
Roles of M1 Macrophages and Their Extracellular Vesicles in Cancer Therapy

Abstract
:
1. Introduction
2. M1 Macrophage-Derived Extracellular Vesicles
2.1. Tumor Cells and the Tumor Microenvironment
2.2. Cells Secrete Extracellular Vesicles to Transmit Biological Signals and Regulate the Surrounding Microenvironment
2.3. EVs Signal by Transferring microRNAs
2.4. M1-EVs Show Promise in Cancer Models through microRNA Transfer
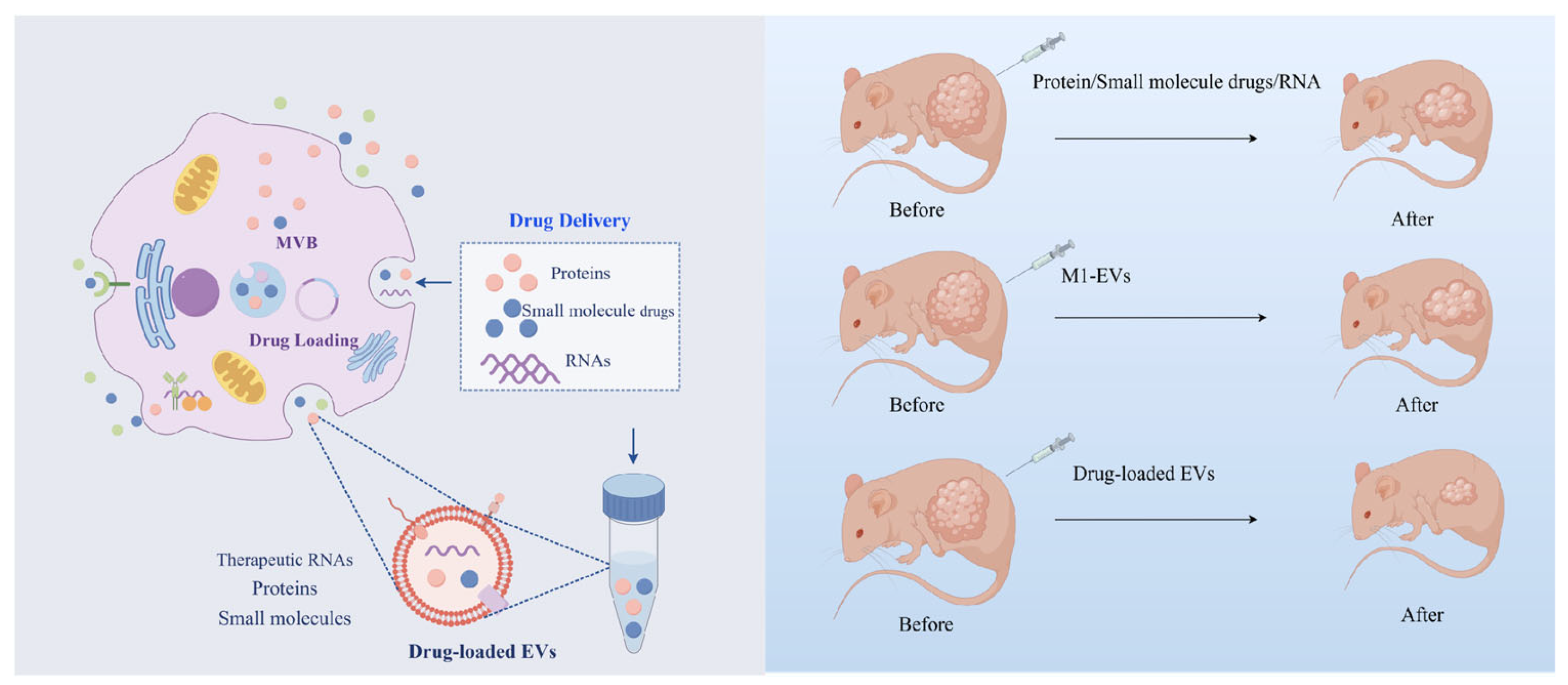
3. M1-EVs Loaded with Drugs Act Synergistically to Fight Cancer
3.1. Advantages of M1-EVs as Drug Carriers
3.2. M1-EVs Loaded with Drugs to Achieve Better Therapeutic Effects
3.3. Future Perspectives of M1-EVs as Drug Carriers
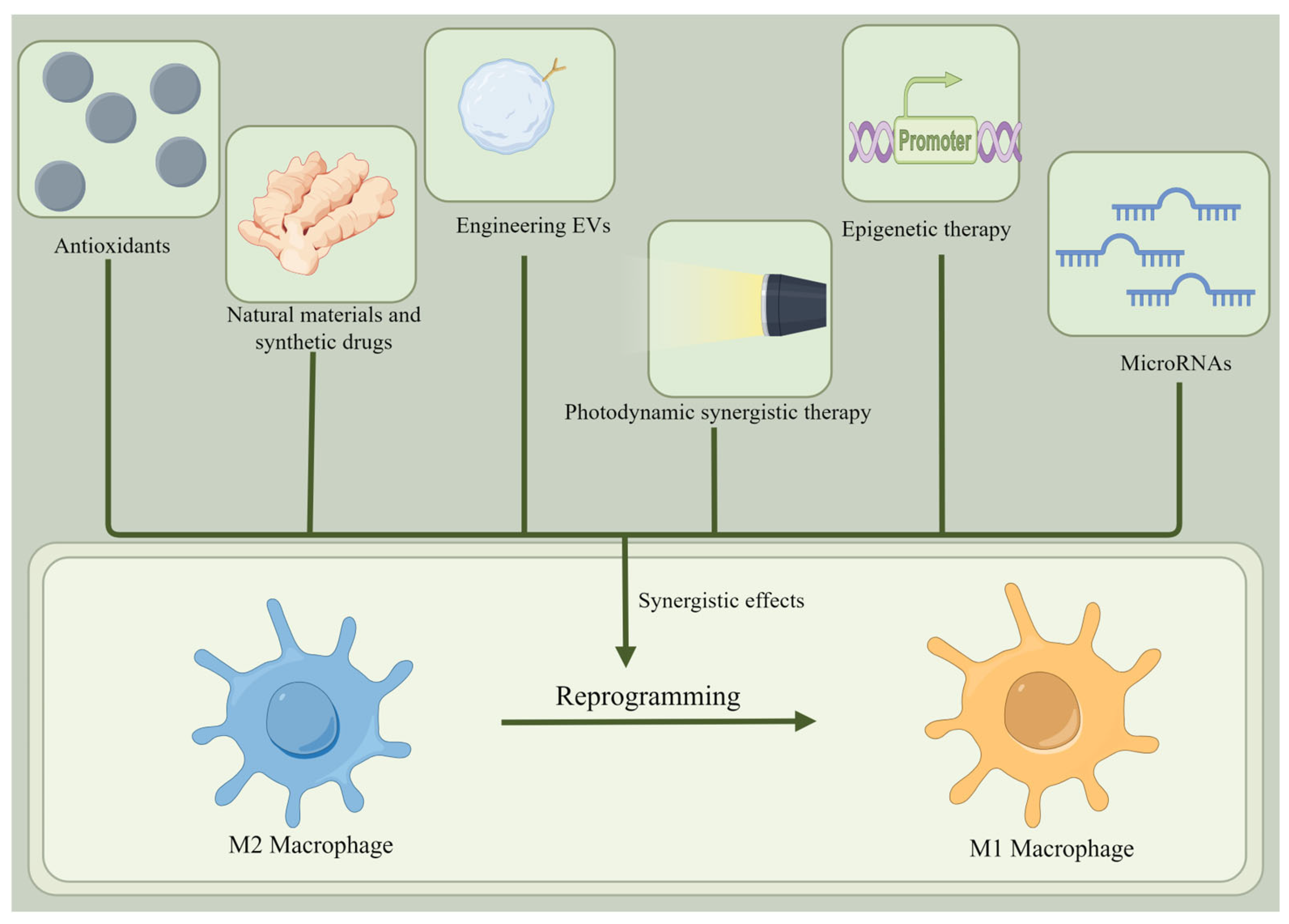
4. Reprogramming of M2 Macrophages into M1 Macrophages to Fight Cancer
4.1. Reprogramming M2 to M1 Macrophages: A New Strategy for Cancer Treatment
4.2. Antioxidants
4.3. Photodynamic Synergistic Therapy
4.4. Epigenetic Therapy
4.5. Natural Materials and Synthetic Drugs
4.6. Engineering EVs
4.7. MicroRNAs
4.8. Synergistic Effects
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, F.; Pang, L.; Dunterman, M.; Lesniak, M.S.; Heimberger, A.B.; Chen, P.W. Macrophages and microglia in glioblastoma: Heterogeneity, plasticity, and therapy. J. Clin. Investig. 2023, 133, e163446. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.M.; Wang, J.Z. Prognostic prediction and multidimensional dissections of a macrophages M0-related gene signature in liver cancer. Front. Endocrinol. 2023, 14, 1153562. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.N.; Wang, J.G.; Lu, D.; Xu, X. Targeting tumor-associated macrophages to synergize tumor immunotherapy. Signal Transduct. Tar. 2021, 6, 75. [Google Scholar] [CrossRef]
- Li, M.Y.; Yang, Y.H.; Xiong, L.T.; Jiang, P.; Wang, J.J.; Li, C.X. Metabolism, metabolites, and macrophages in cancer. J. Hematol. Oncol. 2023, 16, 80. [Google Scholar] [CrossRef]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef]
- Gao, J.; Liang, Y.Z.; Wang, L. Shaping Polarization Of Tumor-Associated Macrophages In Cancer Immunotherapy. Front. Immunol. 2022, 13, 888713. [Google Scholar] [CrossRef]
- Kesapragada, M.; Sun, Y.H.; Zlobina, K.; Recendez, C.; Fregoso, D.; Yang, H.Y.; Aslankoohi, E.; Isseroff, R.; Rolandi, M.; Zhao, M.; et al. Deep learning classification for macrophage subtypes through cell migratory pattern analysis. Front. Cell Dev. Biol. 2024, 12, 1259037. [Google Scholar] [CrossRef] [PubMed]
- He, P.C.; Dai, M.X.; Li, Z.P.; Wang, X.Y.; Liu, H.Y.; He, Y.X.; Jiang, H. Effect of connexin 43 in LPS/IL-4-induced macrophage M1/M2 polarization: An observational study. Medicine 2024, 103, e37811. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Shi, F.; Zhou, Z.; Sun, F.; Sun, M.H.; Sun, Q.; Chen, L.; Li, D.; Jiang, C.Y.; Zhao, R.Z.; et al. M1 macrophage mediated increased reactive oxygen species (ROS) influence wound healing via the MAPK signaling in vitro and in vivo. Toxicol. Appl. Pharm. 2019, 366, 83–95. [Google Scholar] [CrossRef]
- Li, L.; Lv, G.H.; Wang, B.; Kuang, L. XIST/miR-376c-5p/OPN axis modulates the influence of proinflammatory M1 macrophages on osteoarthritis chondrocyte apoptosis. J. Cell. Physiol. 2020, 235, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Orsolic, N.; Kunstic, M.; Kukolj, M.; Gracan, R.; Nemrava, J. Oxidative stress, polarization of macrophages and tumour angiogenesis: Efficacy of caffeic acid. Chem. Biol. Interact. 2016, 256, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.J.; Ma, L.L.; Deng, D.L.; Zhang, T.H.; Han, L.L.; Xu, F.; Huang, S.Q.; Ding, Y.Y.; Chen, X.D. M2 macrophage polarization: A potential target in pain relief. Front. Immunol. 2023, 14, 1243149. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Zhang, J.; Li, D.; Mao, Y.; Mo, F.; Du, W.; Ma, X.L. Prognostic significance of tumor-associated macrophages in ovarian cancer: A meta-analysis. Gynecol. Oncol. 2017, 147, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Jackute, J.; Zemaitis, M.; Pranys, D.; Sitkauskiene, B.; Miliauskas, S.; Vaitkiene, S.; Sakalauskas, R. Distribution of M1 and M2 macrophages in tumor islets and stroma in relation to prognosis of non-small cell lung cancer. BMC Immunol. 2018, 19, 3. [Google Scholar] [CrossRef] [PubMed]
- Ruytinx, P.; Proost, P.; Van Damme, J.; Struyf, E. Chemokine-Induced Macrophage Polarization in Inflammatory Conditions. Front. Immunol. 2018, 9, 1930. [Google Scholar] [CrossRef]
- Wen, Y.K.; Lambrecht, J.; Ju, C.; Tacke, F. Hepatic macrophages in liver homeostasis and diseases-diversity, plasticity and therapeutic opportunities. Cell Mol. Immunol. 2021, 18, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Gharavi, A.T.; Hanjani, N.A.; Movahed, E.; Doroudian, M. The role of macrophage subtypes and exosomes in immunomodulation. Cell. Mol. Biol. Lett. 2022, 27, 83. [Google Scholar] [CrossRef]
- Liao, Z.X.; Ou, D.L.; Hsieh, M.J.; Hsieh, C.C. Synergistic Effect of Repolarization of M2 to M1 Macrophages Induced by Iron Oxide Nanoparticles Combined with Lactate Oxidase. Int. J. Mol. Sci. 2021, 22, 13346. [Google Scholar] [CrossRef]
- Atella, M.O.; Carvalho, A.S.; Da Poian, A.T. Role of macrophages in the onset, maintenance, or control of arthritis caused by alphaviruses. Exp. Biol. Med. 2023, 248, 2039–2044. [Google Scholar] [CrossRef]
