
Microglia-Derived Insulin-like Growth Factor 1 Is Critical for Neurodevelopment

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Behavioral Tests
2.3. Open Field
2.4. Novel Object Recognition (Nor)
2.5. Marble Burying
2.6. qRT-PCR
2.7. Immunohistochemistry
2.8. Stereology
2.9. Electron Microscopy
2.10. Magnetic-Activated Cell Sorting (MACS)
2.11. Statistical Analysis
3. Results
3.1. Postnatal Deficiency of Microglia-Derived Igf1 Leads to Growth Alterations
3.2. Postnatal Deficiency of Microglia-Derived Igf1 Leads to Increased Anxiety and Neophobia
3.3. Postnatal Deficiency of Microglia-Derived Igf1 Leads to Increased Numbers of Microglia
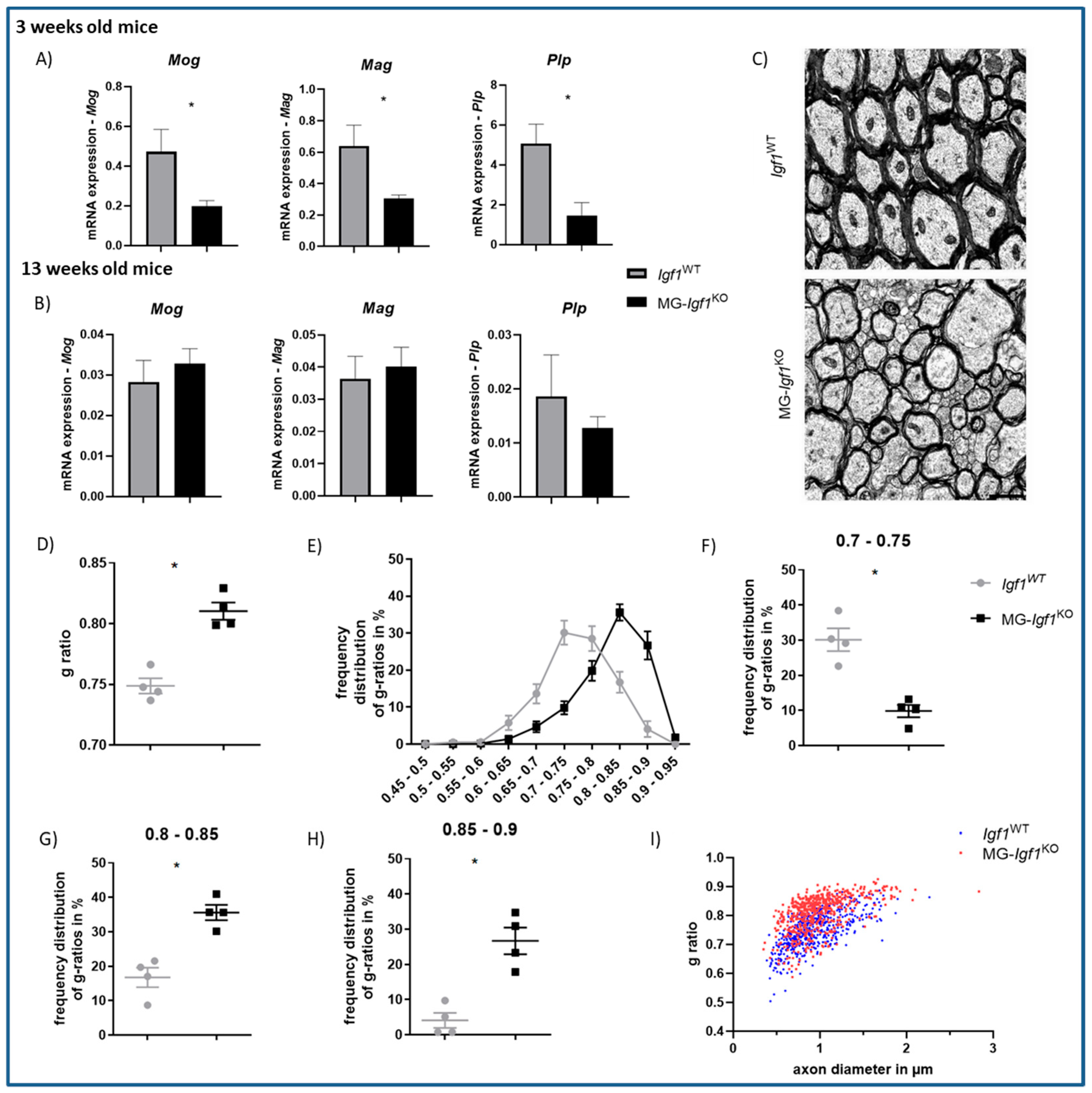
3.4. Postnatal Deficiency of Microglia-Derived Igf1 Leads to Pronounced Myelination Deficits
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benmamar-Badel, A.; Owens, T.; Wlodarczyk, A. Protective Microglial Subset in Development, Aging, and Disease: Lessons From Transcriptomic Studies. Front. Immunol. 2020, 11, 430. [Google Scholar] [CrossRef]
- Ginhoux, F.; Greter, M.; Leboeuf, M.; Nandi, S.; See, P.; Gokhan, S.; Mehler, M.F.; Conway, S.J.; Ng, L.G.; Stanley, E.R.; et al. Fate mapping analysis reveals that adult microglia derive from primitive macrophages. Science 2010, 330, 841–845. [Google Scholar] [CrossRef]
- Ginhoux, F.; Prinz, M. Origin of microglia: Current concepts and past controversies. Cold Spring Harb. Perspect. Biol. 2015, 7, a020537. [Google Scholar] [CrossRef]
- Elmore, M.R.; Najafi, A.R.; Koike, M.A.; Dagher, N.N.; Spangenberg, E.E.; Rice, R.A.; Kitazawa, M.; Matusow, B.; Nguyen, H.; West, B.L.; et al. Colony-stimulating factor 1 receptor signaling is necessary for microglia viability, unmasking a microglia progenitor cell in the adult brain. Neuron 2014, 82, 380–397. [Google Scholar] [CrossRef]
- Paolicelli, R.C.; Bolasco, G.; Pagani, F.; Maggi, L.; Scianni, M.; Panzanelli, P.; Giustetto, M.; Ferreira, T.A.; Guiducci, E.; Dumas, L.; et al. Synaptic pruning by microglia is necessary for normal brain development. Science 2011, 333, 1456–1458. [Google Scholar] [CrossRef]
- Wlodarczyk, A.; Holtman, I.R.; Krueger, M.; Yogev, N.; Bruttger, J.; Khorooshi, R.; Benmamar-Badel, A.; de Boer-Bergsma, J.J.; Martin, N.A.; Karram, K.; et al. A novel microglial subset plays a key role in myelinogenesis in developing brain. EMBO J. 2017, 36, 3292–3308. [Google Scholar] [CrossRef]
- Kettenmann, H.; Kirchhoff, F.; Verkhratsky, A. Microglia: New roles for the synaptic stripper. Neuron 2013, 77, 10–18. [Google Scholar] [CrossRef]
- Ye, P.; Li, L.; Richards, R.G.; DiAugustine, R.P.; D’Ercole, A.J. Myelination is altered in insulin-like growth factor-I null mutant mice. J. Neurosci. 2002, 22, 6041–6051. [Google Scholar] [CrossRef]
