
Fish Cell Spheroids, a Promising In Vitro Model to Mimic In Vivo Research: A Review



Abstract
:
1. Introduction
2. Conventional Spheroid Generation Methods
2.1. Hanging Drop
2.2. Low-Adherence Substrates
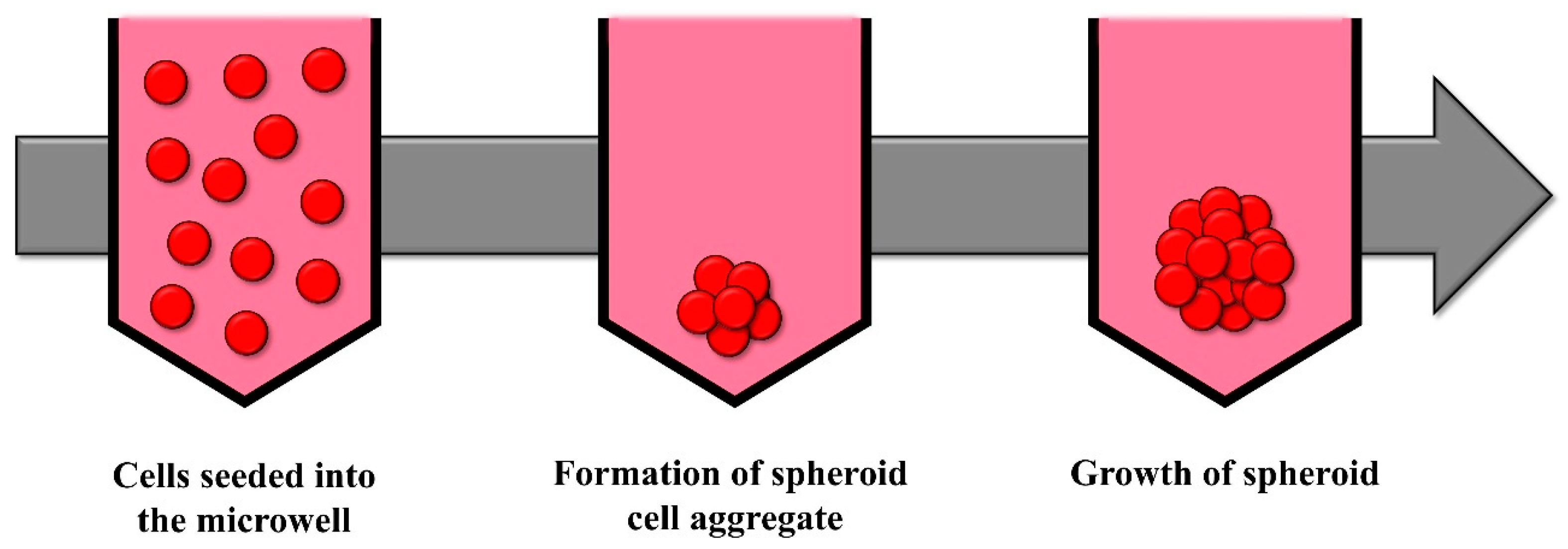
2.3. Wells and Microwells in Ultra-Low Attachment Plates
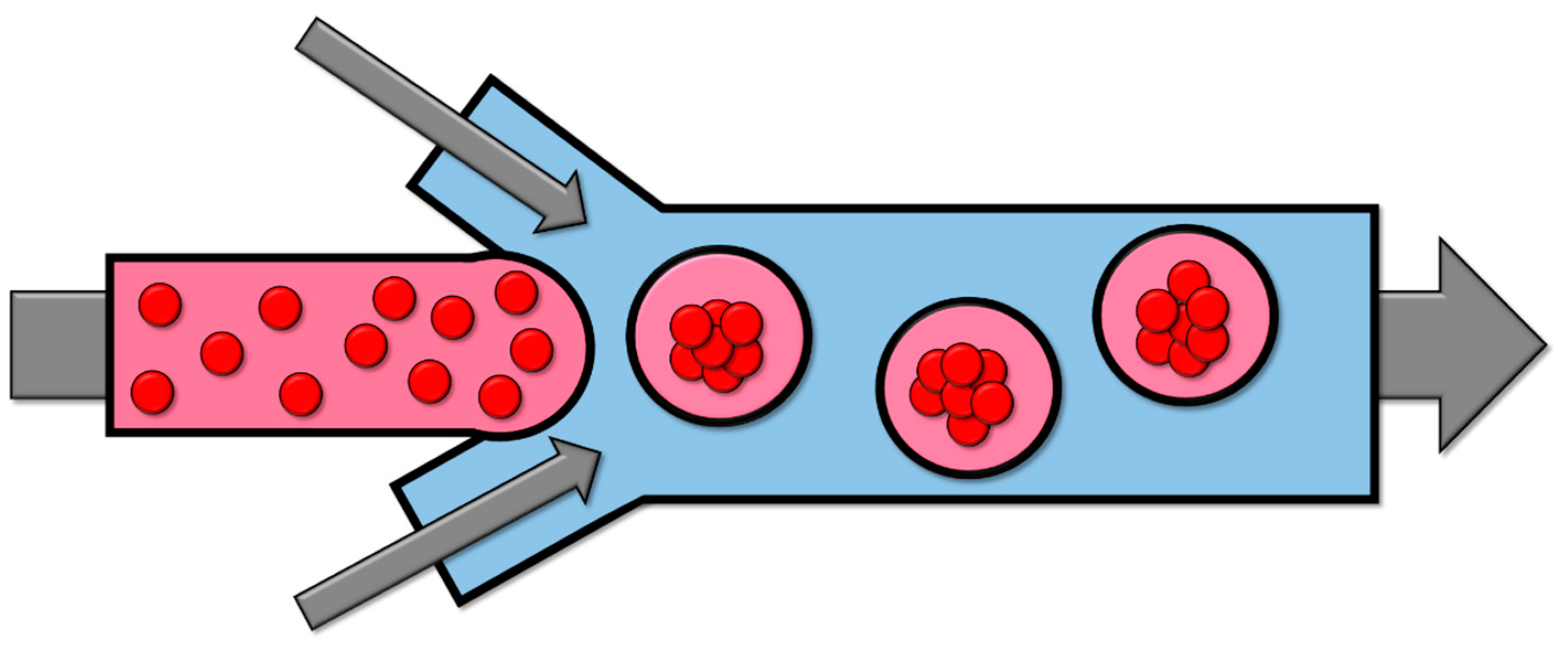
2.4. Microfluidics
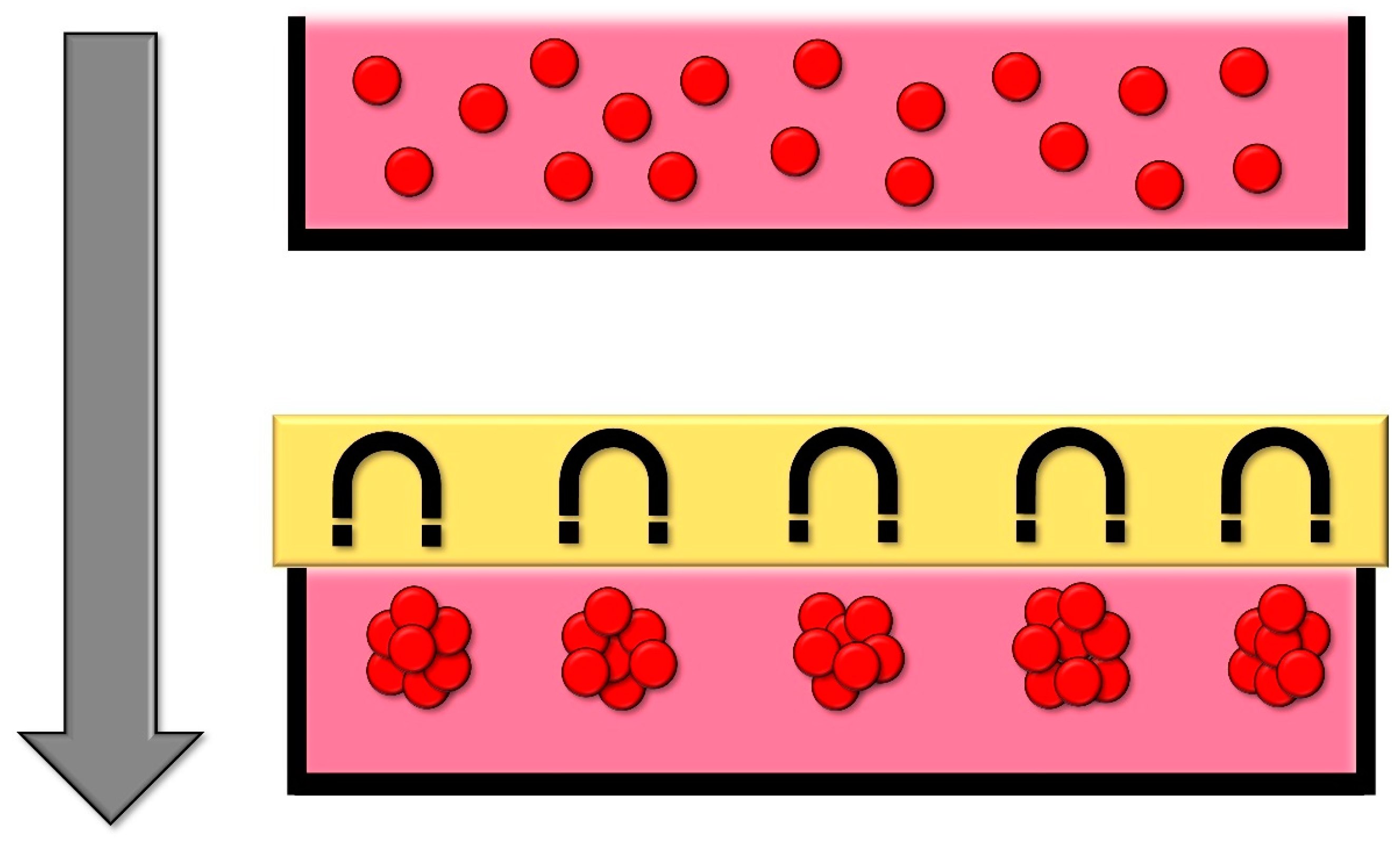
2.5. Magnetic Levitation
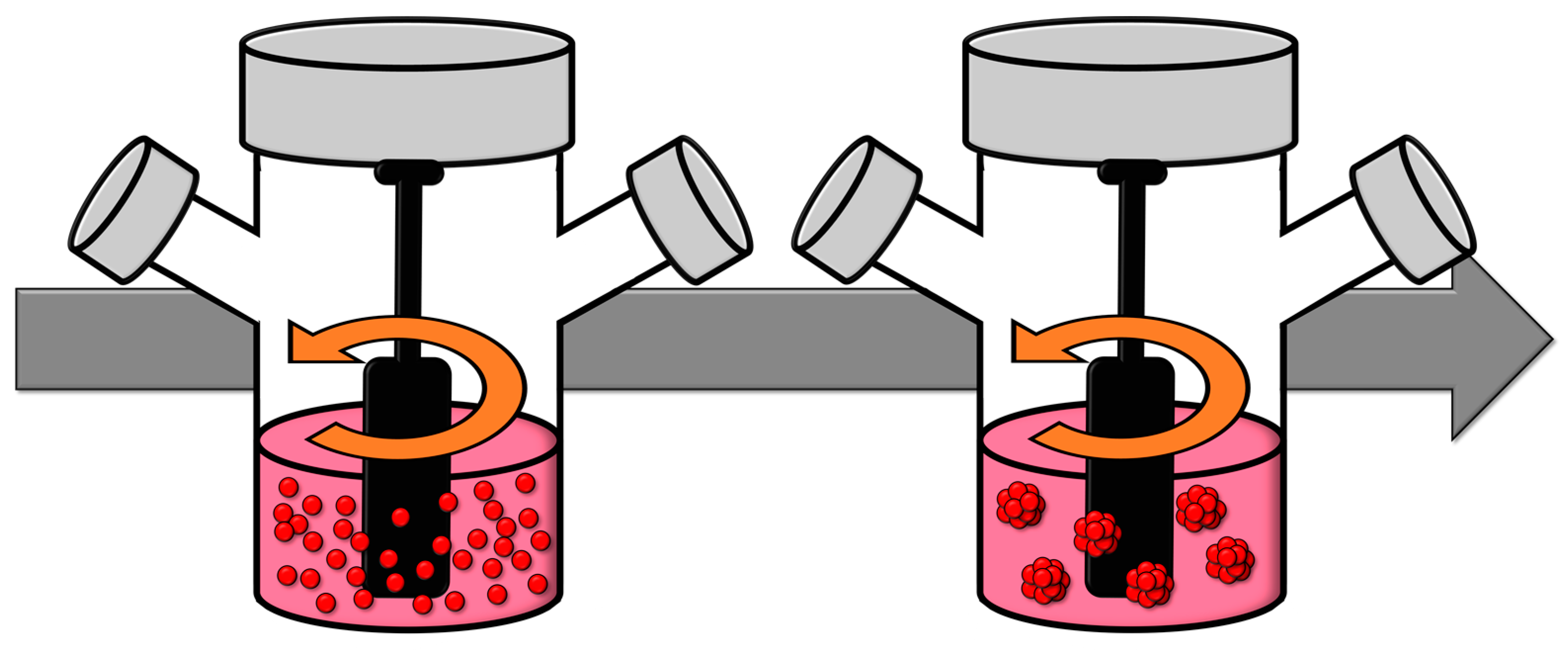
2.6. Spinner Flasks
3. Fish Cell Spheroid Applications
3.1. Characterization and Optimization of Spheroid Culture
3.2. Fish Pathology
3.3. Metabolism
3.4. Toxicology
3.5. Endocrinology
4. Future Prospects
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Nash, C. The History of Aquaculture; John Wiley & Sons: Hoboken, NJ, USA, 2010. [Google Scholar]
- Naylor, R.L.; Kishore, A.; Sumaila, U.R.; Issifu, I.; Hunter, B.P.; Belton, B.; Bush, S.R.; Cao, L.; Gelcich, S.; Gephart, J.A. Blue Food Demand across Geographic and Temporal Scales. Nat. Commun. 2021, 12, 5413. [Google Scholar] [CrossRef] [PubMed]
- FAO. The State of World Fisheries and Aquaculture. towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar]
- Chen, W.; Gao, S. Current Status of Industrialized Aquaculture in China: A Review. Environ. Sci. Pollut. Res. 2023, 30, 32278–32287. [Google Scholar] [CrossRef]