- Park, H.; Patil, T.V.; Dutta, S.D.; Lee, J.; Ganguly, K.; Randhawa, A.; Kim, H.; Lim, K.T. Extracellular Matrix-Bioinspired Anisotropic Topographical Cues of Electrospun Nanofibers: A Strategy of Wound Healing through Macrophage Polarization. Adv. Healthc. Mater. 2024, 13, 2304114. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.F.; Guo, J.Q.; Yang, Y.L.; Liu, J. Macrophage Polarization Induced by Bacteria-Responsive Antibiotic-Loaded Nanozymes for Multidrug Resistance-Bacterial Infections Management. Small 2023, 19, 2204928. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Laversanne, M.; Sung, H.Y.A.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef]
- Jiao, Y.Q.; Yu, Y.J.; Zheng, M.Y.; Yan, M.; Wang, J.P.; Zhang, Y.; Zhang, S.W. Dormant cancer cells and polyploid giant cancer cells: The roots of cancer recurrence and metastasis. Clin. Transl. Med. 2024, 14, e1567. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Lu, Z.R. Molecular imaging of the tumor microenvironment. Adv. Drug Deliv. Rev. 2017, 113, 24–48. [Google Scholar] [CrossRef] [PubMed]
- Murata, M. Inflammation and cancer. Environ. Health Prev. Med. 2018, 23, 50. [Google Scholar] [CrossRef]
- Anderson, N.M.; Simon, M.C. The tumor microenvironment. Curr. Biol. 2020, 30, R921–R925. [Google Scholar] [CrossRef]
- Fidler, I.J. The pathogenesis of cancer metastasis: The ‘seed and soil’ hypothesis revisited. Nat. Rev. Cancer 2003, 3, 453–458. [Google Scholar] [CrossRef]
- Wu, T.; Dai, Y. Tumor microenvironment and therapeutic response. Cancer Lett. 2017, 387, 61–68. [Google Scholar] [CrossRef]
- Gurunathan, S.; Kang, M.H.; Qasim, M.; Khan, K.; Kim, J.H. Biogenesis, Membrane Trafficking, Functions, and Next Generation Nanotherapeutics Medicine of Extracellular Vesicles. Int. J. Nanomed. 2021, 16, 3357–3383. [Google Scholar] [CrossRef]
- Kalra, H.; Drummen, G.P.C.; Mathivanan, S. Focus on Extracellular Vesicles: Introducing the Next Small Big Thing. Int. J. Mol. Sci. 2016, 17, 170. [Google Scholar] [CrossRef]
- Pan, B.T.; Johnstone, R.M. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: Selective externalization of the receptor. Cell 1983, 33, 967–978. [Google Scholar] [CrossRef]
- Johnstone, R.M.; Adam, M.; Hammond, J.R.; Orr, L.; Turbide, C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J. Biol. Chem. 1987, 262, 9412–9420. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.T.; Tian, T.T.; Yu, H.; Cao, C.Y.; Zhang, Z.Y.; He, Z.Z.; Ma, Z.Q.; Cai, R.; Li, F.N.; Pang, W.J. Identification of porcine fast/slow myogenic exosomes and their regulatory effects on lipid accumulation in intramuscular adipocytes. J. Anim. Sci. Biotechnol. 2024, 15, 73. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.M.; Liu, T.Q.; Zhou, M.J. Immune-Cell-Derived Exosomes for Cancer Therapy. Mol. Pharmaceut. 2022, 19, 3042–3056. [Google Scholar] [CrossRef]
- Thery, C. Exosomes: Secreted vesicles and intercellular communications. F1000 Biol. Rep. 2011, 3, 15. [Google Scholar] [CrossRef]
- Yuan, D.F.; Zhao, Y.L.; Banks, W.A.; Bullock, K.M.; Haney, M.; Batrakova, E.; Kabanov, A.V. Macrophage exosomes as natural nanocarriers for protein delivery to inflamed brain. Biomaterials 2017, 142, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Naranjo, J.C.; Wu, H.J.; Ugaz, V.M. Microfluidics for exosome isolation and analysis: Enabling liquid biopsy for personalized medicine. Lab Chip 2017, 17, 3558–3577. [Google Scholar] [CrossRef]
- Kalluri, R. The biology and function of exosomes in cancer. J. Clin. Investig. 2016, 126, 1208–1215. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Lu, M.; Xing, H.N.; Xun, Z.; Yang, T.Z.; Ding, P.T.; Cai, C.F.; Wang, D.K.; Zhao, X.Y. Exosome-based small RNA delivery: Progress and prospects. Asian J. Pharm. Sci. 2018, 13, 1–11. [Google Scholar] [CrossRef]
- Ni, P.S.; Yang, L.D.; Li, F.H. Exercise-derived skeletal myogenic exosomes as mediators of intercellular crosstalk: A major player in health, disease, and exercise. J. Physiol. Biochem. 2023, 79, 501–510. [Google Scholar] [CrossRef]
- Hosseini, R.; Asef-Kabiri, L.; Yousefi, H.; Sarvnaz, H.; Salehi, M.; Akbari, M.E.; Eskandari, N. The roles of tumor-derived exosomes in altered differentiation, maturation and function of dendritic cells. Mol. Cancer 2021, 20, 83. [Google Scholar] [CrossRef] [PubMed]
- Li, Q. Role of exosomes in cellular communication between tumor cells and the tumor microenvironment. Oncol. Lett. 2022, 24, 240. [Google Scholar] [CrossRef] [PubMed]
- Shan, X.; Zhang, C.; Mai, C.; Hu, X.; Cheng, N.; Chen, W.; Peng, D.; Wang, L.; Ji, Z.; Xie, Y. The Biogenesis, Biological Functions, and Applications of Macrophage-Derived Exosomes. Front. Mol. Biosci. 2021, 8, 715461. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Wang, Y.; Huang, L. Exosomes from M1-Polarized Macrophages Potentiate the Cancer Vaccine by Creating a Pro-inflammatory Microenvironment in the Lymph Node. Mol. Ther. 2017, 25, 1665–1675. [Google Scholar] [CrossRef]
- Rashid, H.; Hossain, B.; Siddiqua, T.; Kabir, M.; Noor, Z.; Ahmed, M.; Haque, R. Fecal MicroRNAs as Potential Biomarkers for Screening and Diagnosis of Intestinal Diseases. Front. Mol. Biosci. 2020, 7, 181. [Google Scholar] [CrossRef]
