural and Functional Correlation. Neural Plast. 2021, 2021, 6564585. [Google Scholar] [CrossRef]
- Singh, A.V.; Chandrasekar, V.; Laux, P.; Luch, A.; Dakua, S.P.; Zamboni, P.; Shelar, A.; Yang, Y.; Pandit, V.; Tisato, V.; et al. Micropatterned Neurovascular Interface to Mimic the Blood-Brain Barrier’s Neurophysiology and Micromechanical Function: A BBB-on-CHIP Model. Cells 2022, 11, 2801. [Google Scholar] [CrossRef] [PubMed]
- Rathi, S.; Reche, A.; Dhamdhere, N.; Bolenwar, A. Perspectives on the Application of Nanomaterials in Medical and Dental Practices. Cureus 2023, 15, e43565. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, L.; Carinci, F.; Martini, M.; Gemmati, D.; Nardone, M.; Siciliani, G. Quantitive Evaluation of Dentin Sialoprotein (DSP) Using Microbeads—A Potential Early Marker of Root Resorption. ORAL Implantol. 2016, 9, 132–142. [Google Scholar] [CrossRef]
- Balasa, R.; Barcutean, L.; Mosora, O.; Manu, D. Reviewing the Significance of Blood-Brain Barrier Disruption in Multiple Sclerosis Pathology and Treatment. Int. J. Mol. Sci. 2021, 22, 8370. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Shen, L. Advances and Trends in Omics Technology Development. Front. Med. 2022, 9, 911861. [Google Scholar] [CrossRef] [PubMed]
- Nishanth, K.; Tariq, E.; Nzvere, F.P.; Miqdad, M.; Cancarevic, I. Role of Smoking in the Pathogenesis of Multiple Sclerosis: A Review Article. Cureus 2020, 12, e9564. [Google Scholar] [CrossRef] [PubMed]
- Altieri, C.; Speranza, B.; Corbo, M.R.; Sinigaglia, M.; Bevilacqua, A. Gut-Microbiota, and Multiple Sclerosis: Background, Evidence, and Perspectives. Nutrients 2023, 15, 942. [Google Scholar] [CrossRef] [PubMed]
- Correale, J.; Hohlfeld, R.; Baranzini, S.E. The Role of the Gut Microbiota in Multiple Sclerosis. Nat. Rev. Neurol. 2022, 18, 544–558. [Google Scholar] [CrossRef] [PubMed]
- Cotsapas, C.; Mitrovic, M. Genome-wide Association Studies of Multiple Sclerosis. Clin. Transl. Immunol. 2018, 7, e1018. [Google Scholar] [CrossRef]
- Ziliotto, N.; Marchetti, G.; Scapoli, C.; Bovolenta, M.; Meneghetti, S.; Benazzo, A.; Lunghi, B.; Balestra, D.; Laino, L.A.; Bozzini, N.; et al. C6orf10 Low-Frequency and Rare Variants in Italian Multiple Sclerosis Patients. Front. Genet. 2019, 10, 573. [Google Scholar] [CrossRef]
- Paraboschi, E.M.; Rimoldi, V.; Soldà, G.; Tabaglio, T.; Dall’Osso, C.; Saba, E.; Vigliano, M.; Salviati, A.; Leone, M.; Benedetti, M.D.; et al. Functional Variations Modulating PRKCA Expression and Alternative Splicing Predispose to Multiple Sclerosis. Hum. Mol. Genet. 2014, 23, 6746–6761. [Google Scholar] [CrossRef] [PubMed]
- Paraboschi, E.M.; Cardamone, G.; Rimoldi, V.; Gemmati, D.; Spreafico, M.; Duga, S.; Soldà, G.; Asselta, R. Meta-Analysis of Multiple Sclerosis Microarray Data Reveals Dysregulation in RNA Splicing Regulatory Genes. Int. J. Mol. Sci. 2015, 16, 23463–23481. [Google Scholar] [CrossRef] [PubMed]
- Gemmati, D.; Zeri, G.; Orioli, E.; De Gaetano, F.E.; Salvi, F.; Bartolomei, I.; D’Alfonso, S.; Dall’osso, C.; Leone, M.A.; Singh, A.V.; et al. Polymorphisms in the Genes Coding for Iron Binding and Transporting Proteins Are Associated with Disability, Severity, and Early Progression in Multiple Sclerosis. BMC Med. Genet. 2012, 13, 70. [Google Scholar] [CrossRef]
- Ferlini, A.; Bovolenta, M.; Neri, M.; Gualandi, F.; Balboni, A.; Yuryev, A.; Salvi, F.; Gemmati, D.; Liboni, A.; Zamboni, P. Custom CGH Array Profiling of Copy Number Variations (CNVs) on Chromosome 6p21.32 (HLA Locus) in Patients with Venous Malformations Associated with Multiple Sclerosis. BMC Med. Genet. 2010, 11, 64. [Google Scholar] [CrossRef] [PubMed]
- Knox, E.G.; Aburto, M.R.; Clarke, G.; Cryan, J.F.; O’Driscoll, C.M. The Blood-Brain Barrier in Aging and Neurodegeneration. Mol. Psychiatry 2022, 27, 2659–2673. [Google Scholar] [CrossRef] [PubMed]
- Nishihara, H.; Perriot, S.; Gastfriend, B.D.; Steinfort, M.; Cibien, C.; Soldati, S.; Matsuo, K.; Guimbal, S.; Mathias, A.; Palecek, S.P.; et al. Intrinsic Blood-Brain Barrier Dysfunction Contributes to Multiple Sclerosis Pathogenesis. Brain J. Neurol. 2022, 145, 4334–4348. [Google Scholar] [CrossRef] [PubMed]