- Ueno, M.; Fujita, Y.; Tanaka, T.; Nakamura, Y.; Kikuta, J.; Ishii, M.; Yamashita, T. Layer V cortical neurons require microglial support for survival during postnatal development. Nat. Neurosci. 2013, 16, 543–551. [Google Scholar] [CrossRef]
- Netchine, I.; Azzi, S.; Le Bouc, Y.; Savage, M.O. IGF1 molecular anomalies demonstrate its critical role in fetal, postnatal growth and brain development. Best Pract. Res. Clin. Endocrinol. Metab. 2011, 25, 181–190. [Google Scholar] [CrossRef]
- Beck, K.D.; Powell-Braxton, L.; Widmer, H.R.; Valverde, J.; Hefti, F. Igf1 gene disruption results in reduced brain size, CNS hypomyelination, and loss of hippocampal granule and striatal parvalbumin-containing neurons. Neuron 1995, 14, 717–730. [Google Scholar] [CrossRef]
- Laron, Z. Insulin-like growth factor 1 (IGF-1): A growth hormone. Mol. Pathol. 2001, 54, 311–316. [Google Scholar] [CrossRef]
- Wrigley, S.; Arafa, D.; Tropea, D. Insulin-Like Growth Factor 1: At the Crossroads of Brain Development and Aging. Front. Cell Neurosci. 2017, 11, 14. [Google Scholar] [CrossRef]
- Yona, S.; Kim, K.W.; Wolf, Y.; Mildner, A.; Varol, D.; Breker, M.; Strauss-Ayali, D.; Viukov, S.; Guilliams, M.; Misharin, A.; et al. Fate mapping reveals origins and dynamics of monocytes and tissue macrophages under homeostasis. Immunity 2013, 38, 79–91. [Google Scholar] [CrossRef]
- Liu, J.L.; Grinberg, A.; Westphal, H.; Sauer, B.; Accili, D.; Karas, M.; LeRoith, D. Insulin-like growth factor-I affects perinatal lethality and postnatal development in a gene dosage-dependent manner: Manipulation using the Cre/loxP system in transgenic mice. Mol. Endocrinol. 1998, 12, 1452–1462. [Google Scholar] [CrossRef]
- Sahasrabuddhe, V.; Ghosh, H.S. Cx3Cr1-Cre induction leads to microglial activation and IFN-1 signaling caused by DNA damage in early postnatal brain. Cell Rep. 2022, 38, 110252. [Google Scholar] [CrossRef]
- Lambertsen, K.L.; Gramsbergen, J.B.; Sivasaravanaparan, M.; Ditzel, N.; Sevelsted-Moller, L.M.; Olivan-Viguera, A.; Rabjerg, M.; Wulff, H.; Kohler, R. Genetic KCa3.1-deficiency produces locomotor hyperactivity and alterations in cerebral monoamine levels. PLoS ONE 2012, 7, e47744. [Google Scholar] [CrossRef]
- Antunes, M.; Biala, G. The novel object recognition memory: Neurobiology, test procedure, and its modifications. Cogn. Process 2012, 13, 93–110. [Google Scholar] [CrossRef]
- Wirenfeldt, M.; Dalmau, I.; Finsen, B. Estimation of absolute microglial cell numbers in mouse fascia dentata using unbiased and efficient stereological cell counting principles. Glia 2003, 44, 129–139. [Google Scholar] [CrossRef]
