
Nanoparticle Uptake in the Aging and Oncogenic Drosophila Midgut Measured with Surface-Enhanced Raman Spectroscopy


Abstract
:1. Introduction
2. Materials and Methods
- -
- Fly production
- -
- SERS NP synthesis
- -
- Administration of SERS nanoparticles
- -
- Whole-fly Raman measurements
- -
- Midgut preparation for Raman imaging
- -
- Smurf assay for gut leakiness
- -
- Statistical analysis
3. Results
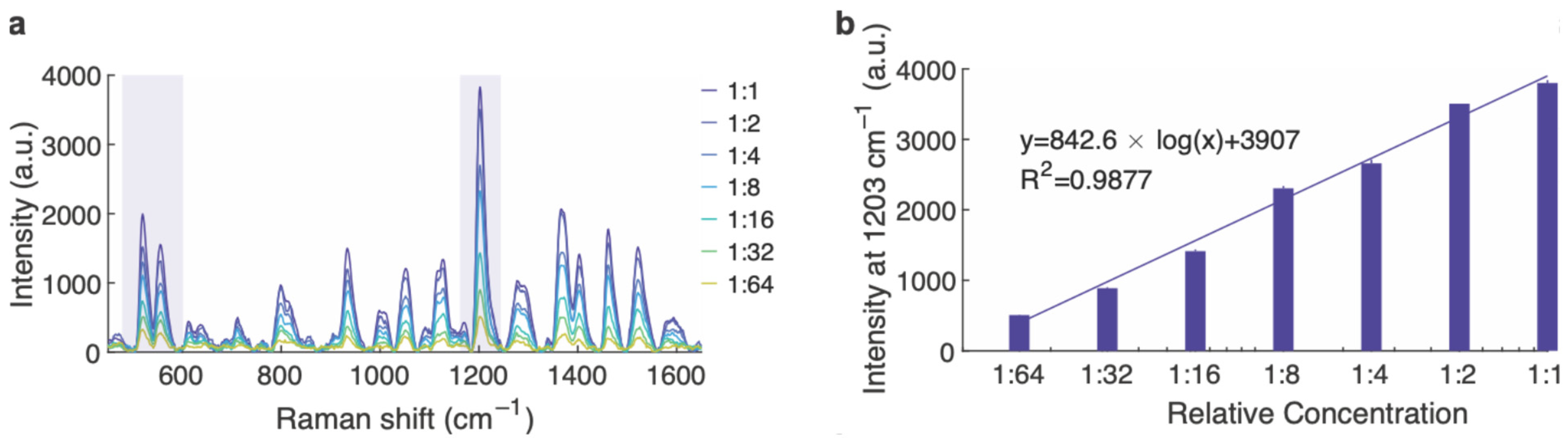
3.1. SERS Nanoparticle Characterization
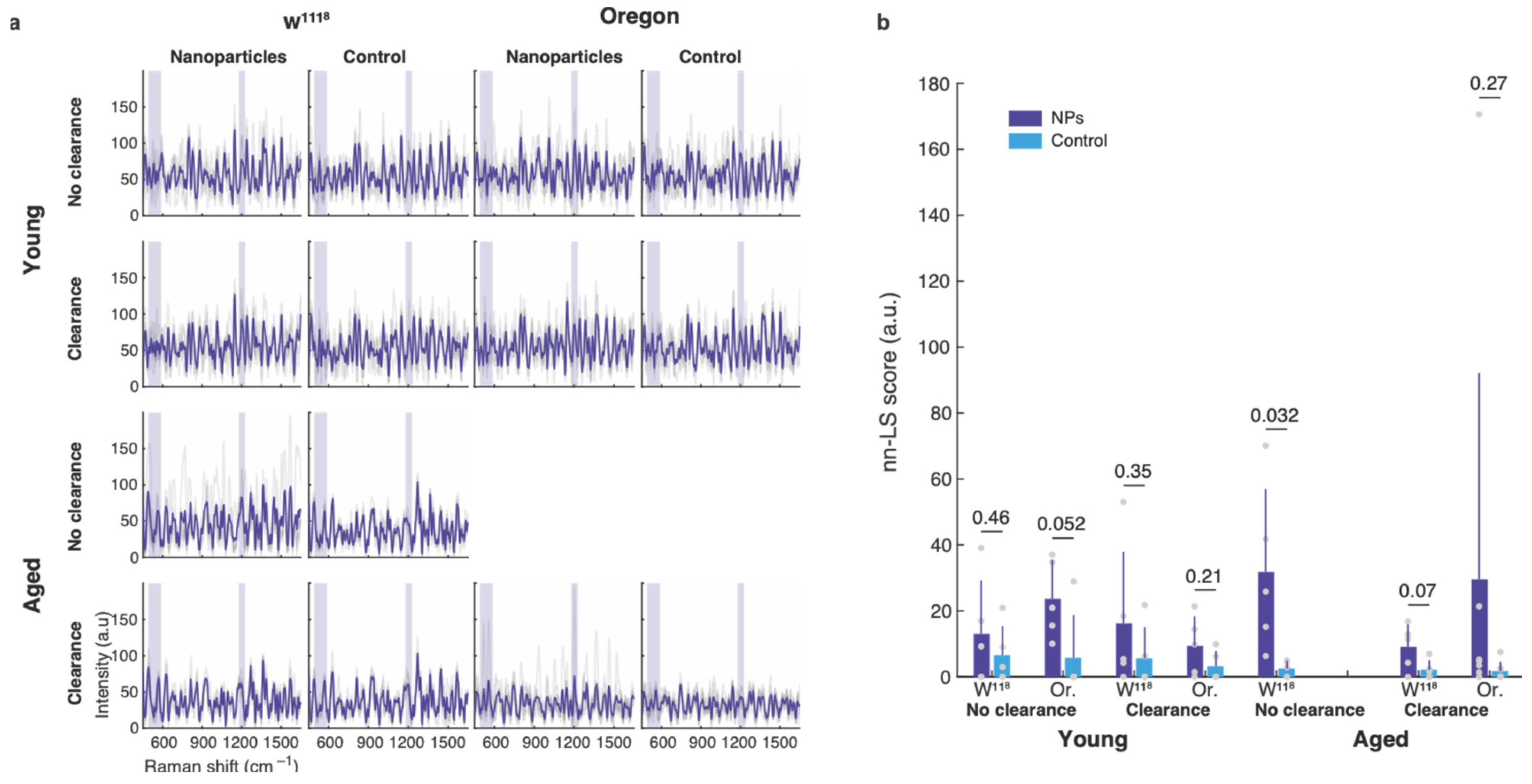
3.2. Nanoparticle Uptake by the Gut of Young and Old Flies
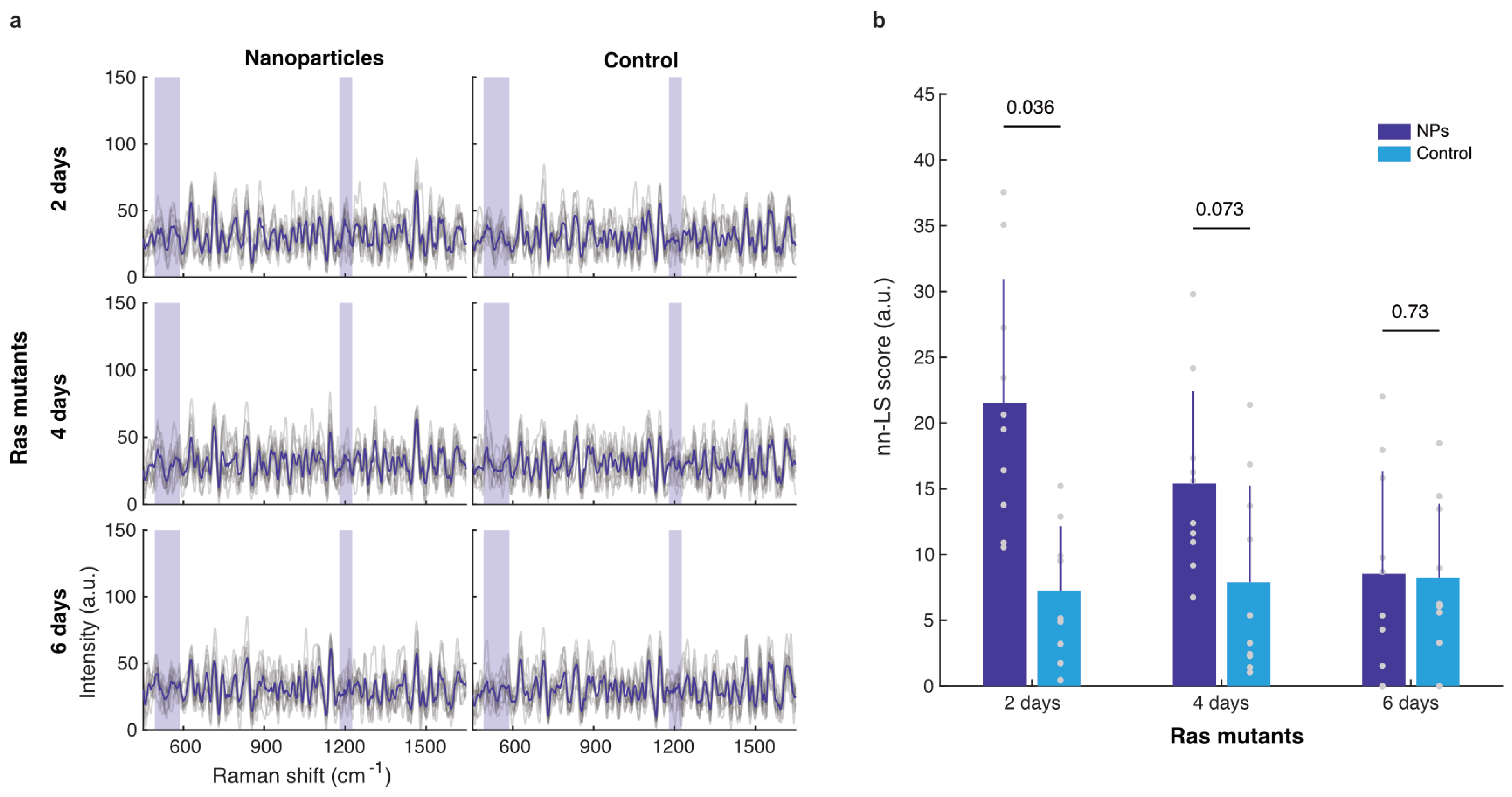
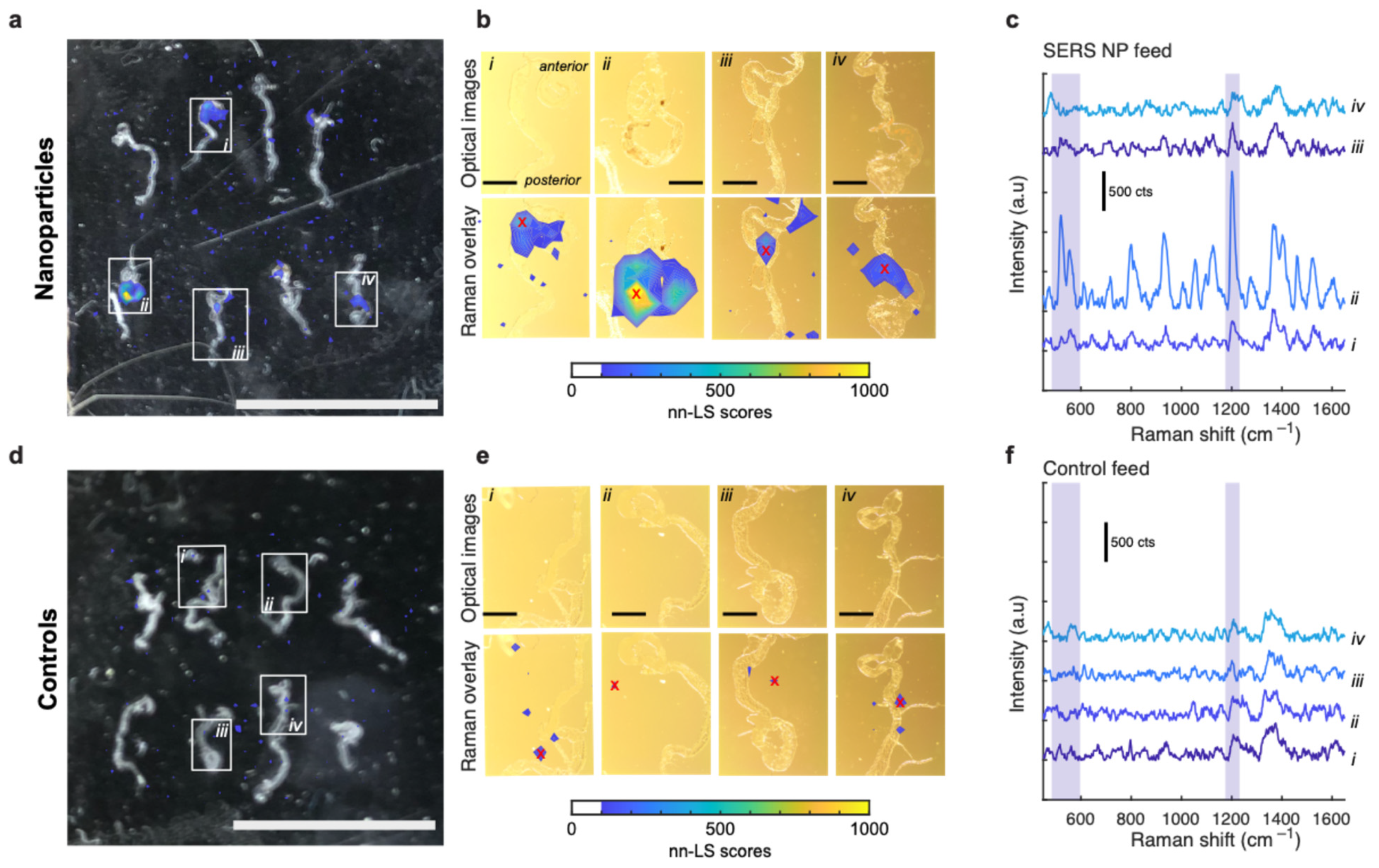
3.3. Nanoparticle Uptake by Oncogenic Flies
3.3.1. Whole-Abdomen Measurements
3.3.2. Excised Gut Measurements
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global Cancer Statistics 2022: Globocan Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef] [PubMed]
- Avram, L.; Stefancu, A.; Crisan, D.; Leopold, N.; Donca, V.; Buzdugan, E.; Craciun, R.; Andras, D.; Coman, I. Recent Advances in Surface-Enhanced Raman Spectroscopy Based Liquid Biopsy for Colorectal Cancer (Review). Exp. Ther. Med. 2020, 20, 213. [Google Scholar] [CrossRef]
- Anderson, R.; Burr, N.E.; Valori, R. Causes of Post-Colonoscopy Colorectal Cancers Based on World Endoscopy Organization System of Analysis. Gastroenterology 2020, 158, 1287–1299.e2. [Google Scholar] [CrossRef] [PubMed]