- Salimi, H.; Klein, R.S. Disruption of the Blood-Brain Barrier During Neuroinflammatory and Neuroinfectious Diseases. Neuroimmune Dis. 2019, 195–234. [Google Scholar] [CrossRef]
- Luchicchi, A.; Preziosa, P.; ’t Hart, B. Editorial: “Inside-Out” vs “Outside-In” Paradigms in Multiple Sclerosis Etiopathogenesis. Front. Cell. Neurosci. 2021, 15, 666529. [Google Scholar] [CrossRef]
- Boscia, F.; Elkjaer, M.L.; Illes, Z.; Kukley, M. Altered Expression of Ion Channels in White Matter Lesions of Progressive Multiple Sclerosis: What Do We Know About Their Function? Front. Cell. Neurosci. 2021, 15, 685703. [Google Scholar] [CrossRef]
- Plantone, D.; Inglese, M.; Salvetti, M.; Koudriavtseva, T. A Perspective of Coagulation Dysfunction in Multiple Sclerosis and in Experimental Allergic Encephalomyelitis. Front. Neurol. 2019, 9, 1175. [Google Scholar] [CrossRef]
- Yang, S.; Chen, Y.; Deng, X.; Jiang, W.; Li, B.; Fu, Z.; Du, M.; Ding, R. Hemoglobin-Induced Nitric Oxide Synthase Overexpression and Nitric Oxide Production Contribute to Blood-Brain Barrier Disruption in the Rat. J. Mol. Neurosci. MN 2013, 51, 352–363. [Google Scholar] [CrossRef]
- Saez-Calveras, N.; Stuve, O. The Role of the Complement System in Multiple Sclerosis: A Review. Front. Immunol. 2022, 13, 970486. [Google Scholar] [CrossRef] [PubMed]
- Tisato, V.; Castiglione, A.; Ciorba, A.; Aimoni, C.; Silva, J.A.; Gallo, I.; D’Aversa, E.; Salvatori, F.; Bianchini, C.; Pelucchi, S.; et al. LINE-1 Global DNA Methylation, Iron Homeostasis Genes, Sex and Age in Sudden Sensorineural Hearing Loss (SSNHL). Hum. Genomics 2023, 17, 112. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Pang, X.; Yeo, A.J.; Xie, S.; Xiang, M.; Shi, B.; Yu, G.; Li, C. The Molecular Mechanisms of Ferroptosis and Its Role in Blood-Brain Barrier Dysfunction. Front. Cell. Neurosci. 2022, 16, 889765. [Google Scholar] [CrossRef] [PubMed]
- Van San, E.; Debruyne, A.C.; Veeckmans, G.; Tyurina, Y.Y.; Tyurin, V.A.; Zheng, H.; Choi, S.M.; Augustyns, K.; van Loo, G.; Michalke, B.; et al. Ferroptosis Contributes to Multiple Sclerosis and Its Pharmacological Targeting Suppresses Experimental Disease Progression. Cell Death Differ. 2023, 30, 2092–2103. [Google Scholar] [CrossRef] [PubMed]
- Sheykhansari, S.; Kozielski, K.; Bill, J.; Sitti, M.; Gemmati, D.; Zamboni, P.; Singh, A.V. Redox Metals Homeostasis in Multiple Sclerosis and Amyotrophic Lateral Sclerosis: A Review. Cell Death Dis. 2018, 9, 348. [Google Scholar] [CrossRef] [PubMed]
- Klistorner, A.; Wang, C.; Yiannikas, C.; Parratt, J.; Dwyer, M.; Barton, J.; Graham, S.L.; You, Y.; Liu, S.; Barnett, M.H. Evidence of Progressive Tissue Loss in the Core of Chronic MS Lesions: A Longitudinal DTI Study. NeuroImage Clin. 2018, 17, 1028–1035. [Google Scholar] [CrossRef] [PubMed]
- Mills, E.; Dong, X.; Wang, F.; Xu, H. Mechanisms of Brain Iron Transport: Insight into Neurodegeneration and CNS Disorders. Future Med. Chem. 2010, 2, 51. [Google Scholar] [CrossRef]
- Tisato, V.; Zuliani, G.; Vigliano, M.; Longo, G.; Franchini, E.; Secchiero, P.; Zauli, G.; Paraboschi, E.M.; Vikram Singh, A.; Serino, M.L.; et al. Gene-Gene Interactions among Coding Genes of Iron-Homeostasis Proteins and APOE-Alleles in Cognitive Impairment Diseases. PLoS ONE 2018, 13, e0193867. [Google Scholar] [CrossRef]
- de Barcelos, I.P.; Troxell, R.M.; Graves, J.S. Mitochondrial Dysfunction and Multiple Sclerosis. Biology 2019, 8, 37. [Google Scholar] [CrossRef]
- Connor, J.R.; Menzies, S.L. Relationship of Iron to Oligodendrocytes and Myelination. Glia 1996, 17, 83–93. [Google Scholar] [CrossRef]
- Tang, C.; Yang, J.; Zhu, C.; Ding, Y.; Yang, S.; Xu, B.; He, D. Iron Metabolism Disorder and Multiple Sclerosis: A Comprehensive Analysis. Front. Immunol. 2024, 15, 1376838. [Google Scholar] [CrossRef] [PubMed]
- van Rensburg, S.J.; Peeters, A.V.; van Toorn, R.; Schoeman, J.; Moremi, K.E.; van Heerden, C.J.; Kotze, M.J. Identification of an Iron-Responsive Subtype in Two Children Diagnosed with Relapsing-Remitting Multiple Sclerosis Using Whole Exome Sequencing. Mol. Genet. Metab. Rep. 2019, 19, 100465. [Google Scholar] [CrossRef] [PubMed]