- Himanshu; Dharmila; Sarkar, D.; Nutan. A Review of Behavioral Tests to Evaluate Different Types of Anxiety and Anti-anxiety Effects. Clin. Psychopharmacol. Neurosci. 2020, 18, 341–351. [Google Scholar] [CrossRef]
- Seibenhener, M.L.; Wooten, M.C. Use of the Open Field Maze to measure locomotor and anxiety-like behavior in mice. J. Vis. Exp. 2015, 96, e52434. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Stewart, A.M.; Song, C.; Berridge, K.C.; Graybiel, A.M.; Fentress, J.C. Neurobiology of rodent self-grooming and its value for translational neuroscience. Nat. Rev. Neurosci. 2016, 17, 45–59. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Tuohimaa, P. Grooming analysis algorithm for neurobehavioural stress research. Brain Res. Brain Res. Protoc. 2004, 13, 151–158. [Google Scholar] [CrossRef]
- Leger, M.; Quiedeville, A.; Bouet, V.; Haelewyn, B.; Boulouard, M.; Schumann-Bard, P.; Freret, T. Object recognition test in mice. Nat. Protoc. 2013, 8, 2531–2537. [Google Scholar] [CrossRef]
- Valvassori, S.S.; Borges, C.P.; Varela, R.B.; Bavaresco, D.V.; Bianchini, G.; Mariot, E.; Arent, C.O.; Resende, W.R.; Budni, J.; Quevedo, J. The different effects of lithium and tamoxifen on memory formation and the levels of neurotrophic factors in the brain of male and female rats. Brain Res. Bull. 2017, 134, 228–235. [Google Scholar] [CrossRef]
- Begolly, S.; Shrager, P.G.; Olschowka, J.A.; Williams, J.P.; O’Banion, M.K. Fractionation Spares Mice From Radiation-Induced Reductions in Weight Gain But Does Not Prevent Late Oligodendrocyte Lineage Side Effects. Int. J. Radiat. Oncol. Biol. Phys. 2016, 96, 449–457. [Google Scholar] [CrossRef][Green Version]
- Al-Samerria, S.; Radovick, S. The Role of Insulin-like Growth Factor-1 (IGF-1) in the Control of Neuroendocrine Regulation of Growth. Cells 2021, 10, 2664. [Google Scholar] [CrossRef]
- Kappeler, L.; De Magalhaes Filho, C.; Dupont, J.; Leneuve, P.; Cervera, P.; Perin, L.; Loudes, C.; Blaise, A.; Klein, R.; Epelbaum, J.; et al. Brain IGF-1 receptors control mammalian growth and lifespan through a neuroendocrine mechanism. PLoS Biol. 2008, 6, e254. [Google Scholar] [CrossRef]
- Ivan, D.C.; Berve, K.C.; Walthert, S.; Monaco, G.; Borst, K.; Bouillet, E.; Ferreira, F.; Lee, H.; Steudler, J.; Buch, T.; et al. Insulin-like growth factor-1 receptor controls the function of CNS-resident macrophages and their contribution to neuroinflammation. Acta Neuropathol. Commun. 2023, 11, 35. [Google Scholar] [CrossRef]
- Carson, M.J.; Behringer, R.R.; Brinster, R.L.; McMorris, F.A. Insulin-like growth factor I increases brain growth and central nervous system myelination in transgenic mice. Neuron 1993, 10, 729–740. [Google Scholar] [CrossRef]