- Zaichenko, D.M.; Mikryukova, A.A.; Astafeva, I.R.; Malakho, S.G.; Kubatiev, A.A.; Moskovtsev, A.A. MicroRNA Biogenesis in Cell Senescence Induced by Chronic Endoplasmic Reticulum Stress. Mol. Biol. 2023, 57, 670–683. [Google Scholar] [CrossRef]
- Hynes, C.; Kakumani, P.K. Regulatory role of RNA-binding proteins in microRNA biogenesis. Front. Mol. Biosci. 2024, 11, 1374843. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Jeppesen, D.K.; Fenix, A.M.; Franklin, J.L.; Higginbotham, J.N.; Zhang, Q.; Zimmerman, L.J.; Liebler, D.C.; Ping, J.; Liu, Q.; Evans, R.; et al. Reassessment of Exosome Composition. Cell 2019, 177, 428–445. [Google Scholar] [CrossRef]
- Chiou, N.T.; Kageyama, R.; Ansel, K.M. Selective Export into Extracellular Vesicles and Function of tRNA Fragments during T Cell Activation. Cell Rep. 2018, 25, 3356–3370. [Google Scholar] [CrossRef] [PubMed]
- Bhome, R.; Del Vecchio, F.; Lee, G.H.; Bullock, M.D.; Primrose, J.N.; Sayan, A.E.; Mirnezami, A.H. Exosomal microRNAs (exomiRs): Small molecules with a big role in cancer. Cancer Lett. 2018, 420, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.N.; Lei, Z.; He, J.; Zhang, L.; Lai, T.M.; Zhou, L.; Wang, N.H.; Tang, Z.; Sui, J.D.; Wu, Y.Z. Single-cell RNA sequencing reveals recruitment of the M2-like CCL8 macrophages in Lewis lung carcinoma-bearing mice following hypofractionated radiotherapy. J. Transl. Med. 2024, 22, 306. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.T.; Suo, B.; Long, G.; Gao, Y.; Song, J.; Zhang, M.Z.; Feng, B.Y.; Shang, C.; Wang, D.W. Exosomal miRNA-16-5p Derived From M1 Macrophages Enhances T Cell-Dependent Immune Response by Regulating PD-L1 in Gastric Cancer. Front. Cell Dev. Biol. 2020, 8, 572689. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.C.; Li, S.; Li, B.; Hu, W.X.; Ding, C.M. Exosomes derived from M1 macrophages inhibit the proliferation of the A549 and H1299 lung cancer cell lines via the miRNA-let-7b-5p-GNG5 axis. PeerJ 2023, 11, e14608. [Google Scholar] [CrossRef]
- Okusha, Y.; Eguchi, T.; Sogawa, C.; Okui, T.; Nakano, K.; Okamoto, K.; Kozaki, K.I. The intranuclear PEX domain of MMP involves proliferation, migration, and metastasis of aggressive adenocarcinoma cells. J. Cell Biochem. 2018, 119, 7363–7376. [Google Scholar] [CrossRef]
- Yan, P.; Wang, J.; Liu, H.; Liu, X.; Fu, R.; Feng, J. M1 macrophage-derived exosomes containing miR-150 inhibit glioma progression by targeting MMP16. Cell Signal 2023, 108, 110731. [Google Scholar] [CrossRef]
- Wang, J.J.; Ji, X.M.; Liu, J.J.; Zhang, X. Serine/Threonine Protein Kinase STK16. Int. J. Mol. Sci. 2019, 20, 1760. [Google Scholar] [CrossRef]
- Wang, X.; Huang, R.; Lu, Z.; Wang, Z.; Chen, X.; Huang, D. Exosomes from M1-polarized macrophages promote apoptosis in lung adenocarcinoma via the miR-181a-5p/ETS1/STK16 axis. Cancer Sci. 2022, 113, 986–1001. [Google Scholar] [CrossRef]
- Wang, L.; Yi, X.; Xiao, X.; Zheng, Q.; Ma, L.; Li, B. Exosomal miR-628-5p from M1 polarized macrophages hinders m6A modification of circFUT8 to suppress hepatocellular carcinoma progression. Cell Mol. Biol. Lett. 2022, 27, 106. [Google Scholar] [CrossRef]
- Milman, N.; Ginini, L.; Gil, Z. Exosomes and their role in tumorigenesis and anticancer drug resistance. Drug Resist. Update 2019, 45, 1–12. [Google Scholar] [CrossRef]
- Mondal, J.; Pillarisetti, S.; Junnuthula, V.; Saha, M.; Hwang, S.R.; Park, I.K.; Lee, Y.K. Hybrid exosomes, exosome-like nanovesicles and engineered exosomes for therapeutic applications. J. Control. Release 2023, 353, 1127–1149. [Google Scholar] [CrossRef] [PubMed]
- Dubey, A.; Lobo, C.L.; Ravi, G.S.; Shetty, A.; Hebbar, S.; El-Zahaby, S.A. Exosomes: Emerging implementation of nanotechnology for detecting and managing novel corona virus-SARS-CoV-2. Asian J. Pharm. Sci. 2022, 17, 20–34. [Google Scholar] [CrossRef]
- Liu, F.J.; Meng, F.S.; Yang, Z.J.; Wang, H.; Ren, Y.H.; Cai, Y.; Zhang, X.W. Exosome-biomimetic nanocarriers for oral drug delivery. Chinese Chem. Lett. 2024, 35, 109335. [Google Scholar] [CrossRef]
- Kim, H.; Kim, E.H.; Kwak, G.; Chi, S.G.; Kim, S.H.; Yang, Y. Exosomes: Cell-Derived Nanoplatforms for the Delivery of Cancer Therapeutics. Int. J. Mol. Sci. 2021, 22, 14. [Google Scholar] [CrossRef]
- Gao, P.Y.; Li, X.R.; Du, X.Z.; Liu, S.; Xu, Y. Diagnostic and Therapeutic Potential of Exosomes in Neurodegenerative Diseases. Front. Aging Neurosci. 2021, 13, 790863. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Zhuang, X.; Xiang, X.; Liu, Y.; Zhang, S.; Liu, C.; Barnes, S.; Grizzle, W.; Miller, D.; Zhang, H.-G. A novel nanoparticle drug delivery system: The anti-inflammatory activity of curcumin is enhanced when encapsulated in exosomes. Mol. Ther. 2010, 18, 1606–1614. [Google Scholar] [CrossRef] [PubMed]
- Shahabipour, F.; Barati, N.; Johnston, T.P.; Derosa, G.; Maffioli, P.; Sahebkar, A. Exosomes: Nanoparticulate tools for RNA interference and drug delivery. J. Cell. Physiol. 2017, 232, 1660–1668. [Google Scholar] [CrossRef]
- Zhao, Y.M.; Zheng, Y.L.; Zhu, Y.; Li, H.Y.; Zhu, H.Y.; Liu, T.Q. Docetaxel-loaded M1 macrophage-derived exosomes for a safe and efficient chemoimmunotherapy of breast cancer. J. Nanobiotechnol. 2022, 20, 359. [Google Scholar] [CrossRef]