- Haider, L. Inflammation, Iron, Energy Failure, and Oxidative Stress in the Pathogenesis of Multiple Sclerosis. Oxid. Med. Cell. Longev. 2015, 2015, 725370. [Google Scholar] [CrossRef]
- Cherchi, F.; Bulli, I.; Venturini, M.; Pugliese, A.M.; Coppi, E. Ion Channels as New Attractive Targets to Improve Re-Myelination Processes in the Brain. Int. J. Mol. Sci. 2021, 22, 7277. [Google Scholar] [CrossRef] [PubMed]
- Zündorf, G.; Reiser, G. Calcium Dysregulation and Homeostasis of Neural Calcium in the Molecular Mechanisms of Neurodegenerative Diseases Provide Multiple Targets for Neuroprotection. Antioxid. Redox Signal. 2011, 14, 1275–1288. [Google Scholar] [CrossRef] [PubMed]
- Gemmati, D.; Vigliano, M.; Burini, F.; Mari, R.; El Mohsein, H.H.A.; Parmeggiani, F.; Serino, M.L. Coagulation Factor XIIIA (F13A1): Novel Perspectives in Treatment and Pharmacogenetics. Curr. Pharm. Des. 2016, 22, 1449–1459. [Google Scholar] [CrossRef] [PubMed]
- Alruwaili, M.; Al-Kuraishy, H.M.; Alexiou, A.; Papadakis, M.; ALRashdi, B.M.; Elhussieny, O.; Saad, H.M.; Batiha, G.E.-S. Pathogenic Role of Fibrinogen in the Neuropathology of Multiple Sclerosis: A Tale of Sorrows and Fears. Neurochem. Res. 2023, 48, 3255–3269. [Google Scholar] [CrossRef] [PubMed]
- Gemmati, D.; Zeri, G.; Orioli, E.; Mari, R.; Moratelli, S.; Vigliano, M.; Marchesini, J.; Grossi, M.E.; Pecoraro, A.; Cuneo, A.; et al. Factor XIII-A Dynamics in Acute Myocardial Infarction: A Novel Prognostic Biomarker? Thromb. Haemost. 2015, 114, 123–132. [Google Scholar] [CrossRef]
- Zamboni, P.; De Mattei, M.; Ongaro, A.; Fogato, L.; Carandina, S.; De Palma, M.; Tognazzo, S.; Scapoli, G.L.; Serino, M.L.; Caruso, A.; et al. Factor XIII Contrasts the Effects of Metalloproteinases in Human Dermal Fibroblast Cultured Cells. Vasc. Endovascular Surg. 2004, 38, 431–438. [Google Scholar] [CrossRef]
- Noll, T.; Wozniak, G. Factor XIII and endothelial barrier function. Hamostaseologie 2002, 22, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Noll, T.; Wozniak, G.; McCarson, K.; Hajimohammad, A.; Metzner, H.J.; Inserte, J.; Kummer, W.; Hehrlein, F.W.; Piper, H.M. Effect of Factor XIII on Endothelial Barrier Function. J. Exp. Med. 1999, 189, 1373–1382. [Google Scholar] [CrossRef] [PubMed]
- Brailoiu, E.; Shipsky, M.M.; Yan, G.; Abood, M.E.; Brailoiu, G.C. Mechanisms of Modulation of Brain Microvascular Endothelial Cells Function by Thrombin. Brain Res. 2017, 1657, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Gan, L.; Ren, L.; Lin, Y.; Ma, C.; Lin, X. Factors Influencing the Blood-Brain Barrier Permeability. Brain Res. 2022, 1788, 147937. [Google Scholar] [CrossRef]
- Reiche, J.; Huber, O. Post-Translational Modifications of Tight Junction Transmembrane Proteins and Their Direct Effect on Barrier Function. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183330. [Google Scholar] [CrossRef] [PubMed]
- Campos, S.B.; Ashworth, S.L.; Wean, S.; Hosford, M.; Sandoval, R.M.; Hallett, M.A.; Atkinson, S.J.; Molitoris, B.A. Cytokine-Induced F-Actin Reorganization in Endothelial Cells Involves RhoA Activation. Am. J. Physiol.-Ren. Physiol. 2009, 296, F487–F495. [Google Scholar] [CrossRef]
- Alvarez, J.I.; Cayrol, R.; Prat, A. Disruption of Central Nervous System Barriers in Multiple Sclerosis. Biochim. Biophys. Acta 2011, 1812, 252–264. [Google Scholar] [CrossRef] [PubMed]
- Plumb, J.; McQuaid, S.; Mirakhur, M.; Kirk, J. Abnormal Endothelial Tight Junctions in Active Lesions and Normal-Appearing White Matter in Multiple Sclerosis. Brain Pathol. Zurich Switz. 2002, 12, 154–169. [Google Scholar] [CrossRef]
- Negri, S.; Faris, P.; Moccia, F. Reactive Oxygen Species and Endothelial Ca2+ Signaling: Brothers in Arms or Partners in Crime? Int. J. Mol. Sci. 2021, 22, 9821. [Google Scholar] [CrossRef]
- Ramakrishna, K.; Nalla, L.V.; Naresh, D.; Venkateswarlu, K.; Viswanadh, M.K.; Nalluri, B.N.; Chakravarthy, G.; Duguluri, S.; Singh, P.; Rai, S.N.; et al. WNT-β Catenin Signaling as a Potential Therapeutic Target for Neurodegenerative Diseases: Current Status and Future Perspective. Diseases 2023, 11, 89. [Google Scholar] [CrossRef]