- Langer, J.; Jimenez de Aberasturi, D.; Aizpurua, J.; Alvarez-Puebla, R.A.; Auguie, B.; Baumberg, J.J.; Bazan, G.C.; Bell, S.E.J.; Boisen, A.; Brolo, A.G.; et al. Present and Future of Surface-Enhanced Raman Scattering. ACS Nano 2020, 14, 28–117. [Google Scholar] [CrossRef] [PubMed]
- Kenry; Nicolson, F.; Clark, L.; Panikkanvalappil, S.R.; Andreiuk, B.; Andreou, C. Advances in Surface Enhanced Raman Spectroscopy for In Vivo Imaging in Oncology. Nanotheranostics 2022, 6, 31–49. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.W.; Khan, A.; Leigh, S.Y.; Wang, D.; Chen, Y.; Meza, D.; Liu, J.T. Comprehensive Spectral Endoscopy of Topically Applied Sers Nanoparticles in the Rat Esophagus. Biomed. Opt. Express 2014, 5, 2883–2895. [Google Scholar] [CrossRef] [PubMed]
- Garai, E.; Sensarn, S.; Zavaleta, C.L.; Van de Sompel, D.; Loewke, N.O.; Mandella, M.J.; Gambhir, S.S.; Contag, C.H. High-Sensitivity, Real-Time, Ratiometric Imaging of Surface-Enhanced Raman Scattering Nanoparticles with a Clinically Translatable Raman Endoscope Device. J. Biomed. Opt. 2013, 18, 096008. [Google Scholar] [CrossRef] [PubMed]
- Garai, E.; Sensarn, S.; Zavaleta, C.L.; Loewke, N.O.; Rogalla, S.; Mandella, M.J.; Felt, S.A.; Friedland, S.; Liu, J.T.; Gambhir, S.S.; et al. A Real-Time Clinical Endoscopic System for Intraluminal, Multiplexed Imaging of Surface-Enhanced Raman Scattering Nanoparticles. PLoS ONE 2015, 10, e0123185. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.; Wang, Y.; Ji, Y.; Mei, R.; Zhao, X.; Song, J.; You, J.; Chen, L.; Wang, X. Inflammatory Bowel Disease Alters In Vivo Distribution of Orally Administrated Nanoparticles: Revealing via Sers Tag Labeling Technique. Talanta 2024, 275, 126172. [Google Scholar] [CrossRef]
- Moisoiu, V.; Stefancu, A.; Gulei, D.; Boitor, R.; Magdo, L.; Raduly, L.; Pasca, S.; Kubelac, P.; Mehterov, N.; Chis, V.; et al. SERS-Based Differential Diagnosis between Multiple Solid Malignancies: Breast, Colorectal, Lung, Ovarian and Oral Cancer. Int. J. Nanomed. 2019, 14, 6165–6178. [Google Scholar] [CrossRef]
- Constantinou, M.; Hadjigeorgiou, K.; Abalde-Cela, S.; Andreou, C. Label-Free Sensing with Metal Nanostructure-Based Surface-Enhanced Raman Spectroscopy for Cancer Diagnosis. ACS Appl. Nano Mater. 2022, 5, 12276–12299. [Google Scholar] [CrossRef] [PubMed]
- Apidianakis, Y.; Rahme, L.G. Drosophila melanogaster as a Model for Human Intestinal Infection and Pathology. Dis. Model. Mech. 2011, 4, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Ugur, B.; Chen, K.C.; Bellen, H.J. Tools and Assays for the Study of Human Diseases. Dis. Models Mech. 2016, 9, 235–244. [Google Scholar] [CrossRef]
- Millburn, G.H.; Crosby, M.A.; Gramates, L.S.; Tweedie, S.; Consortium, F. FlyBase Portals to Human Disease Research Using Drosophila Models. Dis. Models Mech. 2016, 9, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Bangi, E.; Murgia, C.; Teague, A.G.S.; Sansom, O.J.; Cagan, R.L. Functional Exploration of Colorectal Cancer Genomes Using. Nat. Commun. 2016, 7, 13615. [Google Scholar] [CrossRef] [PubMed]