- Zhang, X.H.; Wang, J.P.; Liu, N.; Wu, W.M.; Li, H.; Lu, W.; Guo, X.Q. Umbilical Cord Blood-Derived M1 Macrophage Exosomes Loaded with Cisplatin Target Ovarian Cancer and Reverse Cisplatin Resistance. Mol. Pharmaceut. 2023, 20, 5440–5453. [Google Scholar] [CrossRef]
- Zhang, H.; Cao, K.L.; Xiang, J.R.; Zhang, M.T.; Zhu, M.X.; Xi, Q.H. Hypoxia induces immunosuppression, metastasis and drug resistance in pancreatic cancers. Cancer Lett. 2023, 571, 216345. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.P. Pancreatic Cancer: Current Progress and Future Challenges. Int. J. Biol. Sci. 2016, 12, 270–272. [Google Scholar] [CrossRef] [PubMed]
- Samanta, K.; Setua, S.; Kumari, S.; Jaggi, M.; Yallapu, M.M.; Chauhan, S.C. Gemcitabine Combination Nano Therapies for Pancreatic Cancer. Pharmaceutics 2019, 11, 574. [Google Scholar] [CrossRef]
- Zhao, Y.; Zheng, Y.; Zhu, Y.; Zhang, Y.; Zhu, H.; Liu, T. M1 Macrophage-Derived Exosomes Loaded with Gemcitabine and Deferasirox against Chemoresistant Pancreatic Cancer. Pharmaceutics 2021, 13, 1493. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Tang, C.; Sun, C.; Ying, X.; Shen, R. M1 macrophage-derived exosomes synergistically enhance the anti- bladder cancer effect of gemcitabine. Aging 2022, 14, 7364–7377. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, N.; Wang, J. M1 macrophage-derived exosome-encapsulated cisplatin can enhance its anti-lung cancer effect. Minerva Medica 2023, 114, 634–641. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, H.; Huang, Q.; Peng, C.; Yao, L.; Chen, H.; Qiu, Z.; Wu, Y.; Wang, L.; Chen, W. Exosomes from M1-Polarized Macrophages Enhance Paclitaxel Antitumor Activity by Activating Macrophages-Mediated Inflammation. Theranostics 2019, 9, 1714–1727. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, L.; Tang, M.; Li, H.; Guo, X.; Yang, X. The effects of umbilical cord-derived macrophage exosomes loaded with cisplatin on the growth and drug resistance of ovarian cancer cells. Drug Dev. Ind. Pharm. 2020, 46, 1150–1162. [Google Scholar] [CrossRef]
- Martínez-Santillán, A.; González-Valdez, J. Novel Technologies for Exosome and Exosome-like Nanovesicle Procurement and Enhancement. Biomedicines 2023, 11, 1487. [Google Scholar] [CrossRef]
- Koh, H.B.; Kim, H.J.; Kang, S.W.; Yoo, T.H. Exosome-Based Drug Delivery: Translation from Bench to Clinic. Pharmaceutics 2023, 15, 2042. [Google Scholar] [CrossRef]
- Willis, G.R.; Kourembanas, S.; Mitsialis, S.A. Toward Exosome-Based Therapeutics: Isolation, Heterogeneity, and Fit-for-Purpose Potency. Front. Cardiovasc. Med. 2017, 4, 63. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.C.; Li, P.L.; Zhang, T.Y.; Xu, Z.P.; Huang, X.W.; Wang, R.M.; Du, L.T. Review on Strategies and Technologies for Exosome Isolation and Purification. Front. Bioeng. Biotech. 2022, 9, 811971. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.Y.; Tang, Y.; Cheng, Y.C. Exosomes as Targeted Delivery Drug System: Advances in Exosome Loading, Surface Functionalization and Potential for Clinical Application. Curr. Drug Deliv. 2024, 21, 473–487. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; He, D.; Li, H.Y. Bioglass enhances the production of exosomes and improves their capability of promoting vascularization. Bioact. Mater. 2021, 6, 823–835. [Google Scholar] [CrossRef]
- Yu, J.J.; Wei, Y.; Cui, Z.P.; Tian, J.; Cai, H.B.; Zhang, W.A. Thermosensitive Capturer Coupled with the CD63 Aptamer for Highly Efficient Isolation of Exosomes. Acs Macro Lett. 2024, 13, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Yuan, N.H.; Xiao, L.F.; Chen, J.X.; Liu, B.Y.; Ren, S.Q.; Sheng, X.H.; Qi, X.L.; Wang, Y.Q.; Chen, C.; Guo, K.J.; et al. CREG1 promotes bovine placental trophoblast cells exosome release by targeting IGF2R and participates in regulating organoid differentiation via exosomes transport. Int. J. Biol. Macromol. 2024, 274, 133298. [Google Scholar] [CrossRef] [PubMed]
- Van den Bossche, J.; Baardman, J.; Otto, N.A.; van der Velden, S.; Neele, A.E.; van den Berg, S.M.; Luque-Martin, R.; Chen, H.J.; Boshuizen, M.C.S.; Ahmed, M.; et al. Mitochondrial Dysfunction Prevents Repolarization of Inflammatory Macrophages. Cell Rep. 2016, 17, 684–696. [Google Scholar] [CrossRef] [PubMed]
- Eisel, D.; Das, K.; König, R.; Osen, W.; Eichmüller, S.B. Reprogramming of M2-like macrophages to M1-like phenotype by tumor-antigen specific CD4+T cells. Eur. J. Cancer 2019, 110, S6–S7. [Google Scholar] [CrossRef]
- Fermi, V.; Warta, R.; Wöllner, A.; Lotsch, C.; Jassowicz, L.; Rapp, C.; Knoll, M.; Jungwirth, G.; Jungk, C.; Trong, P.D.; et al. Effective Reprogramming of Patient-Derived M2-Polarized Glioblastoma-Associated Microglia/Macrophages by Treatment with GW2580. Clin. Cancer Res. 2023, 29, 4685–4697. [Google Scholar] [CrossRef]
- Wu, K.T.R.E.; El Zowalaty, A.E.; Sayin, V.I.; Papagiannakopoulos, T. The pleiotropic functions of reactive oxygen species in cancer. Nat. Cancer 2024, 5, 384–399. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, B.M.; Banik, B.K.; Borah, P.; Jain, A. Reactive Oxygen Species (ROS): Key Components in Cancer Therapies. Anti-Cancer Agents 2022, 22, 215–222. [Google Scholar] [CrossRef]
- Lin, S.S.; Li, Y.Y.; Zamyatnin, A.A.; Werner, J.; Bazhin, A.V. Reactive oxygen species and colorectal cancer. J. Cell. Physiol. 2018, 233, 5119–5132. [Google Scholar] [CrossRef] [PubMed]
- Srinivas, U.S.; Tan, B.W.Q.; Vellayappan, B.A.; Jeyasekharan, A.D. ROS and the DNA damage response in cancer. Redox Biol. 2019, 25, 101084. [Google Scholar] [CrossRef] [PubMed]
- Saikolappan, S.; Kumar, B.; Shishodia, G.; Koul, S.; Koul, H.K. Reactive oxygen species and cancer: A complex interaction. Cancer Lett. 2019, 452, 132–143. [Google Scholar] [CrossRef] [PubMed]
- Griess, B.; Datta, K.; Teoh-Fitzgerald, M. Antioxidant Reprogram Macrophages from Pro-Tumor M2 to Anti-Tumor M1. Free Radical Biol. Med. 2016, 100, S56. [Google Scholar] [CrossRef]
- Zhang, B.; Yang, Y.; Yi, J.R.; Zhao, Z.H.; Ye, R. Hyperglycemia modulates M1/M2 macrophage polarization via reactive oxygen species overproduction in ligature-induced periodontitis. J. Periodontal Res. 2021, 56, 991–1005. [Google Scholar] [CrossRef]
- Susser, L.I.; Nguyen, M.A.; Geoffrion, M.; Khacho, M.; Rayner, K.J. Mitochondrial Dynamics Directs Macrophage Polarization During Atherosclerosis: Implications For Disease Regression. Arterioscler. Thromb. Vasc. Biol. 2021, 41, A100. [Google Scholar] [CrossRef]
- Seong, J.B.; Kim, B.; Kim, S.; Kim, M.H.; Park, Y.H.; Lee, Y.; Lee, H.J.; Hong, C.W.; Lee, D.S. Macrophage peroxiredoxin 5 deficiency promotes lung cancer progression via ROS-dependent M2-like polarization. Free Radic. Biol. Med. 2021, 176, 322–334. [Google Scholar] [CrossRef]
- Taufalele, P.V.; Wang, W.J.; Simmons, A.J.; Southard-Smith, A.N.; Chen, B.; Greenlee, J.D.; King, M.R.; Lau, K.S.; Hassane, D.C.; Bordeleau, F.; et al. Matrix stiffness enhances cancer-macrophage interactions and M2-like macrophage accumulation in the breast tumor microenvironment. Acta Biomater. 2023, 163, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Omran, B.; Baek, K.H. Nanoantioxidants: Pioneer Types, Advantages, Limitations, and Future Insights. Molecules 2021, 26, 7031. [Google Scholar] [CrossRef]
- Gu, Z.Y.; Liu, T.Q.; Tang, J.; Yang, Y.N.; Song, H.; Tuong, Z.K.; Fu, J.Y.; Yu, C.Z. Mechanism of Iron Oxide-Induced Macrophage Activation: The Impact of Composition and the Underlying Signaling Pathway. J. Am. Chem. Soc. 2019, 141, 6122–6126. [Google Scholar] [CrossRef]
- Yang, G.; Ni, J.S.; Li, Y.X.; Zha, M.L.; Tu, Y.; Li, K. Acceptor Engineering for Optimized ROS Generation Facilitates Reprogramming Macrophages to M1 Phenotype in Photodynamic Immunotherapy. Angew. Chem. Int. Ed. 2021, 60, 5386–5393. [Google Scholar] [CrossRef]
- Li, X.S.; Lovell, J.F.; Yoon, J.; Chen, X.Y. Clinical development and potential of photothermal and photodynamic therapies for cancer. Nat. Rev. Clin. Oncol. 2020, 17, 657–674. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Diaz, M.; Huang, Y.Y.; Hamblin, M.R. Use of fluorescent probes for ROS to tease apart Type I and Type II photochemical pathways in photodynamic therapy. Methods 2016, 109, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Lu, S.B.; Li, C.; Chen, F.; Ni, J.S.; Zha, M.L.; Li, Y.X.; Gao, J.; Kang, T.Y.; Liu, C.; et al. Type I macrophage activator photosensitizer against hypoxic tumors. Chem. Sci. 2021, 12, 14773–14780. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.Q.; Yuan, H.T.; Wang, P.L.; Yi, L.T.; Wen, Q.L.; Zhou, R.H.; Bai, Y.M.; Ma, J.B.; Wang, X.X.; Liang, Y.T.; et al. Photoimmunotherapy-Induced Pyroptosis Remodels the Microenvironment to Enhance Cancer Immunotherapy. ACS Mater. Lett. 2024, 6, 3750–3762. [Google Scholar] [CrossRef]
- Zheng, Q.F.; Maksimovic, I.; Upad, A.; David, Y. Non-enzymatic covalent modifications: A new link between metabolism and epigenetics. Protein Cell 2020, 11, 401–416. [Google Scholar] [CrossRef]
- Hogg, S.J.; Beavis, P.A.; Dawson, M.A.; Johnstone, R.W. Targeting the epigenetic regulation of antitumour immunity. Nat. Rev. Drug Discov. 2020, 19, 776–800. [Google Scholar] [CrossRef]
- Lu, Y.J.; Chan, Y.T.; Tan, H.Y.; Li, S.; Wang, N.; Feng, Y.B. Epigenetic regulation in human cancer: The potential role of epi-drug in cancer therapy. Mol. Cancer 2020, 19, 79. [Google Scholar] [CrossRef]
- Fontecha-Barriuso, M.; Martin-Sanchez, D.; Ruiz-Andres, O.; Poveda, J.; Sanchez-Niño, M.D.; Valiño-Rivas, L.; Ruiz-Ortega, M.; Ortiz, A.; Sanz, A.B. Targeting epigenetic DNA and histone modifications to treat kidney disease. Nephrol. Dial. Transpl. 2018, 33, 1875–1886. [Google Scholar] [CrossRef]
- Ivashkiv, L.B. Epigenetic regulation of macrophage polarization and function. Trends Immunol. 2013, 34, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Vadevoo, S.M.P.; Gunassekaran, G.R.; Yoo, J.D.; Kwon, T.H.; Hur, K.; Chae, S.; Lee, B. Epigenetic therapy reprograms M2-type tumor-associated macrophages into an M1-like phenotype by upregulating miR-7083-5p. Front. Immunol. 2022, 13, 976196. [Google Scholar] [CrossRef] [PubMed]
- Mougiakakos, D.; Bach, C.; Böttcher, M.; Beier, F.; Röhner, L.; Stoll, A.; Rehli, M.; Gebhard, C.; Lischer, C.; Eberhardt, M.; et al. The IKZF1-IRF4/IRF5 Axis Controls Polarization of Myeloma-Associated Macrophages. Cancer Immunol. Res. 2021, 9, 265–278. [Google Scholar] [CrossRef]