- Hussain, B.; Fang, C.; Huang, X.; Feng, Z.; Yao, Y.; Wang, Y.; Chang, J. Endothelial β-Catenin Deficiency Causes Blood-Brain Barrier Breakdown via Enhancing the Paracellular and Transcellular Permeability. Front. Mol. Neurosci. 2022, 15, 895429. [Google Scholar] [CrossRef] [PubMed]
- Hamill, K.J.; Hiroyasu, S.; Colburn, Z.T.; Ventrella, R.V.; Hopkinson, S.B.; Skalli, O.; Jones, J.C.R. Alpha Actinin-1 Regulates Cell-Matrix Adhesion Organization in Keratinocytes: Consequences for Skin Cell Motility. J. Investig. Dermatol. 2015, 135, 1043–1052. [Google Scholar] [CrossRef]
- Pandey, S.; Dioni, I.; Lambardi, D.; Real-Fernandez, F.; Peroni, E.; Pacini, G.; Lolli, F.; Seraglia, R.; Papini, A.M.; Rovero, P. Alpha Actinin Is Specifically Recognized by Multiple Sclerosis Autoantibodies Isolated Using an N-Glucosylated Peptide Epitope. Mol. Cell. Proteomics MCP 2013, 12, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Padden, M.; Leech, S.; Craig, B.; Kirk, J.; Brankin, B.; McQuaid, S. Differences in Expression of Junctional Adhesion Molecule-A and Beta-Catenin in Multiple Sclerosis Brain Tissue: Increasing Evidence for the Role of Tight Junction Pathology. Acta Neuropathol. 2007, 113, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Stamatovic, S.M.; Sladojevic, N.; Keep, R.F.; Andjelkovic, A.V. Relocalization of Junctional Adhesion Molecule A during Inflammatory Stimulation of Brain Endothelial Cells. Mol. Cell. Biol. 2012, 32, 3414–3427. [Google Scholar] [CrossRef] [PubMed]
- Gregor, M.; Osmanagic-Myers, S.; Burgstaller, G.; Wolfram, M.; Fischer, I.; Walko, G.; Resch, G.P.; Jörgl, A.; Herrmann, H.; Wiche, G. Mechanosensing through Focal Adhesion-Anchored Intermediate Filaments. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2014, 28, 715–729. [Google Scholar] [CrossRef] [PubMed]
- Jiu, Y.; Peränen, J.; Schaible, N.; Cheng, F.; Eriksson, J.E.; Krishnan, R.; Lappalainen, P. Vimentin Intermediate Filaments Control Actin Stress Fiber Assembly through GEF-H1 and RhoA. J. Cell Sci. 2017, 130, 892–902. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Ghamloush, M.M.; Aldawood, A.; Warburton, R.; Toksoz, D.; Hill, N.S.; Tang, D.D.; Kayyali, U.S. Modulating Endothelial Barrier Function by Targeting Vimentin Phosphorylation. J. Cell. Physiol. 2014, 229, 1484–1493. [Google Scholar] [CrossRef]
- Fakih, R.; Diaz-Cruz, C.; Chua, A.S.; Gonzalez, C.; Healy, B.C.; Sattarnezhad, N.; Glanz, B.I.; Weiner, H.L.; Chitnis, T. Food Allergies Are Associated with Increased Disease Activity in Multiple Sclerosis. J. Neurol. Neurosurg. Psychiatry 2019, 90, 629–635. [Google Scholar] [CrossRef]
- Pedotti, R.; DeVoss, J.J.; Youssef, S.; Mitchell, D.; Wedemeyer, J.; Madanat, R.; Garren, H.; Fontoura, P.; Tsai, M.; Galli, S.J.; et al. Multiple Elements of the Allergic Arm of the Immune Response Modulate Autoimmune Demyelination. Proc. Natl. Acad. Sci. USA 2003, 100, 1867–1872. [Google Scholar] [CrossRef]
- Naddafi, F.; Mirshafiey, A. The Neglected Role of Histamine in Alzheimer’s Disease. Am. J. Alzheimers Dis. Other Demen. 2013, 28, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Jadidi-Niaragh, F.; Mirshafiey, A. Histamine and Histamine Receptors in Pathogenesis and Treatment of Multiple Sclerosis. Neuropharmacology 2010, 59, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Shasby, D.M.; Ries, D.R.; Shasby, S.S.; Winter, M.C. Histamine Stimulates Phosphorylation of Adherens Junction Proteins and Alters Their Link to Vimentin. Am. J. Physiol. Lung Cell. Mol. Physiol. 2002, 282, L1330–L1338. [Google Scholar] [CrossRef] [PubMed]
- Dorado, G.; Gálvez, S.; Rosales, T.E.; Vásquez, V.F.; Hernández, P. Analyzing Modern Biomolecules: The Revolution of Nucleic-Acid Sequencing—Review. Biomolecules 2021, 11, 1111. [Google Scholar] [CrossRef] [PubMed]
- You, X.; Vlatkovic, I.; Babic, A.; Will, T.; Epstein, I.; Tushev, G.; Akbalik, G.; Wang, M.; Glock, C.; Quedenau, C.; et al. Neural Circular RNAs Are Derived from Synaptic Genes and Regulated by Development and Plasticity. Nat. Neurosci. 2015, 18, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Bai, Y.; Liao, Z.; Gritsch, D.; Liu, X.; Wang, T.; Borges-Monroy, R.; Ehrlich, A.; Serano, G.E.; Feany, M.B.; et al. Circular RNAs in the Human Brain Are Tailored to Neuron Identity and Neuropsychiatric Disease. BioRxiv Prepr. Serv. Biol. 2023, 14, 5327. [Google Scholar] [CrossRef] [PubMed]