- Vanhala, R.; Turpeinen, U.; Riikonen, R. Low levels of insulin-like growth factor-I in cerebrospinal fluid in children with autism. Dev. Med. Child. Neurol. 2001, 43, 614–616. [Google Scholar] [CrossRef]
- Riikonen, R.; Makkonen, I.; Vanhala, R.; Turpeinen, U.; Kuikka, J.; Kokki, H. Cerebrospinal fluid insulin-like growth factors IGF-1 and IGF-2 in infantile autism. Dev. Med. Child. Neurol. 2006, 48, 751–755. [Google Scholar] [CrossRef]
- Mu, M.D.; Geng, H.Y.; Rong, K.L.; Peng, R.C.; Wang, S.T.; Geng, L.T.; Qian, Z.M.; Yung, W.H.; Ke, Y. A limbic circuitry involved in emotional stress-induced grooming. Nat. Commun. 2020, 11, 2261. [Google Scholar] [CrossRef]
- Hu, C.; Li, H.; Li, J.; Luo, X.; Hao, Y. Microglia: Synaptic modulator in autism spectrum disorder. Front. Psychiatry 2022, 13, 958661. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Target | Assay ID |
|---|---|---|
| Igf1 | Insulin-like growth factor 1 | Mm00439560_m1 |
| Mog | Myelin oligodendrocyte glycoprotein | Mm00447824_m1 |
| Mag | Myelin-associated glycoprotein | Mm00487538_m1 |
| Plp | Myelin proteolipid protein | Mm01297210_m1 |
| Gfap | Glial fibrillary acidic protein | Mm01253033_m1 |
| Sall1 | Sall-like Protein 1 | Mm00491266_m1 |
| Aif1 | Allograft inflammatory factor 1 | Mm00479862_g1 |
| Trem2 | Triggering receptor expressed on myeloid cells 2 | Mm04209424_g1 |
| 18S | 18S ribosomal RNA | 00853255 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rusin, D.; Vahl Becirovic, L.; Lyszczarz, G.; Krueger, M.; Benmamar-Badel, A.; Vad Mathiesen, C.; Sigurðardóttir Schiöth, E.; Lykke Lambertsen, K.; Wlodarczyk, A. Microglia-Derived Insulin-like Growth Factor 1 Is Critical for Neurodevelopment. Cells 2024, 13, 184. https://doi.org/10.3390/cells13020184
Rusin D, Vahl Becirovic L, Lyszczarz G, Krueger M, Benmamar-Badel A, Vad Mathiesen C, Sigurðardóttir Schiöth E, Lykke Lambertsen K, Wlodarczyk A. Microglia-Derived Insulin-like Growth Factor 1 Is Critical for Neurodevelopment. Cells. 2024; 13(2):184. https://doi.org/10.3390/cells13020184
Chicago/Turabian StyleRusin, Dominika, Lejla Vahl Becirovic, Gabriela Lyszczarz, Martin Krueger, Anouk Benmamar-Badel, Cecilie Vad Mathiesen, Eydís Sigurðardóttir Schiöth, Kate Lykke Lambertsen, and Agnieszka Wlodarczyk. 2024. "Microglia-Derived Insulin-like Growth Factor 1 Is Critical for Neurodevelopment" Cells 13, no. 2: 184. https://doi.org/10.3390/cells13020184
APA StyleRusin, D., Vahl Becirovic, L., Lyszczarz, G., Krueger, M., Benmamar-Badel, A., Vad Mathiesen, C., Sigurðardóttir Schiöth, E., Lykke Lambertsen, K., & Wlodarczyk, A. (2024). Microglia-Derived Insulin-like Growth Factor 1 Is Critical for Neurodevelopment. Cells, 13(2), 184. https://doi.org/10.3390/cells13020184