- Li, Y.; Chen, C.; Liu, H.L.; Zhang, Z.F.; Wang, C.L. LARRPM restricts lung adenocarcinoma progression and M2 macrophage polarization through epigenetically regulating LINC00240 and CSF1. Cell. Mol. Biol. Lett. 2022, 27, 91. [Google Scholar] [CrossRef]
- Zhou, Y.N.; Qian, M.Q.; Li, J.L.; Ruan, L.X.; Wang, Y.R.; Cai, C.Y.; Gu, S.X.; Zhao, X.Y. The role of tumor-associated macrophages in lung cancer: From mechanism to small molecule therapy. Biomed. Pharmacother. 2024, 170, 116014. [Google Scholar] [CrossRef]
- Cai, H.; Zhang, Y.C.; Wang, J.; Gu, J.Y. Defects in Macrophage Reprogramming in Cancer Therapy: The Negative Impact of PD-L1/PD-1. Front. Immunol. 2021, 12, 690869. [Google Scholar] [CrossRef]
- Yao, J.J.; Du, Z.H.; Li, Z.B.; Zhang, S.H.; Lin, Y.K.; Li, H.Y.; Zhou, L.; Wang, Y.H.; Yan, G.X.; Wu, X.C.; et al. 6-Gingerol as an arginase inhibitor prevents urethane-induced lung carcinogenesis by reprogramming tumor supporting M2 macrophages to M1 phenotype. Food Funct. 2018, 9, 4611–4620. [Google Scholar] [CrossRef]
- Wanderley, C.W.; Colon, D.F.; Luiz, J.P.M.; Oliveira, F.F.; Viacava, P.R.; Leite, C.A.; Pereira, J.A.; Silva, C.M.; Silva, C.R.; Silva, R.L.; et al. Paclitaxel Reduces Tumor Growth by Reprogramming Tumor-Associated Macrophages to an M1 Profile in a TLR4-Dependent Manner. Cancer Res. 2018, 78, 5891–5900. [Google Scholar] [CrossRef]
- Zhu, W.; Xu, R.; Du, J.; Fu, Y.; Li, S.; Zhang, P.; Liu, L.; Jiang, H. Zoledronic acid promotes TLR-4-mediated M1 macrophage polarization in bisphosphonate-related osteonecrosis of the jaw. FASEB J. 2019, 33, 5208–5219. [Google Scholar] [CrossRef]
- Hernandes, L.C.; Machado, A.R.T.; Tuttis, K.; Ribeiro, D.L.; Aissa, A.F.; Dévoz, P.P.; Antunes, L.M.G. Caffeic acid and chlorogenic acid cytotoxicity, genotoxicity and impact on global DNA methylation in human leukemic cell lines. Genet. Mol. Biol. 2020, 43, e20190347. [Google Scholar] [CrossRef]
- Xue, N.N.; Zhou, Q.; Ji, M.; Jin, J.; Lai, F.F.; Chen, J.; Zhang, M.T.; Jia, J.; Yang, H.R.; Zhang, J.; et al. Chlorogenic acid inhibits glioblastoma growth through repolarizating macrophage from M2 to M1 phenotype. Sci. Rep. 2017, 7, 39011. [Google Scholar] [CrossRef]
- Filbert, E.L.; Bjorck, P.K.; Srivastava, M.K.; Bahjat, F.R.; Yang, X. APX005M, a CD40 agonist antibody with unique epitope specificity and Fc receptor binding profile for optimal therapeutic application. Cancer Immunol. Immunother. 2021, 70, 1853–1865. [Google Scholar] [CrossRef]
- Lim, C.Y.; Chang, J.H.; Lee, W.S.; Kim, J.; Park, I.Y. CD40 Agonists Alter the Pancreatic Cancer Microenvironment by Shifting the Macrophage Phenotype toward M1 and Suppress Human Pancreatic Cancer in Organotypic Slice Cultures. Gut Liver 2022, 16, 645–659. [Google Scholar] [CrossRef]
- Bartneck, M.; Fech, V.; Ehling, J.; Govaere, O.; Warzecha, K.T.; Hittatiya, K.; Vucur, M.; Gautheron, J.; Luedde, T.; Trautwein, C.; et al. Histidine-rich glycoprotein promotes macrophage activation and inflammation in chronic liver disease. Hepatology 2016, 63, 1310–1324. [Google Scholar] [CrossRef]
- Minz, A.P.; Das, B.; Mohapatra, D.; Suresh, V.; Mishra, S.; Senapati, S. Gemcitabine induces polarization of mouse peritoneal macrophages towards M1-like and confers antitumor property by inducing ROS production. Clin. Exp. Metastasis 2022, 39, 783–800. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, M.; Karim, M.R.; Izawa, T.; Kuwamura, M.; Yamate, J. Immunophenotypical Characterization of M1/M2 Macrophages and Lymphocytes in Cisplatin-Induced Rat Progressive Renal Fibrosis. Cells 2021, 10, 257. [Google Scholar] [CrossRef]
- Markham, A. Lurbinectedin: First Approval. Drugs 2020, 80, 1345–1353. [Google Scholar] [CrossRef]
- Chen, W.; Guo, Q.Y.; Zhang, Y.; Liu, Q.W.; Zhang, Y.A.; Zhao, C.F.; Li, X.H.; Bai, X.; Zhang, L.; Shao, S.X. Nocardia rubra cell wall skeleton regulates tumour-associated macrophage polarization by reprogramming M2 macrophages into M1 macrophages via STAT1/STAT6 pathways. Scand. J. Immunol. 2023, 98, e13320. [Google Scholar] [CrossRef] [PubMed]
- Richert, I.; Berchard, P.; Abbes, L.; Novikov, A.; Chettab, K.; Vandermoeten, A.; Dumontet, C.; Karanian, M.; Kerzerho, J.; Caroff, M.; et al. A TLR4 Agonist Induces Osteosarcoma Regression by Inducing an Antitumor Immune Response and Reprogramming M2 Macrophages to M1 Macrophages. Cancers 2023, 15, 4635. [Google Scholar] [CrossRef] [PubMed]
- Griess, B.; Datta, K.; Teoh-Fitzgerald, M. MnTE-2-PyP Inhibits Stat6 Activation and Reprograms Pro-Tumor M2 Macrophages to Anti-Tumor M1. Free Radical Bio Med. 2017, 112, 90. [Google Scholar] [CrossRef]
- Yang, T.; Han, Y.; Chen, J.; Liang, X.; Sun, L. MiR-506 Promotes Antitumor Immune Response in Pancreatic Cancer by Reprogramming Tumor-Associated Macrophages toward an M1 Phenotype. Biomedicines 2023, 11, 2874. [Google Scholar] [CrossRef] [PubMed]
- Zong, S.; Li, J.L.; Ye, Z.Y.; Zhang, X.M.; Yang, L.; Chen, X.; Ye, M. polysaccharide suppresses S180 sarcoma by boosting anti-tumor immune responses and skewing tumor-associated macrophages toward M1 phenotype. Int. J. Biol. Macromol. 2020, 144, 1022–1033. [Google Scholar] [CrossRef] [PubMed]
- Gunassekaran, G.R.; Vadevoo, S.M.P.; Baek, M.C.; Lee, B. M1 macrophage exosomes engineered to foster M1 polarization and target the IL-4 receptor inhibit tumor growth by reprogramming tumor-associated macrophages into M1-like macrophages. Biomaterials 2021, 278, 121137. [Google Scholar] [CrossRef]
- Yu, Y.; Li, T.; Ou, M.; Luo, R.; Chen, H.; Ren, H.; Li, Z.; Sun, J.; Zhang, H.; Peng, S.; et al. OX40L-expressing M1-like macrophage exosomes for cancer immunotherapy. J. Control. Release 2024, 365, 469–479. [Google Scholar] [CrossRef]
- Wu, X.; Cheng, Y.; Zheng, R.; Xu, K.; Yan, J.; Song, P.; Wang, Y.; Rauf, A.; Pan, Y.; Zhang, H. Immunomodulation of Tumor Microenvironment by Arginine-Loaded Iron Oxide Nanoparticles for Gaseous Immunotherapy. ACS Appl. Mater. Interfaces 2021, 13, 19825–19835. [Google Scholar] [CrossRef]
- Zhang, R.Y.; Cheng, K.; Huang, Z.Y.; Zhang, X.S.; Li, Y.; Sun, X.; Yang, X.Q.; Hu, Y.G.; Hou, X.L.; Liu, B.; et al. M1 macrophage-derived exosome for reprograming M2 macrophages and combining endogenous NO gas therapy with enhanced photodynamic synergistic therapy in colorectal cancer. J. Colloid Interf. Sci. 2024, 654, 612–625. [Google Scholar] [CrossRef]
- Zhang, M.H.; Hu, S.Y.; Liu, L.; Dang, P.Y.; Liu, Y.; Sun, Z.Q.; Qiao, B.B.; Wang, C.Z. Engineered exosomes from different sources for cancer-targeted therapy. Signal Transduct. Tar. 2023, 8, 124. [Google Scholar] [CrossRef]
- Kamerkar, S.; Leng, C.; Burenkova, O.; Jang, S.C.; Mccoy, C.; Zhang, K.; Dahlberg, W.; Leblanc, M.; Quillery, H.; Maubant, S.; et al. Engineered exosome- mediated STAT6 knockdown in tumor associated macrophages (TAMs) results in potent single agent activity in a hepatocellular carcinoma (HCC) model. Cancer Res. 2021, 81, 1792. [Google Scholar] [CrossRef]
- Iqbal, Z.; Rehman, K.; Mahmood, A.; Shabbir, M.; Liang, Y.J.; Duan, L.; Zeng, H. Exosome for mRNA delivery: Strategies and therapeutic applications. J. Nanobiotechnol. 2024, 22, 395. [Google Scholar] [CrossRef] [PubMed]
- Huda, M.N.; Nafiujjaman, M.; Deaguero, I.G.; Okonkwo, J.; Hill, M.L.; Kim, T.; Nurunnabi, M. Potential Use of Exosomes as Diagnostic Biomarkers and in Targeted Drug Delivery: Progress in Clinical and Preclinical Applications. ACS Biomater. Sci. Eng. 2021, 7, 2106–2149. [Google Scholar] [CrossRef]
- Si, C.H.; Gao, J.E.; Ma, X. Engineered exosomes in emerging cell-free therapy. Front. Oncol. 2024, 14, 1382398. [Google Scholar] [CrossRef]
- Kittelmann, S.; McGregor, A.P. Modulation and Evolution of Animal Development through microRNA Regulation of Gene Expression. Genes 2019, 10, 321. [Google Scholar] [CrossRef]
- Chen, X.; Ying, X.; Wang, X.J.; Wu, X.L.; Zhu, Q.Y.; Wang, X.P. Exosomes derived from hypoxic epithelial ovarian cancer deliver microRNA-940 to induce macrophage M2 polarization. Oncol. Rep. 2017, 38, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; He, X.J.; Xu, Y.Y.; Zhang, W.; Fu, F. MiR-200b is upregulated in plasma-derived exosomes and functions as an oncogene by promoting macrophage M2 polarization in ovarian cancer. J. Ovarian Res. 2021, 14, 74. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhou, J.R.; Li, X.D.; Wang, X.J.; Lin, Y.Y.; Wang, X.P. Exosomes derived from hypoxic epithelial ovarian cancer cells deliver microRNAs to macrophages and elicit a tumor-promoted phenotype. Cancer Lett. 2018, 435, 80–91. [Google Scholar] [CrossRef]
- Cooks, T.; Pateras, I.S.; Jenkins, L.M.; Patel, K.M.; Robles, A.I.; Morris, J.; Forshew, T.; Appella, E.; Gorgoulis, V.G.; Harris, C.C. Mutant p53 cancers reprogram macrophages to tumor supporting macrophages via exosomal miR-1246. Nat. Commun. 2018, 9, 771. [Google Scholar] [CrossRef]
- Yang, S.J.; Li, J.; Chen, Y.; Hui, R.T.; Zhang, W.L. MicroRNA-216a Promotes M1 Macrophages Polarization Through the Activation of Telomerase. Hypertension 2017, 70, A146. [Google Scholar] [CrossRef]
- Wang, L.; Hu, Y.Y.; Zhao, J.L.; Huang, F.; Liang, S.Q.; Dong, L.; Chen, Y.; Yu, H.C.; Bai, J.; Yang, J.M.; et al. Targeted delivery of miR-99b reprograms tumor-associated macrophage phenotype leading to tumor regression. J. Immunother. Cancer 2020, 8, e000517. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wu, X.; Shen, P.; Wang, J.; Shen, Y.D.; Shen, Y.; Webster, T.J.; Deng, J.J. Applications of Inorganic Nanomaterials in Photothermal Therapy Based on Combinational Cancer Treatment. Int. J. Nanomed. 2020, 15, 1903–1914. [Google Scholar] [CrossRef]
- Aleckovic, M.; Li, Z.Q.; Zhou, N.X.; Qiu, X.T.; Lulseged, B.; Foidart, P.; Huang, X.Y.; Garza, K.; Shu, S.K.; Kesten, N.; et al. Combination Therapies to Improve the Efficacy of Immunotherapy in Triple-negative Breast Cancer. Mol. Cancer Ther. 2023, 22, 1304–1318. [Google Scholar] [CrossRef]
- Jung, S.; Heo, S.; Oh, Y.; Park, K.; Park, S.; Choi, W.; Kim, Y.H.; Jung, S.Y.; Hong, J. Zwitterionic Inhaler with Synergistic Therapeutics for Reprogramming of M2 Macrophage to Pro-Inflammatory Phenotype. Adv. Healthc. Mater. 2023, 12, 2300226. [Google Scholar] [CrossRef]
- Sun, J.H.; Liang, X.L.; Cai, M.Y.; Yan, L.B.; Chen, Z.J.; Guo, L.; Jing, L.; Wang, Y.P.; Zhou, D.F. Protein-Crowned Micelles for Targeted and Synergistic Tumor-Associated Macrophage Reprogramming to Enhance Cancer Treatment. Nano Lett. 2022, 22, 4410–4420. [Google Scholar] [CrossRef]