- Sekar, S.; Liang, W.S. Circular RNA Expression and Function in the Brain. Non-Coding RNA Res. 2019, 4, 23–29. [Google Scholar] [CrossRef]
- Zhou, M.; Li, S.; Huang, C. Physiological and Pathological Functions of Circular RNAs in the Nervous System. Neural Regen. Res. 2023, 19, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Liu, H.; Chang, N.; Fu, W.; Guo, Z.; Wang, Y. Circular RNAs: New Players Involved in the Regulation of Cognition and Cognitive Diseases. Front. Neurosci. 2023, 17, 1097878. [Google Scholar] [CrossRef]
- Cardamone, G.; Paraboschi, E.M.; Soldà, G.; Cantoni, C.; Supino, D.; Piccio, L.; Duga, S.; Asselta, R. Not Only Cancer: The Long Non-Coding RNA MALAT1 Affects the Repertoire of Alternatively Spliced Transcripts and Circular RNAs in Multiple Sclerosis. Hum. Mol. Genet. 2019, 28, 1414–1428. [Google Scholar] [CrossRef]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs Are a Large Class of Animal RNAs with Regulatory Potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Yuan, X.; Cai, Q.; Tang, C.; Gao, J. Circular RNA as an Epigenetic Regulator in Chronic Liver Diseases. Cells 2021, 10, 1945. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Joseph, S.; Xia, M.; Teng, F.; Chen, X.; Huang, R.; Zhai, L.; Deng, W. Circular RNAs Acting as miRNAs’ Sponges and Their Roles in Stem Cells. J. Clin. Med. 2022, 11, 2909. [Google Scholar] [CrossRef]
- Verduci, L.; Tarcitano, E.; Strano, S.; Yarden, Y.; Blandino, G. CircRNAs: Role in Human Diseases and Potential Use as Biomarkers. Cell Death Dis. 2021, 12, 468. [Google Scholar] [CrossRef] [PubMed]
- Salvatori, F.; D’Aversa, E.; Serino, M.L.; Singh, A.V.; Secchiero, P.; Zauli, G.; Tisato, V.; Gemmati, D. miRNAs Epigenetic Tuning of Wall Remodeling in the Early Phase after Myocardial Infarction: A Novel Epidrug Approach. Int. J. Mol. Sci. 2023, 24, 13268. [Google Scholar] [CrossRef] [PubMed]
- He, A.T.; Liu, J.; Li, F.; Yang, B.B. Targeting Circular RNAs as a Therapeutic Approach: Current Strategies and Challenges. Signal Transduct. Target. Ther. 2021, 6, 185. [Google Scholar] [CrossRef] [PubMed]
- Pisignano, G.; Michael, D.C.; Visal, T.H.; Pirlog, R.; Ladomery, M.; Calin, G.A. Going Circular: History, Present, and Future of circRNAs in Cancer. Oncogene 2023, 42, 2783. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhang, J.; Zheng, X.; Pan, Z.; Zhao, F.; Gao, Y. CIRI-Deep Enables Single-Cell and Spatial Transcriptomic Analysis of Circular RNAs with Deep Learning. Adv. Sci. Weinh. Baden-Wurtt. Ger. 2024, 11, e2308115. [Google Scholar] [CrossRef]
- Piwecka, M.; Rajewsky, N.; Rybak-Wolf, A. Single-Cell and Spatial Transcriptomics: Deciphering Brain Complexity in Health and Disease. Nat. Rev. Neurol. 2023, 19, 346–362. [Google Scholar] [CrossRef]
- Huang, A.; Zheng, H.; Wu, Z.; Chen, M.; Huang, Y. Circular RNA-Protein Interactions: Functions, Mechanisms, and Identification. Theranostics 2020, 10, 3503–3517. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, C.; Du, Y.; Li, Z.; Li, M.; Hou, P.; Shen, Z.; Chu, S.; Zheng, J.; Bai, J. Expanding Uncapped Translation and Emerging Function of Circular RNA in Carcinomas and Noncarcinomas. Mol. Cancer 2022, 21, 13. [Google Scholar] [CrossRef] [PubMed]
- Lindner, G.; Takenaka, K.; Santucci, K.; Gao, Y.; Janitz, M. Protein-Coding Circular RNAs—Mechanism, Detection, and Their Role in Cancer and Neurodegenerative Diseases. Biochem. Biophys. Res. Commun. 2023, 678, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Cardamone, G.; Paraboschi, E.M.; Soldà, G.; Liberatore, G.; Rimoldi, V.; Cibella, J.; Airi, F.; Tisato, V.; Cantoni, C.; Gallia, F.; et al. The Circular RNA Landscape in Multiple Sclerosis: Disease-Specific Associated Variants and Exon Methylation Shape Circular RNA Expression Profile. Mult. Scler. Relat. Disord. 2023, 69, 104426. [Google Scholar] [CrossRef]