- Santoro, M.M. Zebrafish as a Model to Explore Cell Metabolism. Trends Endocrinol. Metab. 2014, 25, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, H.; Shimada, Y.; Zang, L.; Terasawa, M.; Nishiura, K.; Matsuda, K.; Toombs, C.; Langdon, C.; Nishimura, N. Novel Anti-Obesity Properties of Palmaria mollis in Zebrafish and Mouse Models. Nutrients 2018, 10, 1401. [Google Scholar] [CrossRef] [PubMed]
- de Souza, I.R.; Canavez, A.D.P.M.; Schuck, D.C.; Gagosian, V.S.C.; de Souza, I.R.; de Albuquerque Vita, N.; da Silva Trindade, E.; Cestari, M.M.; Lorencini, M.; Leme, D.M. A 3D Culture Method of Spheroids of Embryonic and Liver Zebrafish Cell Lines. J. Vis. Exp. 2023, 191, e64859. [Google Scholar] [CrossRef]
- Bols, N.C. Biotechnology and Aquaculture: The Role of Cell Cultures. Biotechnol. Adv. 1991, 9, 31–49. [Google Scholar] [CrossRef]
- Russell, W.M.S.; Burch, R.L.; Hume, C.W. The Principles of Humane Experimental Technique; Methuen & Co Ltd.: London, UK, 1959. [Google Scholar]
- de Souza, I.R.; Canavez, A.D.P.M.; Schuck, D.C.; Gagosian, V.S.C.; de Souza, I.R.; Vicari, T.; da Silva Trindade, E.; Cestari, M.M.; Lorencini, M.; Leme, D.M. Development of 3D Cultures of Zebrafish Liver and Embryo Cell Lines: A Comparison of Different Spheroid Formation Methods. Ecotoxicology 2021, 30, 1893–1909. [Google Scholar] [CrossRef]
- Goswami, M.; Yashwanth, B.S.; Trudeau, V.; Lakra, W.S. Role and Relevance of Fish Cell Lines in Advanced in Vitro Research. Mol. Biol. Rep. 2022, 49, 1–19. [Google Scholar] [CrossRef]
- Ruiz-Palacios, M.; Almeida, M.; Martins, M.A.; Oliveira, M.; Esteban, M.Á.; Cuesta, A. Establishment of a Brain Cell Line (FuB-1) from Mummichog (Fundulus heteroclitus) and Its Application to Fish Virology, Immunity and Nanoplastics Toxicology. Sci. Total Environ. 2020, 708, 134821. [Google Scholar] [CrossRef]
- Suryakodi, S.; Majeed, S.A.; Taju, G.; Vimal, S.; Sivakumar, S.; Ahmed, A.N.; Shah, F.A.; Bhat, S.A.; Sarma, D.; Begum, A. Development and Characterization of Novel Cell Lines from Kidney and Eye of Rainbow Trout, Oncorhynchus mykiss for Virological Studies. Aquaculture 2021, 532, 736027. [Google Scholar] [CrossRef]
- Li, Y.; Jia, P.; Yu, F.; Li, W.; Mao, C.; Yi, M.; Gu, Q.; Jia, K. Establishment and Characterization of a Liver Cell Line, ALL, Derived from Yellowfin Sea Bream, Acanthopagrus latus, and Its Application to Fish Virology. J. Fish. Dis. 2022, 45, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Ali, F.; Lazar, R.; Haffner, D.; Adeli, K. Development of a Rapid and Simple Genotoxicity Assay Using a Brown Bullhead Fish Cell-Line: Application to Toxicological Surveys of Sediments in the Huron-Erie Corridor. J. Great Lakes Res. 1993, 19, 342–351. [Google Scholar] [CrossRef]
- Shúilleabháin, S.N.; Mothersill, C.; Sheehan, D.; O’Brien, N.M.; O’Halloran, J.; Van Pelt, F.; Davoren, M. In Vitro Cytotoxicity Testing of Three Zinc Metal Salts Using Established Fish Cell Lines. Toxicol. Vitr. 2004, 18, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Kolarova, J.; Velisek, J.; Svobodova, Z. Comparison of in Vitro (Fish Cell Line) and in Vivo (Fish and Crustacean) Acute Toxicity Tests in Aquatic Toxicology. Vet. Med. 2021, 66, 350–355. [Google Scholar] [CrossRef]
- Collet, B.; Collins, C.; Lester, K. Engineered Cell Lines for Fish Health Research. Dev. Comp. Immunol. 2018, 80, 34–40. [Google Scholar] [CrossRef]
- Corripio-Miyar, Y.; Secombes, C.J.; Zou, J. Long-Term Stimulation of Trout Head Kidney Cells with the Cytokines MCSF, IL-2 and IL-6: Gene Expression Dynamics. Fish Shellfish Immunol. 2012, 32, 35–44. [Google Scholar] [CrossRef]
- Villalba, M.; Pérez, V.; Herrera, L.; Stepke, C.; Maldonado, N.; Fredericksen, F.; Yáñez, A.; Olavarría, V.H. Infectious Pancreatic Necrosis Virus Infection of Fish Cell Lines: Preliminary Analysis of Gene Expressions Related to Extracellular Matrix Remodeling and Immunity. Vet. Immunol. Immunopathol. 2017, 193–194, 10–17. [Google Scholar] [CrossRef]
- Fierro-Castro, C.; Barrioluengo, L.; López-Fierro, P.; Razquin, B.E.; Carracedo, B.; Villena, A.J. Fish Cell Cultures as in Vitro Models of Pro-Inflammatory Responses Elicited by Immunostimulants. Fish Shellfish Immunol. 2012, 33, 389–400. [Google Scholar] [CrossRef]
- Brubacher, J.L.; Secombes, C.J.; Zou, J.; Bols, N.C. Constitutive and LPS-Induced Gene Expression in a Macrophage-like Cell Line from the Rainbow Trout (Oncorhynchus mykiss). Dev. Comp. Immunol. 2000, 24, 565–574. [Google Scholar] [CrossRef]
- Nagpure, N.S.; Kumar, A.; Dubey, A.; Mishra, A.K.; Kumar, R.; Goswami, M. Establishment of National Repository of Fish Cell Lines (NRFC) at NBFGR, Lucknow. Available online: https://www.researchgate.net/publication/260436225 (accessed on 10 September 2024).
- Griffith, L.G.; Swartz, M.A. Capturing Complex 3D Tissue Physiology in Vitro. Nat. Rev. Mol. Cell Biol. 2006, 7, 211–224. [Google Scholar] [CrossRef]
- Breslin, S.; O’Driscoll, L. Three-Dimensional Cell Culture: The Missing Link in Drug Discovery. Drug Discov. Today 2013, 18, 240–249. [Google Scholar] [CrossRef]
- Jensen, C.; Teng, Y. Is It Time to Start Transitioning from 2D to 3D Cell Culture? Front. Mol. Biosci. 2020, 7, 33. [Google Scholar] [CrossRef]
- Habanjar, O.; Diab-Assaf, M.; Caldefie-Chezet, F.; Delort, L. 3D Cell Culture Systems: Tumor Application, Advantages, and Disadvantages. Int. J. Mol. Sci. 2021, 22, 12200. [Google Scholar] [CrossRef]
- Pereira, I.L.; Lopes, C.; Rocha, E.; Madureira, T. V Establishing Brown Trout Primary Hepatocyte Spheroids as a New Alternative Experimental Model—Testing the Effects of 5α-Dihydrotestosterone on Lipid Pathways. Aquat. Toxicol. 2022, 253, 106331. [Google Scholar] [CrossRef]
- Součková, K.; Jasík, M.; Sovadinová, I.; Sember, A.; Sychrová, E.; Konieczna, A.; Bystrý, V.; Dyková, I.; Blažek, R.; Lukšíková, K.; et al. From Fish to Cells: Establishment of Continuous Cell Lines from Embryos of Annual Killifish Nothobranchius Furzeri and N. Kadleci. Aquat. Toxicol. 2023, 259, 106517. [Google Scholar] [CrossRef]
- Wang, T.; Desmet, J.; Pérez-Albaladejo, E.; Porte, C. Development of Fish Liver PLHC-1 Spheroids and Its Applicability to Investigate the Toxicity of Plastic Additives. Ecotoxicol. Environ. Saf. 2023, 259, 115016. [Google Scholar] [CrossRef]
- Rodd, A.L.; Messier, N.J.; Vaslet, C.A.; Kane, A.B. A 3D Fish Liver Model for Aquatic Toxicology: Morphological Changes and Cyp1a Induction in PLHC-1 Microtissues after Repeated Benzo (a) Pyrene Exposures. Aquat. Toxicol. 2017, 186, 134–144. [Google Scholar] [CrossRef]
- Pommerenke, C.; Nagel, S.; Haake, J.; Koelz, A.L.; Christgen, M.; Steenpass, L.; Eberth, S. Molecular Characterization and Subtyping of Breast Cancer Cell Lines Provide Novel Insights into Cancer Relevant Genes. Cells 2024, 13, 301. [Google Scholar] [CrossRef]
- Ben-David, U.; Siranosian, B.; Ha, G.; Tang, H.; Oren, Y.; Hinohara, K.; Strathdee, C.A.; Dempster, J.; Lyons, N.J.; Burns, R.; et al. Genetic and Transcriptional Evolution Alters Cancer Cell Line Drug Response. Nature 2018, 560, 325–330. [Google Scholar] [CrossRef]
- Yamada, K.M.; Collins, J.W.; Cruz Walma, D.A.; Doyle, A.D.; Morales, S.G.; Lu, J.; Matsumoto, K.; Nazari, S.S.; Sekiguchi, R.; Shinsato, Y. Extracellular Matrix Dynamics in Cell Migration, Invasion and Tissue Morphogenesis. Int. J. Exp. Pathol. 2019, 100, 144–152. [Google Scholar] [CrossRef]
- Wang, F.; Weaver, V.M.; Petersen, O.W.; Larabell, C.A.; Dedhar, S.; Briand, P.; Lupu, R.; Bissell, M.J. Reciprocal Interactions between Β1-Integrin and Epidermal Growth Factor Receptor in Three-Dimensional Basement Membrane Breast Cultures: A Different Perspective in Epithelial Biology. Proc. Natl. Acad. Sci. USA 1998, 95, 14821–14826. [Google Scholar] [CrossRef]
- Whitesides, G.M. The Origins and the Future of Microfluidics. Nature 2006, 442, 368–373. [Google Scholar] [CrossRef]
- Liu, D.; Chen, S.; Win Naing, M. A Review of Manufacturing Capabilities of Cell Spheroid Generation Technologies and Future Development. Biotechnol. Bioeng. 2021, 118, 542–554. [Google Scholar] [CrossRef]
- Haycock, J.W. 3D Cell Culture: A Review of Current Approaches and Techniques. In 3D Cell Culture: Methods in Molecular Biology; Haycock, J., Ed.; Humana Press: Totowa, NJ, USA, 2010; Volume 695, pp. 1–15. [Google Scholar]
- Meseguer, J.; Esteban Abad, M.; de los, Á.; Mulero Méndez, V.F.; Cuesta Peñafiel, A.; Sepulcre Cortés, M.P. Esferoides y Esferas Líquidas. Cultivos Celulares En 3D Para Mimetizar El Ambiente de Las Células En El Organismo. Eubacteria 2015, 2015, 15–20. [Google Scholar]
- Kim, Y.; Rajagopalan, P. 3D Hepatic Cultures Simultaneously Maintain Primary Hepatocyte and Liver Sinusoidal Endothelial Cell Phenotypes. PLoS ONE 2010, 5, e15456. [Google Scholar] [CrossRef]
- Bates, R.C.; Edwards, N.S.; Yates, J.D. Spheroids and Cell Survival. Crit. Rev. Oncol. Hematol. 2000, 36, 61–74. [Google Scholar] [CrossRef]
- Subramanian, K.; Owens, D.J.; Raju, R.; Firpo, M.; O’Brien, T.D.; Verfaillie, C.M.; Hu, W.-S. Spheroid Culture for Enhanced Differentiation of Human Embryonic Stem Cells to Hepatocyte-like Cells. Stem Cells Dev. 2014, 23, 124–131. [Google Scholar] [CrossRef]
- Cui, X.; Hartanto, Y.; Zhang, H. Advances in Multicellular Spheroids Formation. J. R. Soc. Interface 2017, 14, 20160877. [Google Scholar] [CrossRef]
- Alves, R.F.; Lopes, C.; Rocha, E.; Madureira, T. V A Step Forward in the Characterization of Primary Brown Trout Hepatocytic Spheroids as Experimental Models. Animals 2023, 13, 2277. [Google Scholar] [CrossRef]
- Moldaschl, J.; Chariyev-Prinz, F.; Toegel, S.; Keck, M.; Hiden, U.; Egger, D.; Kasper, C. Spheroid Trilineage Differentiation Model of Primary Mesenchymal Stem/Stromal Cells under Hypoxia and Serum-Free Culture Conditions. Front. Bioeng. Biotechnol. 2024, 12, 1444363. [Google Scholar] [CrossRef]
- Gheytanchi, E.; Naseri, M.; Karimi-Busheri, F.; Atyabi, F.; Mirsharif, E.S.; Bozorgmehr, M.; Ghods, R.; Madjd, Z. Morphological and Molecular Characteristics of Spheroid Formation in HT-29 and Caco-2 Colorectal Cancer Cell Lines. Cancer Cell Int. 2021, 21, 204. [Google Scholar] [CrossRef]
- Mirabelli, P.; Coppola, L.; Salvatore, M. Cancer Cell Lines Are Useful Model Systems for Medical Research. Cancers 2019, 11, 1098. [Google Scholar] [CrossRef]
- Hendriks, D.F.G.; Hurrell, T.; Riede, J.; van der Horst, M.; Tuovinen, S.; Ingelman-Sundberg, M. Mechanisms of Chronic Fialuridine Hepatotoxicity as Revealed in Primary Human Hepatocyte Spheroids. Toxicol. Sci. 2019, 171, 385–395. [Google Scholar] [CrossRef]
- Yun, C.; Kim, S.H.; Kim, K.M.; Yang, M.H.; Byun, M.R.; Kim, J.-H.; Kwon, D.; Pham, H.T.M.; Kim, H.-S.; Kim, J.-H. Advantages of Using 3D Spheroid Culture Systems in Toxicological and Pharmacological Assessment for Osteogenesis Research. Int. J. Mol. Sci. 2024, 25, 2512. [Google Scholar] [CrossRef]
- Fey, S.J.; Wrzesinski, K. Determination of Drug Toxicity Using 3D Spheroids Constructed from an Immortal Human Hepatocyte Cell Line. Toxicol. Sci. 2012, 127, 403–411. [Google Scholar] [CrossRef]
- Takayama, K.; Kawabata, K.; Nagamoto, Y.; Kishimoto, K.; Tashiro, K.; Sakurai, F.; Tachibana, M.; Kanda, K.; Hayakawa, T.; Furue, M.K. 3D Spheroid Culture of HESC/HiPSC-Derived Hepatocyte-like Cells for Drug Toxicity Testing. Biomaterials 2013, 34, 1781–1789. [Google Scholar] [CrossRef]
- Ding, H.; Jambunathan, K.; Jiang, G.; Margolis, D.M.; Leng, I.; Ihnat, M.; Ma, J.-X.; Mirsalis, J.; Zhang, Y. 3D Spheroids of Human Primary Urine-Derived Stem Cells in the Assessment of Drug-Induced Mitochondrial Toxicity. Pharmaceutics 2022, 14, 1042. [Google Scholar] [CrossRef]
- Mukundan, S.; Singh, P.; Shah, A.; Kumar, R.; O’Neill, K.C.; Carter, C.L.; Russell, D.G.; Subbian, S.; Parekkadan, B. In Vitro Miniaturized Tuberculosis Spheroid Model. Biomedicines 2021, 9, 1209. [Google Scholar] [CrossRef]
- Nath, S.; Devi, G.R. Three-Dimensional Culture Systems in Cancer Research: Focus on Tumor Spheroid Model. Pharmacol. Ther. 2016, 163, 94–108. [Google Scholar] [CrossRef]
- Nunes, A.S.; Barros, A.S.; Costa, E.C.; Moreira, A.F.; Correia, I.J. 3D Tumor Spheroids as in Vitro Models to Mimic in Vivo Human Solid Tumors Resistance to Therapeutic Drugs. Biotechnol. Bioeng. 2019, 116, 206–226. [Google Scholar] [CrossRef]
- Gilazieva, Z.; Ponomarev, A.; Rutland, C.; Rizvanov, A.; Solovyeva, V. Promising Applications of Tumor Spheroids and Organoids for Personalized Medicine. Cancers 2020, 12, 2727. [Google Scholar] [CrossRef] [PubMed]
- Weiswald, L.-B.; Bellet, D.; Dangles-Marie, V. Spherical Cancer Models in Tumor Biology. Neoplasia 2015, 17, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Herter, S.; Morra, L.; Schlenker, R.; Sulcova, J.; Fahrni, L.; Waldhauer, I.; Lehmann, S.; Reisländer, T.; Agarkova, I.; Kelm, J.M. A Novel Three-Dimensional Heterotypic Spheroid Model for the Assessment of the Activity of Cancer Immunotherapy Agents. Cancer Immunol. Immunother. 2017, 66, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.-Y.; Chang, Y.-H.; Yeh, Y.-C.; Chen, C.-H.; Lin, K.M.; Huang, C.-C.; Chang, Y.; Sung, H.-W. The Use of Injectable Spherically Symmetric Cell Aggregates Self-Assembled in a Thermo-Responsive Hydrogel for Enhanced Cell Transplantation. Biomaterials 2009, 30, 5505–5513. [Google Scholar] [CrossRef]
- Liu, J.; Abate, W.; Xu, J.; Corry, D.; Kaul, B.; Jackson, S.K. Three-Dimensional Spheroid Cultures of A549 and HepG2 Cells Exhibit Different Lipopolysaccharide (LPS) Receptor Expression and LPS-Induced Cytokine Response Compared with Monolayer Cultures. Innate Immun. 2011, 17, 245–255. [Google Scholar]
- Rodrigues, A.V.; Alexandre-Pires, G.; Valério-Bolas, A.; Santos-Mateus, D.; Rafael-Fernandes, M.; Pereira, M.A.; Ligeiro, D.; Nunes, T.; Alves-Azevedo, R.; Santos, M. 3D-Hepatocyte Culture Applied to Parasitology: Immune Activation of Canine Hepatic Spheroids Exposed to Leishmania infantum. Biomedicines 2020, 8, 628. [Google Scholar] [CrossRef]
- O’Rourke, F.; Mändle, T.; Urbich, C.; Dimmeler, S.; Michaelis, U.R.; Brandes, R.P.; Flötenmeyer, M.; Döring, C.; Hansmann, M.; Lauber, K. Reprogramming of Myeloid Angiogenic Cells by B Artonella henselae Leads to Microenvironmental Regulation of Pathological Angiogenesis. Cell. Microbiol. 2015, 17, 1447–1463. [Google Scholar] [CrossRef]
- Steenblock, C.; Fliedner, S.; Spinas, G.A.; Ofir, R.; Kugelmeier, P.; Ludwig, B.; Bechmann, N. Development of Adrenal 3-Dimensional Spheroid Cultures: Potential for the Treatment of Adrenal Insufficiency and Neurodegenerative Diseases. Explor. Endocr. Metab. Dis. 2024, 1, 27–38. [Google Scholar] [CrossRef]
- Park, H.; Kim, J.; Ryou, C. A Three-dimensional Spheroid Co-culture System of Neurons and Astrocytes Derived from Alzheimer’s Disease Patients for Drug Efficacy Testing. Cell Prolif. 2023, 56, e13399. [Google Scholar] [CrossRef]
- Alves, R.F.; Rocha, E.; Madureira, T. V Fish Hepatocyte Spheroids–A Powerful (Though Underexplored) Alternative in Vitro Model to Study Hepatotoxicity. Comp. Biochem. Physiol. Part. C Toxicol. Pharmacol. 2022, 262, 109470. [Google Scholar] [CrossRef]
- Faber, M.N.; Sojan, J.M.; Saraiva, M.; van West, P.; Secombes, C.J. Development of a 3D Spheroid Cell Culture System from Fish Cell Lines for in Vitro Infection Studies: Evaluation with Saprolegnia parasitica. J. Fish. Dis. 2021, 44, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Hultman, M.T.; Løken, K.B.; Grung, M.; Reid, M.J.; Lillicrap, A. Performance of Three-dimensional Rainbow Trout (Oncorhynchus Mykiss) Hepatocyte Spheroids for Evaluating Biotransformation of Pyrene. Environ. Toxicol. Chem. 2019, 38, 1738–1747. [Google Scholar] [CrossRef] [PubMed]
- Langan, L.M.; Dodd, N.J.F.; Owen, S.F.; Purcell, W.M.; Jackson, S.K.; Jha, A.N. Direct Measurements of Oxygen Gradients in Spheroid Culture System Using Electron Parametric Resonance Oximetry. PLoS ONE 2016, 11, e0149492. [Google Scholar]
- Langan, L.M.; Owen, S.F.; Trznadel, M.; Dodd, N.J.F.; Jackson, S.K.; Purcell, W.M.; Jha, A.N. Spheroid Size Does Not Impact Metabolism of the β-Blocker Propranolol in 3D Intestinal Fish Model. Front. Pharmacol. 2018, 9, 947. [Google Scholar] [CrossRef] [PubMed]
- Suryakodi, S.; Mithra, S.; Nafeez Ahmed, A.; Kanimozhi, K.; Rajkumar, V.; Taju, G.; Abdul Majeed, S.; Sahul Hameed, A.S. Development of Three-Dimensional (3D) Spheroid Culture System from Rainbow Trout Kidney Cell Line (RTK) for in Vitro Production of Fish Viral Pathogen. Aquac. Int. 2024, 32, 5029–5044. [Google Scholar] [CrossRef]
- Järvinen, P.; Kakko, M.; Sikanen, T. Cytotoxicity of Pharmaceuticals and Their Mixtures toward Scaffold-Free 3D Spheroid Cultures of Rainbow Trout (Oncorhynchus mykiss) Hepatocytes. Eur. J. Pharm. Sci. 2024, 199, 106817. [Google Scholar] [CrossRef]
- Lammel, T.; Tsoukatou, G.; Jellinek, J.; Sturve, J. Development of Three-Dimensional (3D) Spheroid Cultures of the Continuous Rainbow Trout Liver Cell Line RTL-W1. Ecotoxicol. Environ. Saf. 2019, 167, 250–258. [Google Scholar] [CrossRef]
- Baron, M.G.; Mintram, K.S.; Owen, S.F.; Hetheridge, M.J.; Moody, A.J.; Purcell, W.M.; Jackson, S.K.; Jha, A.N. Pharmaceutical Metabolism in Fish: Using a 3-D Hepatic in Vitro Model to Assess Clearance. PLoS ONE 2017, 12, e0168837. [Google Scholar] [CrossRef] [PubMed]
- Cravedi, J.-P.; Paris, A.; Monod, G.; Devaux, A.; Flouriot, G.; Valotaire, Y. Maintenance of Cytochrome P450 Content and Phase I and Phase II Enzyme Activities in Trout Hepatocytes Cultured as Spheroidal Aggregates. Comp. Biochem. Physiol. Part. C Pharmacol. Toxicol. Endocrinol. 1996, 113, 241–246. [Google Scholar] [CrossRef]
- Uchea, C.; Sarda, S.; Schulz-Utermoehl, T.; Owen, S.; Chipman, K.J. In Vitro Models of Xenobiotic Metabolism in Trout for Use in Environmental Bioaccumulation Studies. Xenobiotica 2013, 43, 421–431. [Google Scholar] [CrossRef]
- Uchea, C.; Owen, S.F.; Chipman, J.K. Functional Xenobiotic Metabolism and Efflux Transporters in Trout Hepatocyte Spheroid Cultures. Toxicol. Res. 2015, 4, 494–507. [Google Scholar] [CrossRef] [PubMed]
- Flouriot, G.; Monod, G.; Valotaire, Y.; Devaux, A.; Cravedi, J.-P. Xenobiotic Metabolizing Enzyme Activities in Aggregate Culture of Rainbow Trout Hepatocytes. Mar. Environ. Res. 1995, 39, 293–297. [Google Scholar] [CrossRef]
- Baron, M.G.; Purcell, W.M.; Jackson, S.K.; Owen, S.F.; Jha, A.N. Towards a More Representative in Vitro Method for Fish Ecotoxicology: Morphological and Biochemical Characterisation of Three-Dimensional Spheroidal Hepatocytes. Ecotoxicology 2012, 21, 2419–2429. [Google Scholar] [CrossRef]
- Flouriot, G.; Vaillant, C.; Salbert, G.; Pelissero, C.; Guiraud, J.M.; Valotaire, Y. Monolayer and Aggregate Cultures of Rainbow Trout Hepatocytes: Long-Term and Stable Liver-Specific Expression in Aggregates. J. Cell Sci. 1993, 105, 407–416. [Google Scholar] [CrossRef]
- Sullivan, K.M.; Park, C.G.; Ito, J.D.; Kandel, M.; Popescu, G.; Kim, Y.J.; Kong, H. Park. Matrix Softness-Mediated 3D Zebrafish Hepatocyte Modulates Response to Endocrine Disrupting Chemicals. Environ. Sci. Technol. 2022, 54, 13797–13806. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.; Park, C.B.; Baek, I.H.; Jeong, K.; Baik, S. Differential Effects of CBZ-Induced Catalysis and Cytochrome Gene Expression in Three Dimensional Zebrafish Liver Cellculture. J. Environ. Anal. Toxicol. 2016, 6, 404. [Google Scholar] [CrossRef]
- Park, C.G.; Ryu, C.S.; Sung, B.; Manz, A.; Kong, H.; Kim, Y.J. Transcriptomic and Physiological Analysis of Endocrine Disrupting Chemicals Impacts on 3D Zebrafish Liver Cell Culture System. Aquat. Toxicol. 2022, 245, 106105. [Google Scholar] [CrossRef]
- Biswas, T.; Rajendran, N.; Hassan, H.; Zhao, C.; Rohner, N. 3D Spheroid Culturing of Astyanax Mexicanus Liver-Derived Cell Lines Recapitulates Distinct Transcriptomic and Metabolic States of in Vivo Tissue Environment. bioRxiv 2023, 342, 2006–2023. [Google Scholar] [CrossRef]
- Tsuruwaka, Y.; Shimada, E. Reprocessing Seafood Waste: Challenge to Develop Aquatic Clean Meat from Fish Cells. npj Sci. Food 2022, 6, 7. [Google Scholar] [CrossRef]
- Yamaguchi, A. Evaluation of Fish Pituitary Spheroids to Study Annual Endocrine Reproductive Control. Gen. