
Impact of RBMS 3 Progression on Expression of EMT Markers

 , ,
, ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients Cohort
2.2. Immunohistochemistry
2.3. Cell Lines
2.4. Lentiviral Transduction of Breast Cancer Cell Lines
2.4.1. Overexpressing RBMS 3 In Vitro Model
2.4.2. Silencing RBMS 3 In Vitro Model
2.5. Migration Assay
2.6. Immunofluorescence Reactions
2.7. Real-Time PCR
2.8. Western Blotting
2.9. Statistical Analysis
3. Results
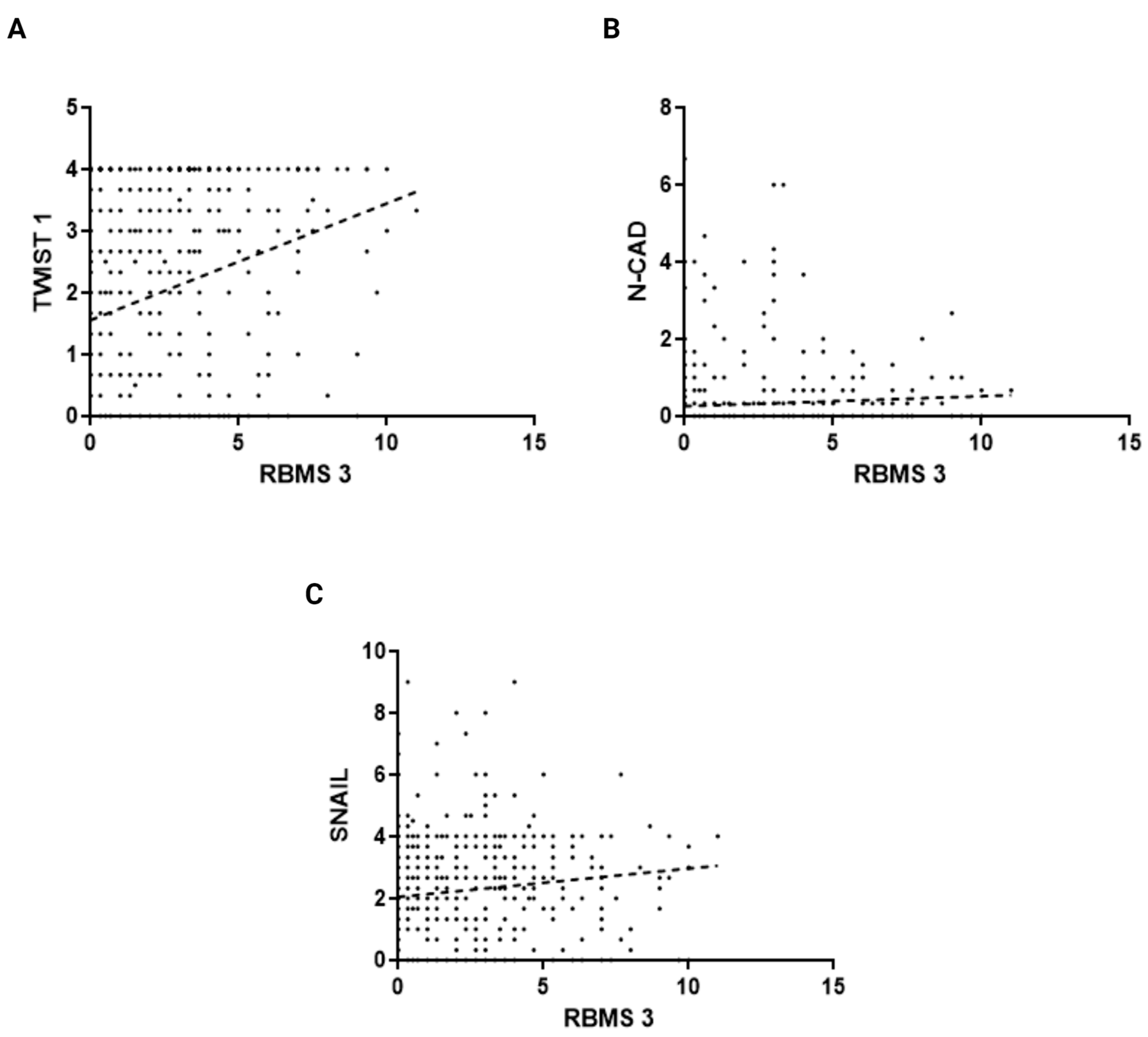
3.1. The Immunohistochemical Intensity of RBMS 3 Expression in Cancer Cells Correlates with Expression of Common EMT Markers in Invasive Ductal Breast Cancer (IDC) Tissue Samples
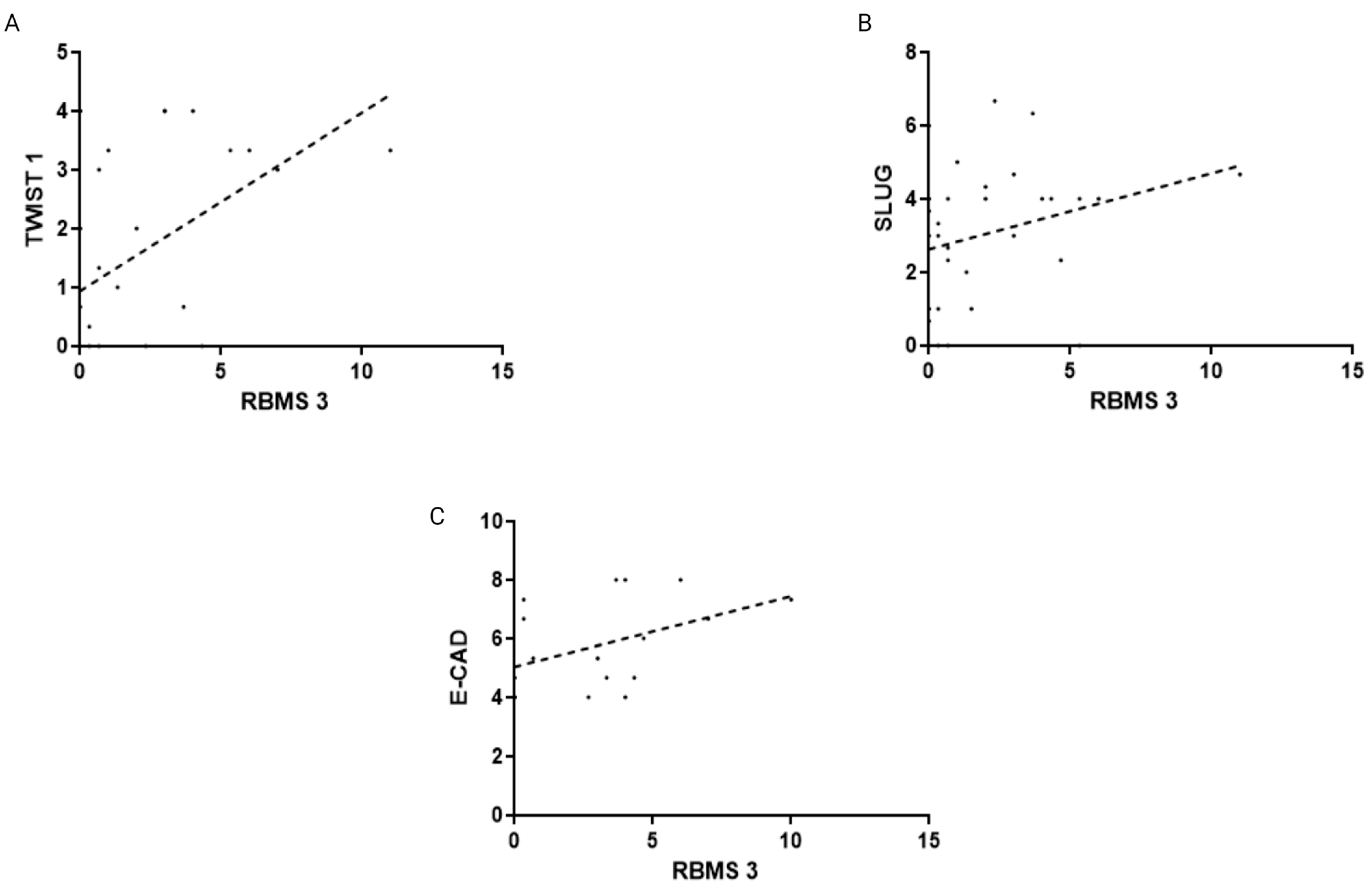
3.2. The Immunohistochemical Intensity of RBMS 3 Expression Displays Different Correlations with Common EMT Markers Depending on the Molecular Subtype of the Tumor
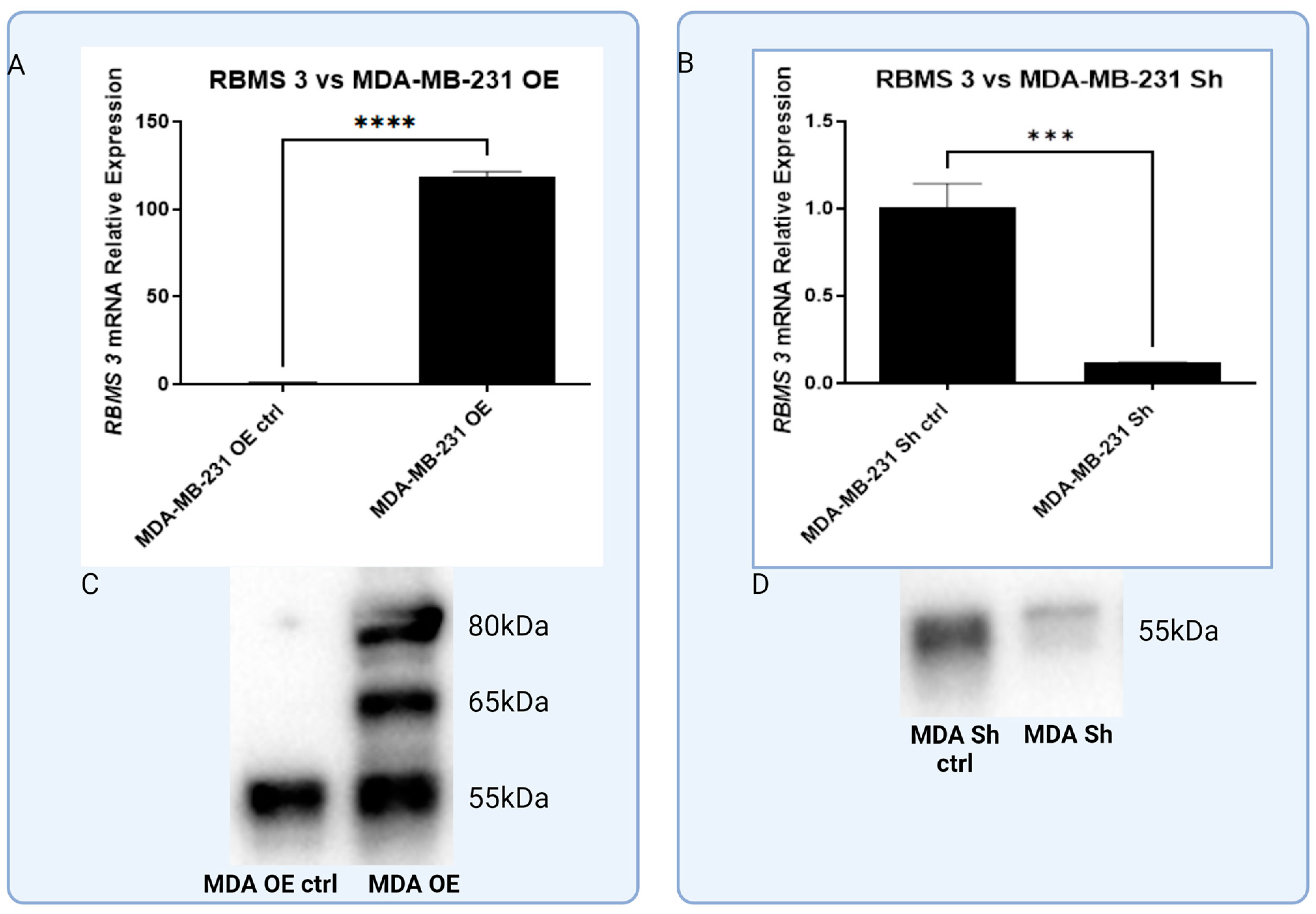
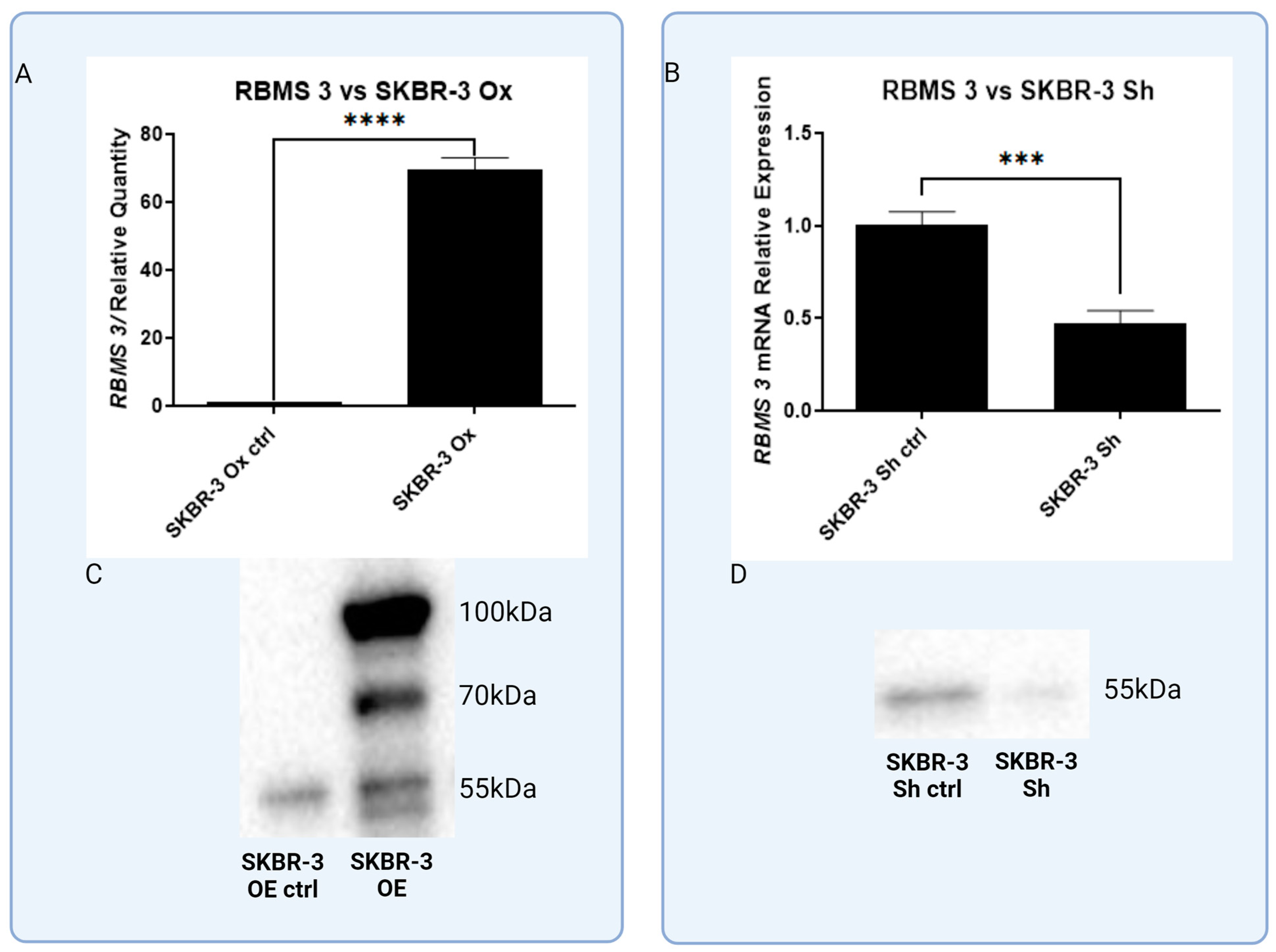
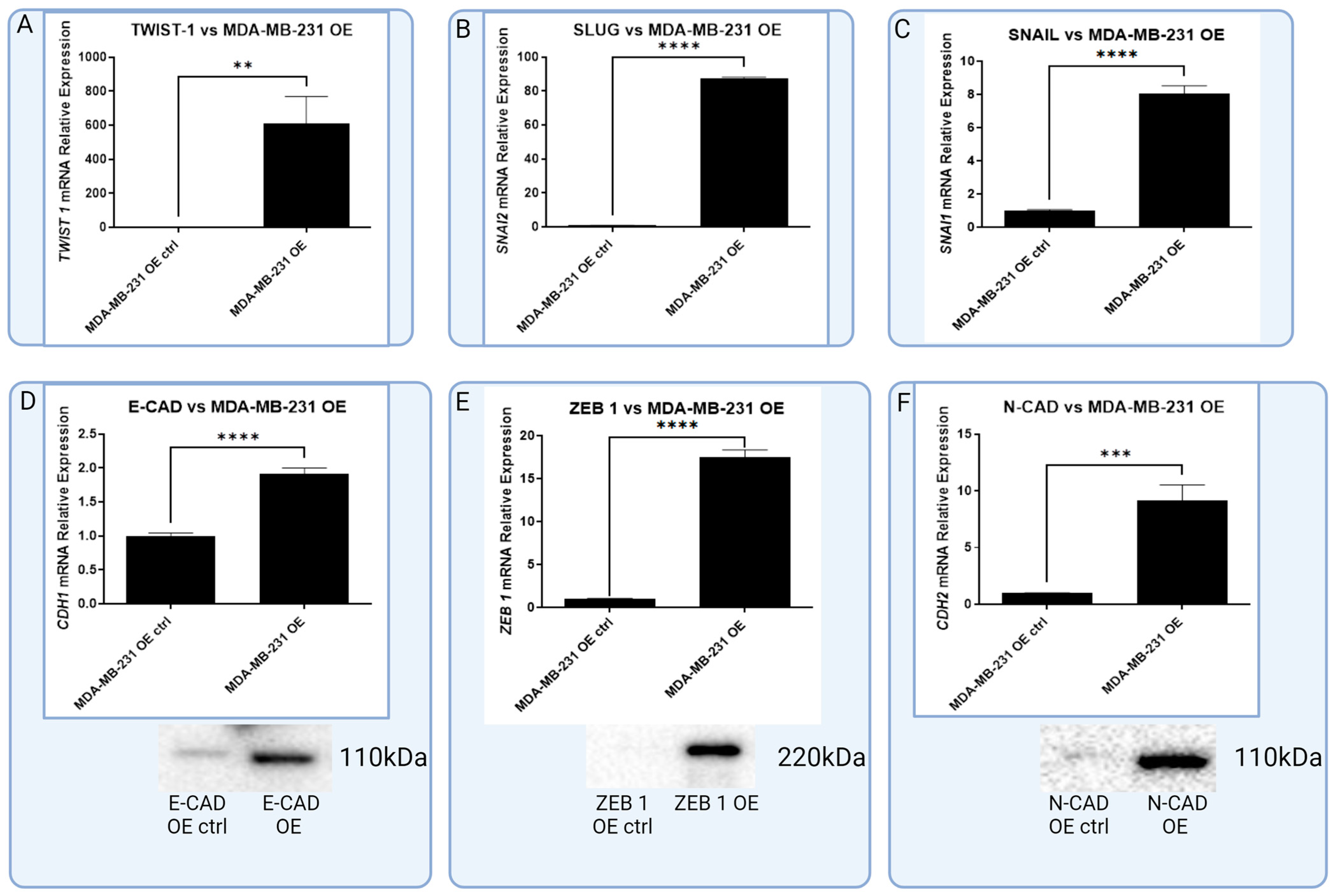
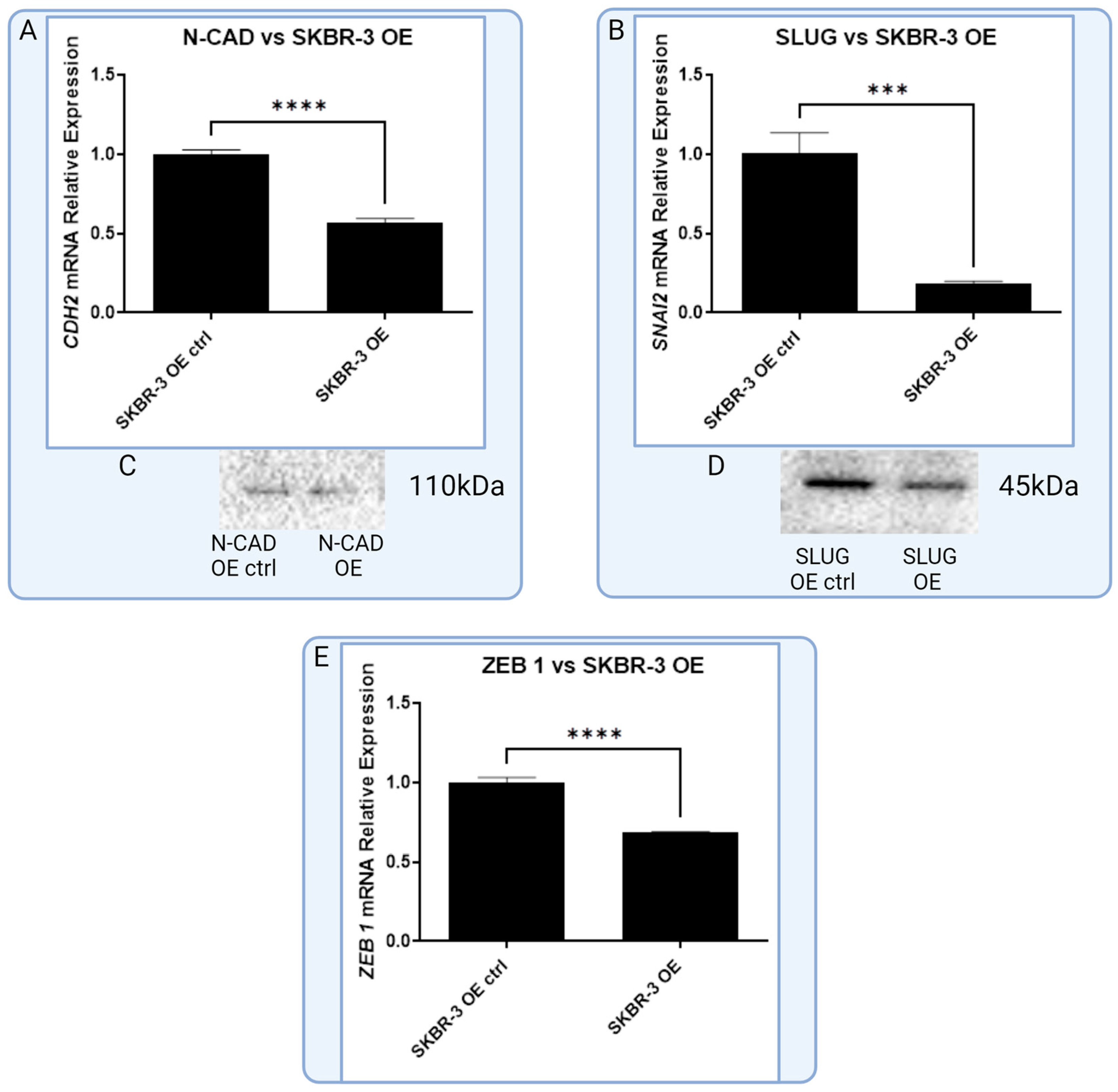
3.3. In Vitro Models of Triple Negative and HER-2-Positive Breast Cancer Cell Lines with Overexpression of RBMS 3 Lead to Observation of Additional Specific Bands
3.4. In Vitro Model of RBMS 3 Overexpression Unveils Differences in RBMS 3 Impacts on EMT Markers between Molecular Subtypes
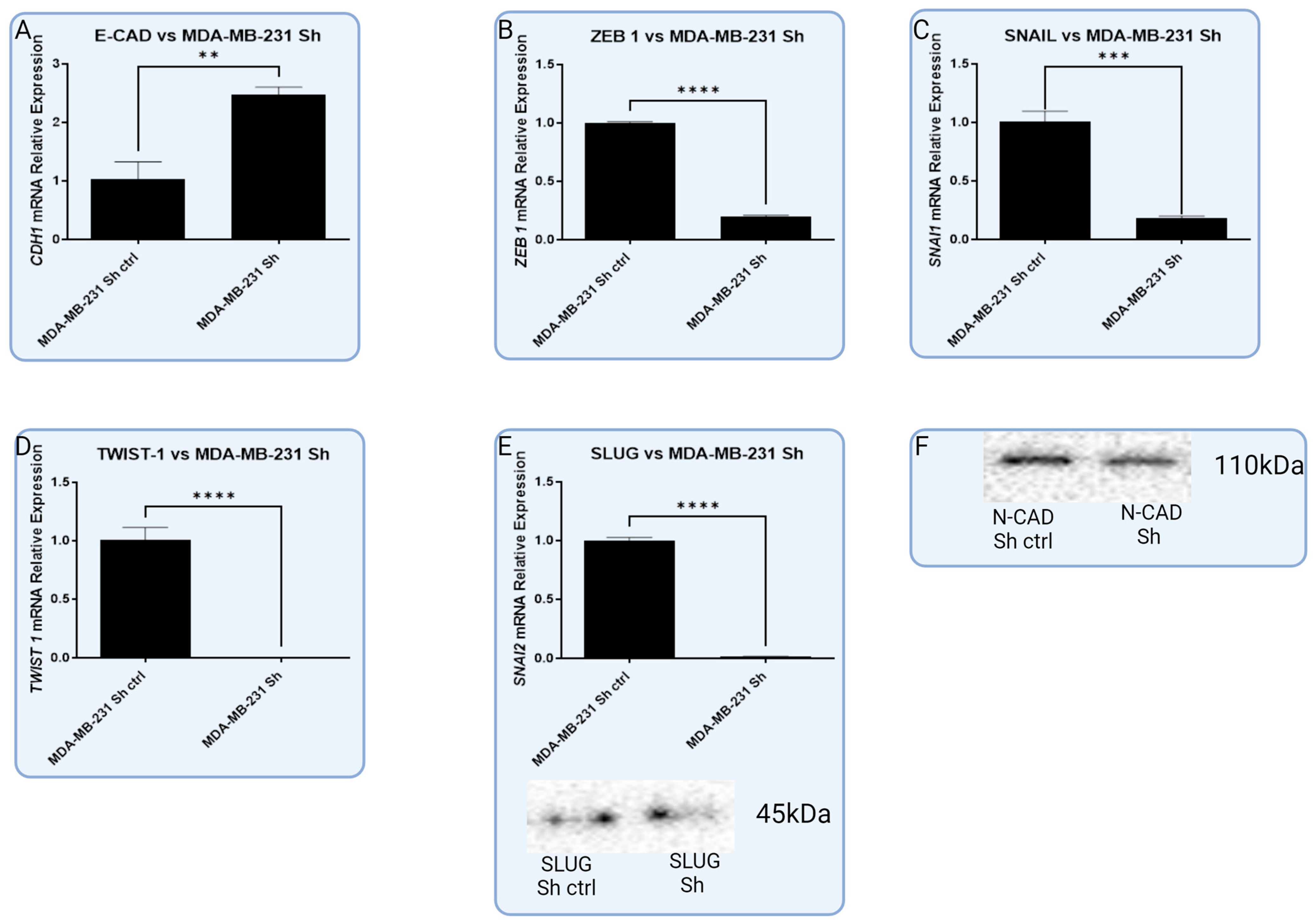
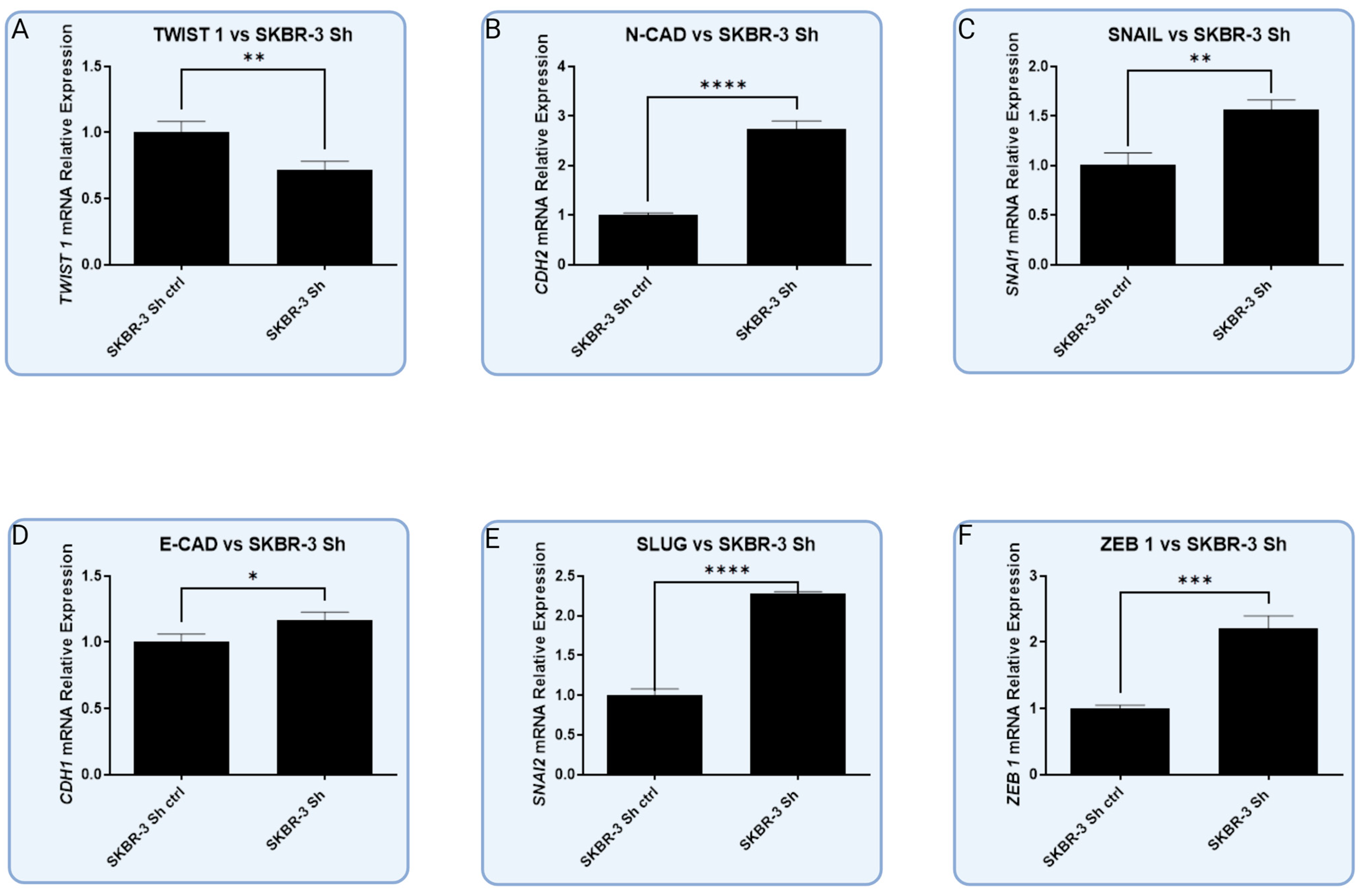
3.5. In Vitro Models with Lentiviral-Silenced RBMS 3 Expression Impact Common Markers of EMT Differently Depending on the Molecular Subtype
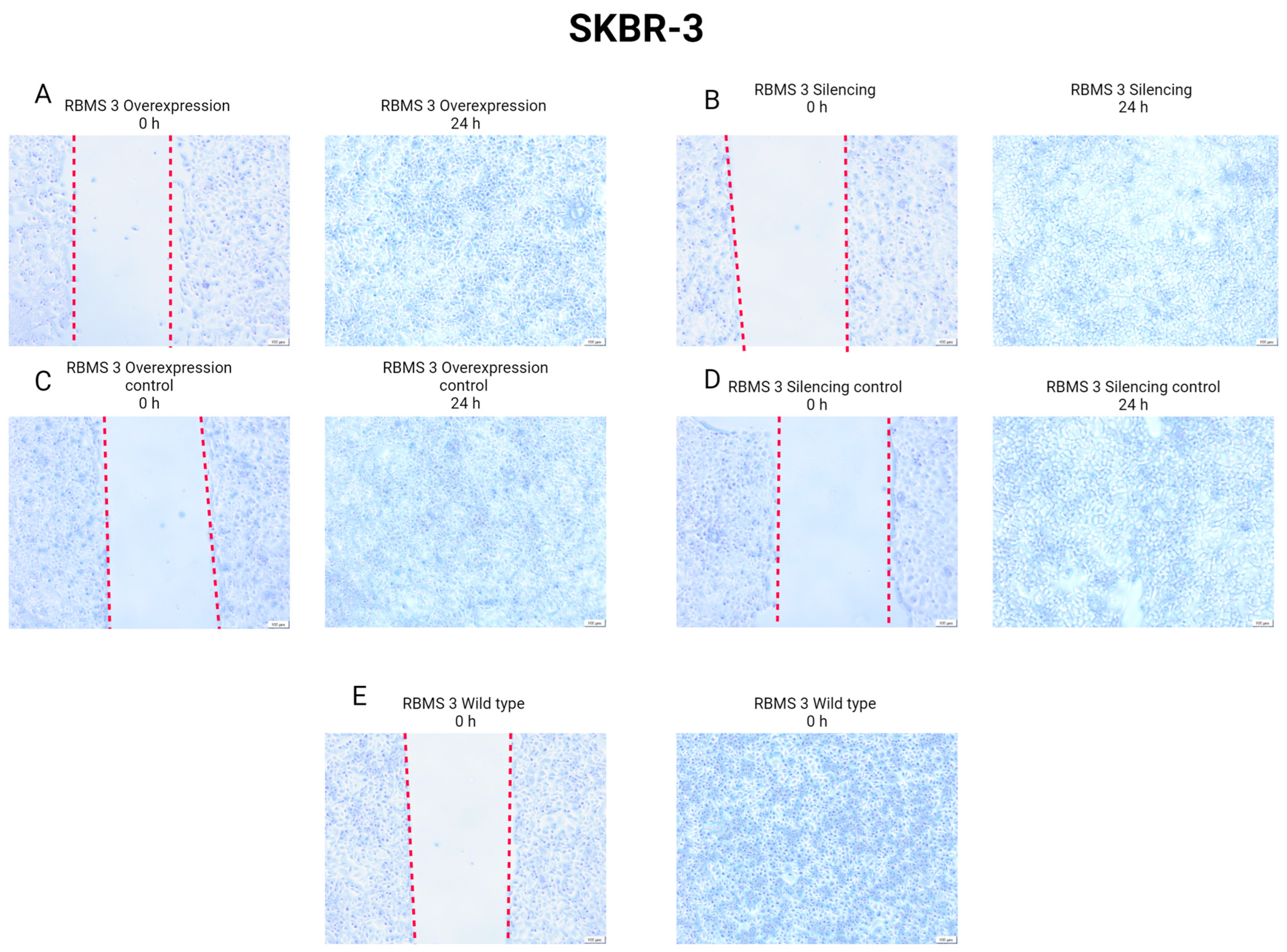
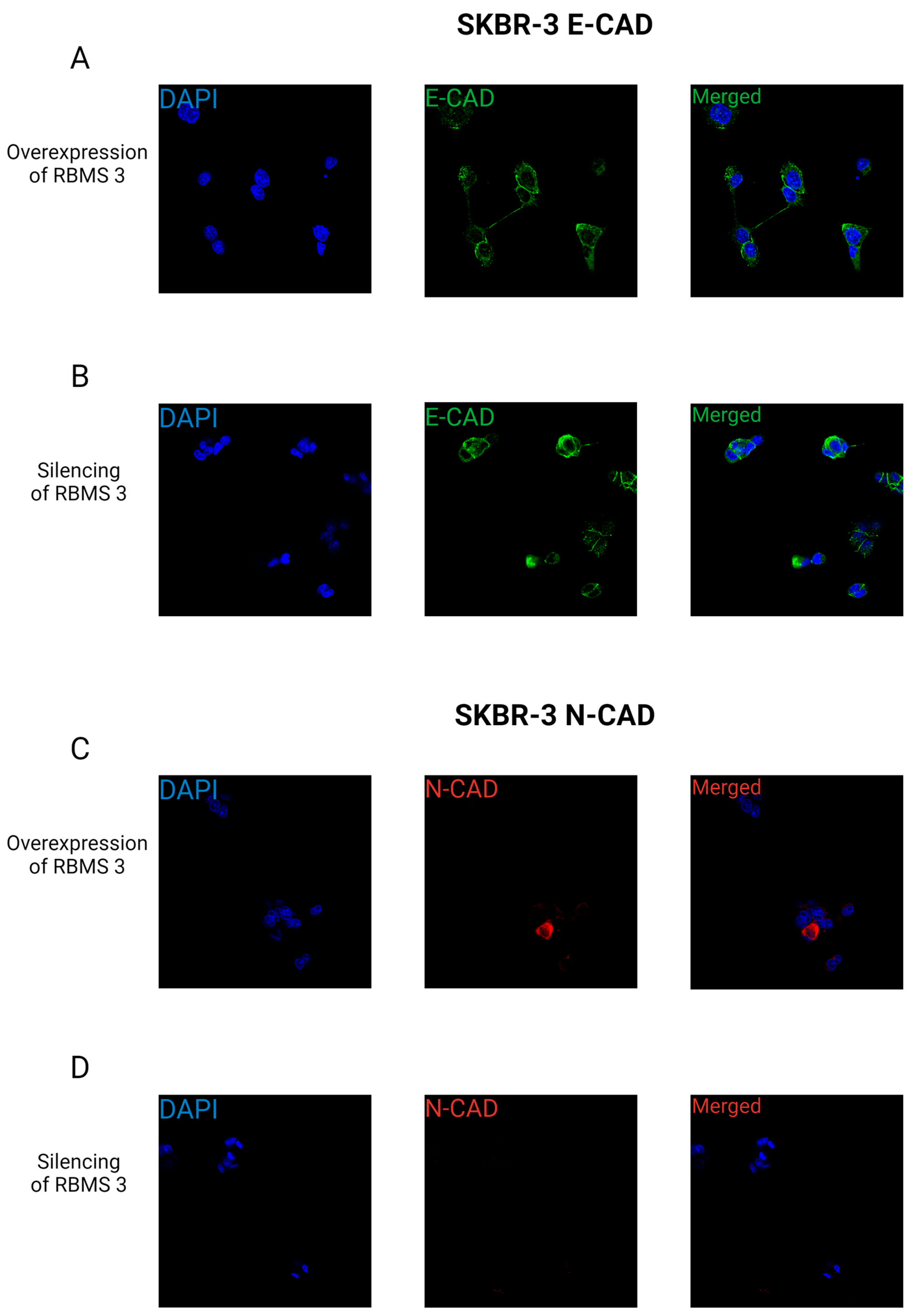
3.6. Changes in RBMS 3 Expression Impact Motility and the Localization of E-CAD and N-CAD in Studied Cells