- Sciaccotta, R.; Murdaca, G.; Caserta, S.; Rizzo, V.; Gangemi, S.; Allegra, A. Circular RNAs: A New Approach to Multiple Sclerosis. Biomedicines 2023, 11, 2883. [Google Scholar] [CrossRef] [PubMed]
- D’Anca, M.; Buccellato, F.R.; Fenoglio, C.; Galimberti, D. Circular RNAs: Emblematic Players of Neurogenesis and Neurodegeneration. Int. J. Mol. Sci. 2022, 23, 4134. [Google Scholar] [CrossRef]
- Paraboschi, E.M.; Cardamone, G.; Soldà, G.; Duga, S.; Asselta, R. Interpreting Non-Coding Genetic Variation in Multiple Sclerosis Genome-Wide Associated Regions. Front. Genet. 2018, 9, 647. [Google Scholar] [CrossRef] [PubMed]
- Cardamone, G.; Paraboschi, E.M.; Rimoldi, V.; Duga, S.; Soldà, G.; Asselta, R. The Characterization of GSDMB Splicing and Backsplicing Profiles Identifies Novel Isoforms and a Circular RNA That Are Dysregulated in Multiple Sclerosis. Int. J. Mol. Sci. 2017, 18, 576. [Google Scholar] [CrossRef]
- Hoque, P.; Romero, B.; Akins, R.E.; Batish, M. Exploring the Multifaceted Biologically Relevant Roles of circRNAs: From Regulation, Translation to Biomarkers. Cells 2023, 12, 2813. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Song, F.; Yang, Q.; Zhou, Y.; Shao, C.; Shen, Y.; Zhao, Z.; Tang, Q.; Hou, Y.; Xie, J. Characterization of Tissue-Specific Biomarkers with the Expression of circRNAs in Forensically Relevant Body Fluids. Int. J. Legal Med. 2019, 133, 1321–1331. [Google Scholar] [CrossRef]
- Eslahi, M.; Nematbakhsh, N.; Dastmalchi, N.; Teimourian, S.; Safaralizadeh, R. An Updated Review of Epigenetic-Related Mechanisms and Their Contribution to Multiple Sclerosis Disease. CNS Neurol. Disord. Drug Targets 2023, 22, 381–393. [Google Scholar] [CrossRef]
- Munger, K.; Ascherio, A. Understanding the Joint Effects of EBV and Vitamin D in MS. Mult. Scler. Houndmills Basingstoke Engl. 2013, 19, 1554–1555. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, E.M. Circular RNA in Multiple Sclerosis: Pathogenicity and Potential Biomarker Development: A Systematic Review. Epigenetics Insights 2023, 16, 25168657231213195. [Google Scholar] [CrossRef]
- Yang, R.; Chen, J.; Xu, B.; Yang, B.; Fu, J.; Xiao, S.; Tan, C.; Chen, H.; Wang, X. Circ_2858 Helps Blood-Brain Barrier Disruption by Increasing VEGFA via Sponging miR-93-5p during Escherichia Coli Meningitis. Mol. Ther. Nucleic Acids 2020, 22, 708–721. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Huang, C.; Wen, X.; Liu, W.; Huang, X.; Li, Y.; Zang, J.; Weng, Z.; Lu, D.; Tsang, C.K.; et al. Circular RNA Circ-FoxO3 Attenuates Blood-Brain Barrier Damage by Inducing Autophagy during Ischemia/Reperfusion. Mol. Ther. J. Am. Soc. Gene Ther. 2022, 30, 1275–1287. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zeng, H.; Lei, L.; Tong, X.; Yang, L.; Yang, Y.; Li, S.; Zhou, Y.; Luo, L.; Huang, J.; et al. Tight Junctions and Their Regulation by Non-Coding RNAs. Int. J. Biol. Sci. 2021, 17, 712–727. [Google Scholar] [CrossRef]
- Nelles, D.G.; Hazrati, L.-N. Ependymal Cells and Neurodegenerative Disease: Outcomes of Compromised Ependymal Barrier Function. Brain Commun. 2022, 4, fcac288. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.; Chai, Y.; Sun, Y.; Guo, Z.; Wang, X.; Wang, Z.; Wang, Z.; Wang, Z. Hsa_circ_0074158 Regulates the Endothelial Barrier Function in Sepsis and Its Potential Value as a Biomarker. Front. Genet. 2022, 13, 1002344. [Google Scholar] [CrossRef] [PubMed]
- Kremerskothen, J.; Teber, I.; Wendholt, D.; Liedtke, T.; Böckers, T.M.; Barnekow, A. Brain-Specific Splicing of Alpha-Actinin 1 (ACTN1) mRNA. Biochem. Biophys. Res. Commun. 2002, 295, 678–681. [Google Scholar] [CrossRef] [PubMed]
- Le, S.; Hu, X.; Yao, M.; Chen, H.; Yu, M.; Xu, X.; Nakazawa, N.; Margadant, F.M.; Sheetz, M.P.; Yan, J. Mechanotransmission and Mechanosensing of Human Alpha-Actinin 1. Cell Rep. 2017, 21, 2714–2723. [Google Scholar] [CrossRef]
- Bai, Y.; Zhang, Y.; Han, B.; Yang, L.; Chen, X.; Huang, R.; Wu, F.; Chao, J.; Liu, P.; Hu, G.; et al. Circular RNA DLGAP4 Ameliorates Ischemic Stroke Outcomes by Targeting miR-143 to Regulate Endothelial-Mesenchymal Transition Associated with Blood–Brain Barrier Integrity. J. Neurosci. 2018, 38, 32–50. [Google Scholar] [CrossRef]