- Broderick, N.A.; Lemaitre, B. Gut-Associated Microbes of Drosophila melanogaster. Gut Microbes 2012, 3, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Capo, F.; Wilson, A.; Di Cara, F. The Intestine of Drosophila melanogaster: An Emerging Versatile Model System to Study Intestinal Epithelial Homeostasis and Host-Microbial Interactions in Humans. Microorganisms 2019, 7, 336. [Google Scholar] [CrossRef] [PubMed]
- Ji, D.; Sun, H.; Yang, W.; Gao, M.; Xu, H. Transfer of Human Microbiome to Drosophila Gut Model. Microorganisms 2022, 10, 553. [Google Scholar] [CrossRef] [PubMed]
- Ludington, W.B.; Ja, W.W. Drosophila melanogaster as a Model for the Gut Microbiome. PLoS Pathog. 2020, 16, e1008398. [Google Scholar] [CrossRef]
- Apidianakis, Y.; Pitsouli, C.; Perrimon, N.; Rahme, L. Synergy between Bacterial Infection and Genetic Predisposition in Intestinal Dysplasia. Proc. Natl. Acad. Sci. USA 2009, 106, 20883–20888. [Google Scholar] [CrossRef]
- Andreou, C.; Oseledchyk, A.; Nicolson, F.; Berisha, N.; Pal, S.; Kircher, M.F. Surface-enhanced Resonance Raman Scattering Nanoprobe Ratiometry for Detecting Microscopic Ovarian Cancer via Folate Receptor Targeting. J. Vis. Exp. 2019, 145, e58389. [Google Scholar]
- Martins, R.R.; McCracken, A.W.; Simons, M.J.P.; Henriques, C.M.; Rera, M. How to Catch a Smurf?—Ageing and Beyond… In Vivo Assessment of Intestinal Permeability in Multiple Model Organisms. Bio Protoc. 2018, 8, e2722. [Google Scholar] [CrossRef] [PubMed]