4. Discussion
Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022, GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef] [PubMed]
- Global Burden of Disease 2019 Cancer Collaboration; Kocarnik, J.M.; Compton, K.; Dean, F.E.; Fu, W.; Gaw, B.L.; Harvey, J.D.; Henrikson, H.J.; Lu, D.; Pennini, A.; et al. Cancer Incidence, Mortality, Years of Life Lost, Years Lived With Disability, and Disability-Adjusted Life Years for 29 Cancer Groups From 2010 to 2019, A Systematic Analysis for the Global Burden of Disease Study 2019. JAMA Oncol. 2022, 8, 420–444. [Google Scholar] [CrossRef]
- Chen, S.; Cao, Z.; Prettner, K.; Kuhn, M.; Yang, J.; Jiao, L.; Wang, Z.; Li, W.; Geldsetzer, P.; Bärnighausen, T.; et al. Estimates and Projections of the Global Economic Cost of 29 Cancers in 204 Countries and Territories From 2020 to 2050. JAMA Oncol. 2023, 9, 465–472. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Loibl, S.; Poortmans, P.; Morrow, M.; Denkert, C.; Curigliano, G. Breast cancer. Lancet 2021, 397, 1750–1769, Erratum in Lancet 2021, 397, 1710. https://doi.org/10.1016/S0140-6736(21)00838-2. [Google Scholar] [CrossRef] [PubMed]
- Łukasiewicz, S.; Czeczelewski, M.; Forma, A.; Baj, J.; Sitarz, R.; Stanisławek, A. Breast Cancer-Epidemiology, Risk Factors, Classification, Prognostic Markers, and Current Treatment Strategies—An Updated Review. Cancers 2021, 13, 4287. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zagami, P.; Carey, L.A. Triple negative breast cancer: Pitfalls and progress. NPJ Breast Cancer 2022, 8, 95. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Waks, A.G.; Winer, E.P. Breast Cancer Treatment: A Review. JAMA 2019, 321, 288–300. [Google Scholar] [CrossRef] [PubMed]
- Aouad, P.; Quinn, H.M.; Berger, A.; Brisken, C. Tumor dormancy: EMT beyond invasion and metastasis. Genesis 2024, 62, e23552. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Weinberg, R.A. Epithelial-to-mesenchymal transition in cancer: Complexity and opportunities. Front. Med. 2018, 12, 361–373. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bakir, B.; Chiarella, A.M.; Pitarresi, J.R.; Rustgi, A.K. EMT, MET, Plasticity, and Tumor Metastasis. Trends Cell Biol. 2020, 30, 764–776. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Greenburg, G.; Hay, E.D. Epithelia suspended in collagen gels can lose polarity and express characteristics of migrating mesenchymal cells. J. Cell Biol. 1982, 95, 333–339. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lachat, C.; Peixoto, P.; Hervouet, E. Epithelial to Mesenchymal Transition History: From Embryonic Development to Cancers. Biomolecules 2021, 11, 782. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428, Erratum in J. Clin. Investig. 2010, 120, 1786.. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Cho, E.S.; Kang, H.E.; Kim, N.H.; Yook, J.I. Therapeutic implications of cancer epithelial-mesenchymal transition (EMT). Arch. Pharm. Res. 2019, 42, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Tamma, R.; Annese, T. Epithelial-Mesenchymal Transition in Cancer: A Historical Overview. Transl. Oncol. 2020, 13, 100773. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Brabletz, T.; Kalluri, R.; Nieto, M.A.; Weinberg, R.A. EMT in cancer. Nat. Rev. Cancer 2018, 18, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Grzegrzolka, J.; Biala, M.; Wojtyra, P.; Kobierzycki, C.; Olbromski, M.; Gomulkiewicz, A.; Piotrowska, A.; Rys, J.; Podhorska-Okolow, M.; Dziegiel, P. Expression of EMT Markers SLUG and TWIST in Breast Cancer. Anticancer Res. 2015, 35, 3961–3968. [Google Scholar] [PubMed]
- Dongre, A.; Weinberg, R.A. New insights into the mechanisms of epithelial-mesenchymal transition and implications for cancer. Nat. Rev. Mol. Cell Biol. 2019, 20, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Penkov, D.; Ni, R.; Else, C.; Piñol-Roma, S.; Ramirez, F.; Tanaka, S. Cloning of a human gene closely related to the genes coding for the c-myc single-strand binding proteins. Gene 2000, 243, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Block, C.J.; Mitchell, A.V.; Wu, L.; Glassbrook, J.; Craig, D.; Chen, W.; Dyson, G.; DeGracia, D.; Polin, L.; Ratnam, M.; et al. GRNA binding protein RBMS3 is a common EMT effector that modulates triple-negative breast cancer progression via stabilizing PRRX1, mRNA. Oncogene 2021, 40, 6430–6442. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Xue, W.; Zhang, J.; Zhu, Y.; Huang, W. Identify Functional lncRNAs in Nonalcoholic Fatty Liver Disease by Constructing a ceRNA Network. ACS Omega 2022, 7, 22522–22530. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wang, J.J.; Liu, X.Y.; Du, W.; Liu, J.Q.; Sun, B.; Zheng, Y.P. RBMS3 delays disc degeneration by inhibiting Wnt/β-catenin signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Singh, S.; Chen, Y.; Hamadeh, I.S.; Langaee, T.; McDonough, C.W.; Holliday, L.S.; Lamba, J.K.; Moreb, J.S.; Katz, J.; et al. Pharmacogenomics of osteonecrosis of the jaw. Bone 2019, 124, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Yin, T.; Zhang, Y.; Zhao, Y.; Zhang, X.; Han, S.; Wang, Y.; Yang, B. Tumor suppressor function of RBMS3 overexpression in EOC associated with immune cell infiltration. Heliyon 2024, 10, e30603. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Li, Y.; Wang, S.; Li, G.; Gao, C.; Cui, Z.; Cong, M.; Hu, J.; Zhang, M.; Jin, X.; Sun, H.; et al. The RNA-binding protein RBMS3 inhibits the progression of colon cancer by regulating the stability of LIMS1 mRNA. Cancer Med. 2024, 13, e7129. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lv, S.L.; Zhou, X.; Li, Y.J.; Luo, L.Y.; Huang, D.Q. RBMS3, a downstream target of AMPK, Exerts Inhibitory Effects on Invasion and Metastasis of Lung Cancer. J. Cancer 2023, 14, 2784–2797. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Li, X.; Chu, X.; Wang, H.; Xiang, L.; Yue, X. Serum exosomal miR-3662 down-regulates the expression of RBMS3 to promote malignant progression and gemcitabine resistance of breast cancer cells. Chem. Biol. Drug Des. 2024, 103, e14488. [Google Scholar] [CrossRef] [PubMed]
- Mi, Y.; Dong, M.; Zuo, X.; Cao, Q.; Gu, X.; Mi, H.; Xiao, F. Genome-wide identification and analysis of epithelial-mesenchymal transition-related RNA-binding proteins and alternative splicing in a human breast cancer cell line. Sci. Rep. 2024, 14, 11753. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Rusak, A.; Jablonska, K.; Piotrowska, A.; Grzegrzolka, J.; Wojnar, A.; Dziegiel, P. Correlation of Expression of CHI3L1 and Nogo-A and their Role in Angiogenesis in Invasive Ductal Breast Carcinoma. Anticancer Res. 2019, 39, 2341–2350. [Google Scholar] [CrossRef] [PubMed]
- Rusak, A.; Buzalewicz, I.; Mrozowska, M.; Wiatrak, B.; Haczkiewicz-Leśniak, K.; Olbromski, M.; Kmiecik, A.; Krzyżak, E.; Pietrowska, A.; Moskal, J.; et al. Multimodal study of CHI3L1 inhibition and its effect on angiogenesis, migration, immune response and refractive index of cellular structures in glioblastoma. Biomed. Pharmacother. 2023, 161, 114520. [Google Scholar] [CrossRef] [PubMed]
- Remmele, W.; Stegner, H.E. Vorschlag zur einheitlichen Definition eines Immunreaktiven Score (IRS) für den immunhistochem-ischen Ostrogenrezeptor-Nachweis (ER-ICA) im Mammakarzinomgewebe [Recommendation for uniform definition of an immunoreactive score (IRS) for immunohistochemical estrogen receptor detection (ER-ICA) in breast cancer tissue]. Pathologe 1987, 8, 138–140. (In German) [Google Scholar] [PubMed]
- Berg, S.; Kutra, D.; Kroeger, T.; Straehle, C.N.; Kausler, B.X.; Haubold, C.; Schiegg, M.; Ales, J.; Beier, T.; Rudy, M.; et al. ilastik: Interactive machine learning for (bio)image analysis. Nat. Methods 2019, 16, 1226–1232. [Google Scholar] [CrossRef] [PubMed]
- Diller, T.; Thompson, J.; Steer, B. Biological validation of a novel process and product for quantitating western blots. J. Biotechnol. 2021, 326, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, D.S.; Karthikeyan, S.K.; Korla, P.K.; Patel, H.; Shovon, A.R.; Athar, M.; Netto, G.J.; Qin, Z.S.; Kumar, S.; Manne, U.; et al. UALCAN: An update to the integrated cancer data analysis platform. Neoplasia 2022, 25, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Rodriguez, I.P.; Chakravarthi, B.V.S.K.; Varambally, S. UALCAN: A portal for facilitating tumor subgroup gene expression and survival analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Górnicki, T.; Lambrinow, J.; Mrozowska, M.; Romanowicz, H.; Smolarz, B.; Piotrowska, A.; Gomułkiewicz, A.; Podhorska-Okołów, M.; Dzięgiel, P.; Grzegrzółka, J. Expression of RBMS3 in Breast Cancer Progression. Int. J. Mol. Sci. 2023, 24, 2866. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Dong, S.; Ma, M.; Li, M.; Guo, Y.; Zuo, X.; Gu, X.; Zhang, M.; Shi, Y. LncRNA MEG3 regulates breast cancer proliferation and apoptosis through miR-141-3p/RBMS3 axis. Genomics 2021, 113, 1689–1704. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Liang, Z.; Xia, Y.; Li, S.; Liang, J.; Hu, Z.; Tang, C.; Zhao, Q.; Gong, Q.; Ouyang, Y. Disruption of RBMS3 suppresses PD-L1 and enhances antitumor immune activities and therapeutic effects of auranofin against triple-negative breast cancer. Chem. Biol. Interact. 2023, 369, 110260. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Quan, L.; Ling, Y. RBMS3 Inhibits the Proliferation and Metastasis of Breast Cancer Cells. Oncol. Res. 2018, 26, 9–15. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhu, L.; Xi, P.W.; Li, X.X.; Sun, X.; Zhou, W.B.; Xia, T.S.; Shi, L.; Hu, Y.; Ding, Q.; Wei, J.F. The RNA binding protein RBMS3 inhibits the metastasis of breast cancer by regulating Twist1 expression. J. Exp. Clin. Cancer Res. 2019, 38, 105, Erratum in J. Exp. Clin. Cancer Res. 2020, 39, 21. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Orrantia-Borunda, E.; Anchondo-Nuñez, P.; Acuña-Aguilar, L.E.; Gómez-Valles, F.O.; Ramírez-Valdespino, C.A. Subtypes of Breast Cancer. In Breast Cancer [Internet]; Mayrovitz, H.N., Ed.; Chapter 3; Exon Publications: Brisbane, Austrilia, 2022. Available online: https://www.ncbi.nlm.nih.gov/books/NBK583808/ (accessed on 6 July 2024).
- Padmanaban, V.; Krol, I.; Suhail, Y.; Szczerba, B.M.; Aceto, N.; Bader, J.S.; Ewald, A.J. E-cadherin is required for metastasis in multiple models of breast cancer. Nature 2019, 573, 439–444. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Nieman, M.T.; Prudoff, R.S.; Johnson, K.R.; Wheelock, M.J. N-cadherin promotes motility in human breast cancer cells regardless of their E-cadherin expression. J. Cell Biol. 1999, 147, 631–644. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Friedl, P. Prespecification and plasticity: Shifting mechanisms of cell migration. Curr. Opin. Cell Biol. 2004, 16, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, M.; Christofori, G.; Lehembre, F. Distinct mechanisms of tumor invasion and metastasis. Trends Mol. Med. 2007, 13, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Tedja, R.; Roberts, C.M.; Alvero, A.B.; Cardenas, C.; Yang-Hartwich, Y.; Spadinger, S.; Pitruzzello, M.; Yin, G.; Glackin, C.A.; Mor, G. Protein kinase Cα-mediated phosphorylation of Twist1 at Ser-144 prevents Twist1 ubiquitination and stabilizes it. J. Biol. Chem. 2019, 294, 5082–5093. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Viñas-Castells, R.; Frías, Á.; Robles-Lanuza, E.; Zhang, K.; Longmore, G.D.; García de Herreros, A.; Díaz, V.M. Nuclear ubiquitination by FBXL5 modulates Snail1 DNA binding and stability. Nucleic Acids Res. 2014, 42, 1079–1094. [Google Scholar] [CrossRef]
- Wang, S.P.; Wang, W.L.; Chang, Y.L.; Wu, C.T.; Chao, Y.C.; Kao, S.H.; Yuan, A.; Lin, C.W.; Yang, S.C.; Chan, W.K.; et al. p53 controls cancer cell invasion by inducing the MDM2-mediated degradation of Slug. Nat. Cell Biol. 2009, 11, 694–704, Erratum in Nat. Cell Biol. 2009, 11, 914. [Google Scholar] [CrossRef] [PubMed]
- Trillsch, F.; Kuerti, S.; Eulenburg, C.; Burandt, E.; Woelber, L.; Prieske, K.; Eylmann, K.; Oliveira-Ferrer, L.; Milde-Langosch, K.; Mahner, S. E-Cadherin fragments as potential mediators for peritoneal metastasis in advanced epithelial ovarian cancer. Br. J. Cancer 2016, 114, 213–220. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- David, J.M.; Rajasekaran, A.K. Dishonorable discharge: The oncogenic roles of cleaved E-cadherin fragments. Cancer Res. 2012, 72, 2917–2923. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Perez-Oquendo, M.; Gibbons, D.L. Regulation of ZEB1 Function and Molecular Associations in Tumor Progression and Metastasis. Cancers 2022, 14, 1864. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Patients | |
|---|---|---|
| IHC | % | |
| n = 524 | ||
| Age | ||
| ≤60 | 165 | 31.49 |
| >60 | 359 | 68.51 |
| Tumor grade | ||
| G1 | 87 | 16.60 |
| G2 | 342 | 65.27 |
| G3 | 92 | 17.56 |
| unknown | 3 | 0.57 |
| Tumor size | ||
| pT1 | 325 | 62.02 |
| pT2 | 168 | 32.06 |
| pT3 | 3 | 0.57 |
| pT4 | 9 | 1.72 |
| unknown | 19 | 3.63 |
| Lymph nodes | ||
| pN0 | 314 | 59.92 |
| pN1–N3 | 180 | 34.35 |
| pNx | 30 | 5.73 |
| Stage | ||
| I | 224 | 42.75 |
| II | 257 | 49.05 |
| III | 18 | 3.44 |
| IV | 0 | 0 |
| ER | ||
| Negative | 177 | 33.78 |
| Positive | 344 | 65.65 |
| Unknown | 3 | 0.57 |
| PR | ||
| Negative | 183 | 34.92 |
| Positive | 338 | 64.50 |
| Unknown | 3 | 0.57 |
| HER-2 | ||
| Negative | 272 | 51.91 |
| Positive | 36 | 6.87 |
| Unknown | 216 | 51.91 |
| Molecular tumor types | ||
| Triple negative | 37 | 7.06 |
| Other types | 484 | 92.3 |
| Unknown | 3 | 0.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Górnicki, T.; Lambrinow, J.; Mrozowska, M.; Krawczyńska, K.; Staszko, N.; Kmiecik, A.; Piotrowska, A.; Gomułkiewicz, A.; Romanowicz, H.; Smolarz, B.; et al. Impact of RBMS 3 Progression on Expression of EMT Markers. Cells 2024, 13, 1548. https://doi.org/10.3390/cells13181548
Górnicki T, Lambrinow J, Mrozowska M, Krawczyńska K, Staszko N, Kmiecik A, Piotrowska A, Gomułkiewicz A, Romanowicz H, Smolarz B, et al. Impact of RBMS 3 Progression on Expression of EMT Markers. Cells. 2024; 13(18):1548. https://doi.org/10.3390/cells13181548
Chicago/Turabian StyleGórnicki, Tomasz, Jakub Lambrinow, Monika Mrozowska, Klaudia Krawczyńska, Natalia Staszko, Alicja Kmiecik, Aleksandra Piotrowska, Agnieszka Gomułkiewicz, Hanna Romanowicz, Beata Smolarz, and et al. 2024. "Impact of RBMS 3 Progression on Expression of EMT Markers" Cells 13, no. 18: 1548. https://doi.org/10.3390/cells13181548