- Feng, Z.; Zhang, L.; Wang, S.; Hong, Q. Circular RNA circDLGAP4 Exerts Neuroprotective Effects via Modulating miR-134-5p/CREB Pathway in Parkinson’s Disease. Biochem. Biophys. Res. Commun. 2020, 522, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wu, P.; Ma, Y.; Xue, Y.; Liu, Y.; Zheng, J.; Liu, X.; He, Q.; Ma, J.; Liu, L.; et al. Circular RNA USP1 Regulates the Permeability of Blood-Tumour Barrier via miR-194-5p/FLI1 Axis. J. Cell. Mol. Med. 2020, 24, 342–355. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Xu, B.; Yang, B.; Fu, J.; Chen, H.; Wang, X. Non-Coding RNAs: The Extensive and Interactive Regulators of the Blood-Brain Barrier Permeability. RNA Biol. 2021, 18, 108–116. [Google Scholar] [CrossRef]
- Yang, L.; Han, B.; Zhang, Y.; Bai, Y.; Chao, J.; Hu, G.; Yao, H. Engagement of Circular RNA HECW2 in the Nonautophagic Role of ATG5 Implicated in the Endothelial-Mesenchymal Transition. Autophagy 2018, 14, 404–418. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.; Ni, C.; Jia, C.; Rong, X.; Chu, M.; Wu, R.; Han, B. CircRNA7632 Down-Regulation Alleviates Endothelial Cell Dysfunction in Kawasaki Disease via Regulating IL-33 Expression. Cell Stress Chaperones 2023, 28, 363–374. [Google Scholar] [CrossRef]
- Meng, L.; Zhang, Y.; Wu, P.; Li, D.; Lu, Y.; Shen, P.; Yang, T.; Shi, G.; Chen, Q.; Yuan, H.; et al. CircSTX6 Promotes Pancreatic Ductal Adenocarcinoma Progression by Sponging miR-449b-5p and Interacting with CUL2. Mol. Cancer 2022, 21, 121. [Google Scholar] [CrossRef]
- Rossi, F.; Beltran, M.; Damizia, M.; Grelloni, C.; Colantoni, A.; Setti, A.; Di Timoteo, G.; Dattilo, D.; Centrón-Broco, A.; Nicoletti, C.; et al. Circular RNA ZNF609/CKAP5 mRNA Interaction Regulates Microtubule Dynamics and Tumorigenicity. Mol. Cell 2022, 82, 75–89.e9. [Google Scholar] [CrossRef]
- Zhu, X.; Yang, M.; Zhao, P.; Li, S.; Zhang, L.; Huang, L.; Huang, Y.; Fei, P.; Yang, Y.; Zhang, S.; et al. Catenin α 1 Mutations Cause Familial Exudative Vitreoretinopathy by Overactivating Norrin/β-Catenin Signaling. J. Clin. Investig. 2021, 131, e139869. [Google Scholar] [CrossRef]
- Chan, Y.H.; Harith, H.H.; Israf, D.A.; Tham, C.L. Differential Regulation of LPS-Mediated VE-Cadherin Disruption in Human Endothelial Cells and the Underlying Signaling Pathways: A Mini Review. Front. Cell Dev. Biol. 2019, 7, 280. [Google Scholar] [CrossRef]
- Sudhir, P.-R.; Lin, S.-T.; Chia-Wen, C.; Yang, S.-H.; Li, A.F.-Y.; Lai, R.-H.; Wang, M.-J.; Chen, Y.-T.; Chen, C.-F.; Jou, Y.-S.; et al. Loss of PTPRM Associates with the Pathogenic Development of Colorectal Adenoma-Carcinoma Sequence. Sci. Rep. 2015, 5, 9633. [Google Scholar] [CrossRef]
- Cash, A.; de Jager, C.; Brickler, T.; Soliman, E.; Ladner, L.; Kaloss, A.M.; Zhu, Y.; Pridham, K.J.; Mills, J.; Ju, J.; et al. Endothelial Deletion of EPH Receptor A4 Alters Single-Cell Profile and Tie2/Akap12 Signaling to Preserve Blood-Brain Barrier Integrity. Proc. Natl. Acad. Sci. USA 2023, 120, e2204700120. [Google Scholar] [CrossRef] [PubMed]
- Hatanaka, K.; Lanahan, A.A.; Murakami, M.; Simons, M. Fibroblast Growth Factor Signaling Potentiates VE-Cadherin Stability at Adherens Junctions by Regulating SHP2. PLoS ONE 2012, 7, e37600. [Google Scholar] [CrossRef]
- Huang, B.; Krafft, P.R.; Ma, Q.; Rolland, W.B.; Caner, B.; Lekic, T.; Manaenko, A.; Le, M.; Tang, J.; Zhang, J.H. Fibroblast Growth Factors Preserve Blood-Brain Barrier Integrity through RhoA Inhibition after Intracerebral Hemorrhage in Mice. Neurobiol. Dis. 2012, 46, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Klimaschewski, L.; Claus, P. Fibroblast Growth Factor Signalling in the Diseased Nervous System. Mol. Neurobiol. 2021, 58, 3884–3902. [Google Scholar] [CrossRef]
- Kaya, M.; Ahishali, B. Basic Physiology of the Blood-Brain Barrier in Health and Disease: A Brief Overview. Tissue Barriers 2021, 9, 1840913. [Google Scholar] [CrossRef]
- Wang, J.; Xu, F.; Zhu, X.; Li, X.; Li, Y.; Li, J. Targeting microRNAs to Regulate the Integrity of the Blood-Brain Barrier. Front. Bioeng. Biotechnol. 2021, 9, 673415. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Zhang, Y.; Hua, J.; Yang, X.; Zhang, X.; Duan, M.; Zhu, X.; Huang, W.; Chao, J.; Zhou, R.; et al. Silencing microRNA-143 Protects the Integrity of the Blood-Brain Barrier: Implications for Methamphetamine Abuse. Sci. Rep. 2016, 6, 35642. [Google Scholar] [CrossRef]
- Gasparello, J.; d’Aversa, E.; Breveglieri, G.; Borgatti, M.; Finotti, A.; Gambari, R. In Vitro Induction of Interleukin-8 by SARS-CoV-2 Spike Protein Is Inhibited in Bronchial Epithelial IB3-1 Cells by a miR-93-5p agomiR. Int. Immunopharmacol. 2021, 101, 108201. [Google Scholar] [CrossRef]
- Tisato, V.; Silva, J.A.; Scarpellini, F.; Capucci, R.; Marci, R.; Gallo, I.; Salvatori, F.; D’Aversa, E.; Secchiero, P.; Serino, M.L.; et al. Epigenetic Role of LINE-1 Methylation and Key Genes in Pregnancy Maintenance. Sci. Rep. 2024, 14, 3275. [Google Scholar] [CrossRef]
- Vaccarezza, M.; Papa, V.; Milani, D.; Gonelli, A.; Secchiero, P.; Zauli, G.; Gemmati, D.; Tisato, V. Sex/Gender-Specific Imbalance in CVD: Could Physical Activity Help to Improve Clinical Outcome Targeting CVD Molecular Mechanisms in Women? Int. J. Mol. Sci. 2020, 21, 1477. [Google Scholar] [CrossRef]
- Tornese, G.; Iafusco, D.; Monasta, L.; Agnoletto, C.; Tisato, V.; Ventura, A.; Zauli, G.; Secchiero, P. The Levels of Circulating TRAIL at the Onset of Type 1 Diabetes Are Markedly Decreased in Patients with Ketoacidosis and with the Highest Insulin Requirement. Acta Diabetol. 2014, 51, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Johannes, C.; Moremi, K.E.; Kemp, M.C.; Whati, L.; Engel-Hills, P.; Kidd, M.; van Toorn, R.; Jaftha, M.; van Rensburg, S.J.; Kotze, M.J. Pathology-Supported Genetic Testing Presents Opportunities for Improved Disability Outcomes in Multiple Sclerosis. Pers. Med. 2023, 20, 107–130. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| circRNA ID | Protein Target | Regulation | Ref. |
|---|---|---|---|
| hsa_circ_0074158 | CTNNA1 | Suppressor | [147] |
| hsa_circ_0004188 | PTPRM | Suppressor | |
| hsa_circ_ACTN1_2453 | ACTN1 | Suppressor | |
| hsa_circ_0002872 | PTPRM | Suppressor | |
| hsa_circ_0074171 | CTNNA1 | Suppressor | |
| hsa_circ_0046813 | PTPRM | Suppressor | |
| hsa_circ_0002913 | ACTN1 | Suppressor | |
| hsa_circ_0032321 | ACTN1 | Suppressor | |
| hsa_circ_0008194 | CTNNA1 | Suppressor | |
| hsa_circ_0007440 | CTNNA1 | Suppressor | |
| hsa_circ_0006114 | PTPRM | Enhancer | |
| hsa_circ_0008016 | FGFR1 | Enhancer | |
| hsa_circ_PTPRM_4378 | PTPRM | Enhancer | |
| hsa_circ_ACTN1_2450 | ACTN1 | Enhancer | |
| hsa_circ_0007644 | ACTN1 | Enhancer | |
| hsa_circ_0005564 | FGFR1 | Enhancer | |
| hsa_circ_DLGAP4 | ZO-1, claudin-5, occludin | Enhancer | [150,151] |
| hsa_circ_USP1 | ZO-1, claudin-5, occludin | Enhancer | [152] |
| has_circ_HECW2 | ZO-1, claudin-5, occludin | Suppressor | [153,154] |
| has_circ_7632 | vimentin | Enhancer | [155] |
| hsa_circ_Cul2 | vimentin | Enhancer | [156] |
| hsa_circ_2858 | VEGFA | Enhancer | [143,153] |
| hsa_circ_ZNF609 | CKAP5 | Enhancer | [157] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Aversa, E.; Salvatori, F.; Vaccarezza, M.; Antonica, B.; Grisafi, M.; Singh, A.V.; Secchiero, P.; Zauli, G.; Tisato, V.; Gemmati, D. circRNAs as Epigenetic Regulators of Integrity in Blood–Brain Barrier Architecture: Mechanisms and Therapeutic Strategies in Multiple Sclerosis. Cells 2024, 13, 1316. https://doi.org/10.3390/cells13161316
D’Aversa E, Salvatori F, Vaccarezza M, Antonica B, Grisafi M, Singh AV, Secchiero P, Zauli G, Tisato V, Gemmati D. circRNAs as Epigenetic Regulators of Integrity in Blood–Brain Barrier Architecture: Mechanisms and Therapeutic Strategies in Multiple Sclerosis. Cells. 2024; 13(16):1316. https://doi.org/10.3390/cells13161316
Chicago/Turabian StyleD’Aversa, Elisabetta, Francesca Salvatori, Mauro Vaccarezza, Bianca Antonica, Miriana Grisafi, Ajay Vikram Singh, Paola Secchiero, Giorgio Zauli, Veronica Tisato, and Donato Gemmati. 2024. "circRNAs as Epigenetic Regulators of Integrity in Blood–Brain Barrier Architecture: Mechanisms and Therapeutic Strategies in Multiple Sclerosis" Cells 13, no. 16: 1316. https://doi.org/10.3390/cells13161316