- Rera, M.; Bahadorani, S.; Cho, J.; Koehler, C.L.; Ulgherait, M.; Hur, J.H.; Ansari, W.S.; Lo, T., Jr.; Jones, D.L.; Walker, D.W. Modulation of Longevity and Tissue Homeostasis by the Drosophila Pgc-1 Homolog. Cell Metab. 2011, 14, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Regan, J.C.; Khericha, M.; Dobson, A.J.; Bolukbasi, E.; Rattanavirotkul, N.; Partridge, L. Sex Difference in Pathology of the Ageing Gut Mediates the Greater Response of Female Lifespan to Dietary Restriction. eLife 2016, 5, e10956. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Bagheri, P.; Chang, P.; Zeng, A.; Hao, J.; Fung, A.; Wu, J.Y.; Shi, L. Direct Imaging of Lipid Metabolic Changes in Drosophila Ovary During Aging Using Do-Srs Microscopy. Front. Aging 2021, 2, 819903. [Google Scholar] [CrossRef] [PubMed]
- Shianiou, G.; Teloni, S.; Apidianakis, Y. Intestinal Immune Deficiency and Juvenile Hormone Signaling Mediate a Metabolic Trade-Off in Adult Drosophila Females. Metabolites 2023, 13, 340. [Google Scholar] [CrossRef]
- Siudeja, K.; Nassari, S.; Gervais, L.; Skorski, P.; Lameiras, S.; Stolfa, D.; Zande, M.; Bernard, V.; Frio, T.R.; Bardin, A.J. Frequent Somatic Mutation in Adult Intestinal Stem Cells Drives Neoplasia and Genetic Mosaicism during Aging. Cell Stem Cell 2015, 17, 663–674. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Saavedra, P.; Perrimon, N. Cancer cachexia: Lessons from Drosophila. Dis. Model. Mech. 2022, 15, dmm049298. [Google Scholar] [CrossRef]
- Martin, T.A.; Jiang, W.G. Loss of Tight Junction Barrier Function and Its Role in Cancer Metastasis. Biochim. Biophys. Acta 2009, 1788, 872–891. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | OR | w1118 | ||||||
|---|---|---|---|---|---|---|---|---|
| Condition | Young | Aged | Young | Aged | ||||
| n | % | n | % | n | % | n | % | |
| Total | 63 | 100.0 | 17 | 100.0 | 42 | 100.0 | 24 | 100.0 |
| Dead | 2 | 3.2 | 1 | 5.9 | 2 | 4.8 | 3 | 12.5 |
| Non-Smurf | 58 | 92.1 | 9 | 52.9 | 34 | 81.0 | 11 | 45.8 |
| Smurf | 3 | 4.8 | 7 | 41.2 | 6 | 14.3 | 10 | 41.7 |
| p-value | 0.0004 | 0.0124 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Christou, M.; Fidelix, A.; Apidianakis, Y.; Andreou, C. Nanoparticle Uptake in the Aging and Oncogenic Drosophila Midgut Measured with Surface-Enhanced Raman Spectroscopy. Cells 2024, 13, 1344. https://doi.org/10.3390/cells13161344
Christou M, Fidelix A, Apidianakis Y, Andreou C. Nanoparticle Uptake in the Aging and Oncogenic Drosophila Midgut Measured with Surface-Enhanced Raman Spectroscopy. Cells. 2024; 13(16):1344. https://doi.org/10.3390/cells13161344
Chicago/Turabian StyleChristou, Maria, Ayobami Fidelix, Yiorgos Apidianakis, and Chrysafis Andreou. 2024. "Nanoparticle Uptake in the Aging and Oncogenic Drosophila Midgut Measured with Surface-Enhanced Raman Spectroscopy" Cells 13, no. 16: 1344. https://doi.org/10.3390/cells13161344