- Tan, S.M.; Xia, L.Z.; Yi, P.; Han, Y.Q.; Tang, L.; Pan, Q.; Tian, Y.T.; Rao, S.; Oyang, L.; Liang, J.X.; et al. Exosomal miRNAs in tumor microenvironment. J. Exp. Clin. Canc Res. 2020, 39, 67. [Google Scholar] [CrossRef]
- Najafi, M.; Goradel, N.H.; Farhood, B.; Salehi, E.; Nashtaei, M.S.; Khanlarkhani, N.; Khezri, Z.; Majidpoor, J.; Abouzaripour, M.; Habibi, M.; et al. Macrophage polarity in cancer: A review. J. Cell. Biochem. 2019, 120, 2756–2765. [Google Scholar] [CrossRef]
- Liu, J.Y.; Geng, X.F.; Hou, J.X.; Wu, G.S. New insights into M1/M2 macrophages: Key modulators in cancer progression. Cancer Cell Int. 2021, 21, 389. [Google Scholar] [CrossRef]
- Nowicki, K.W.; Hosaka, K.; Walch, F.J.; Scott, E.W.; Hoh, B.L. M1 macrophages are required for murine cerebral aneurysm formation. J. NeuroInterventional Surg. 2018, 10, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Liu, X.Y.; Zhang, Q.; Liu, H.; Zhang, P.; Tian, Z.B.; Zhang, C.P.; Li, X.Y. Crosstalk Among YAP, LncRNA, and Tumor-Associated Macrophages in Tumorigenesis Development. Front. Oncol. 2022, 11, 810893. [Google Scholar] [CrossRef] [PubMed]
- Li, S.S.; Yu, J.L.; Huber, A.; Kryczek, I.; Wang, Z.W.; Jiang, L.; Li, X.; Du, W.; Li, G.P.; Wei, S.; et al. Metabolism drives macrophage heterogeneity in the tumor microenvironment. Cell Rep. 2022, 39, 110609. [Google Scholar] [CrossRef]
- Wang, X.M.; Xu, Y.W.; Sun, Q.; Zhou, X.T.; Ma, W.Z.; Wu, J.B.; Zhuang, J.; Sun, C.G. New insights from the single-cell level: Tumor associated macrophages heterogeneity and personalized therapy. Biomed. Pharmacother. 2022, 153, 113343. [Google Scholar] [CrossRef]
- Yang, S.; Wang, M.Y.; Hua, Y.Z.; Li, J.Y.; Zheng, H.J.; Cui, M.; Huang, N.; Liu, Q.F.; Liao, Q. Advanced insights on tumor-associated macrophages revealed by single-cell RNA sequencing: The intratumor heterogeneity, functional phenotypes, and cellular interactions. Cancer Lett. 2024, 584, 216610. [Google Scholar] [CrossRef] [PubMed]
- Laviron, M.; Petit, M.; Weber-Delacroix, E.; Combes, A.J.; Arkal, A.R.; Barthélémy, S.; Courau, T.; Hume, D.A.; Combadière, C.; Krummel, M.F.; et al. Tumor-associated macrophage heterogeneity is driven by tissue territories in breast cancer. Cell Rep. 2022, 39, 110865. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.A.; Qiu, W.J.; Ling, Y.G.; Qi, X.L.; Liu, J.; Yang, H.; Chu, L.Z. The role of tumor-associated macrophages in glioma cohort: Through both traditional RNA sequencing and single cell RNA sequencing. Front. Oncol. 2023, 13, 1249448. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer Types | Delivery Drug Types |
|---|---|
| Pancreatic cancer | GEM+ DFX [74] |
| Breast cancer | PTX [77], DTX [69] |
| Ovarian cancer | Cisplatin [70,78] |
| Lung cancer | Cisplatin [76], microRNA-let-7b-5p-GNG5 axis [55] |
| Bladder cancer | GEM [75] |
| Methods | Principles | References |
|---|---|---|
| Antioxidants | Reducing levels of extracellular oxidation capable of promoting M2 macrophage production using antioxidants. | [90,91,92,93,94,95,96,97,98,99,100] |
| Photodynamic synergistic therapy | The extracellular ROS generated by type I photosensitizers plays a role. | [101,102,103,104,105,106] |
| Epigenetic therapy | Regulation of chromatin structure and gene expression using epigenetic approaches. | [107,108,109,110,111,112,113,114] |
| Natural materials and synthetic drugs | Targeting molecules and signaling pathways that play crucial roles in macrophage polarization. | [115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136] |
| Engineering EVs | EVs are modified and encapsulated with drugs and then targeted to M2 macrophages. | [137,138,139,140,141] |
| MicroRNAs | MicroRNA affects the polarization state of macrophages by regulating gene expression. | [53,142,143,144,145,146,147,148] |
| Synergistic effects | Multiple methods are combined to achieve reprogramming of M2 macrophages for better anti-cancer effects. | [149,150,151,152] |
| Functional Molecules | Targets |
|---|---|
| 6-Gingerol [117] | Arginase |
| Paclitaxel [118] | TLR4 |
| ZA [119] | TLR4 |
| CHA [121] | JAK-STAT1 and NF-κB pathways |
| CD40 agonists [123] | CD40 receptor |
| HRG [124] | Toll-like receptor |
| Nocardia rubra cell wall skeleton [128] | STAT1/STAT6 pathways |
| TLR4 Agonist [129] | TLR4 |
| MnTE-2-PyP [130] | Stat6 |
| MiR-506 [131] | STAT3 |
| Lachnum polysaccharide [132] | TLR4 |
| M1 EVs engineered to carry NF-κB p50 siRNA and miR-511–3p [133] | IL4R |
| Lipo-MP-LPS [129] | TLR4 |
| OX40L M1-exos [134] | OX40/OX40L pathway |
| 5-aza-dC and TSA [112] | miR-7083-5p |
| LPFe3O4 NPs [135] | NO |
| IL4R-Exo [133] | IL4R |
| M1-EVs nanoprobe [136] | ROS, NO |
| The iron oxide nanoparticles combined with LOX [18] | Carboxylic acid |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, W.; Yang, F.; Zhang, X. Roles of M1 Macrophages and Their Extracellular Vesicles in Cancer Therapy. Cells 2024, 13, 1428. https://doi.org/10.3390/cells13171428
Zhou W, Yang F, Zhang X. Roles of M1 Macrophages and Their Extracellular Vesicles in Cancer Therapy. Cells. 2024; 13(17):1428. https://doi.org/10.3390/cells13171428
Chicago/Turabian StyleZhou, Wenli, Fengtang Yang, and Xiuzhen Zhang. 2024. "Roles of M1 Macrophages and Their Extracellular Vesicles in Cancer Therapy" Cells 13, no. 17: 1428. https://doi.org/10.3390/cells13171428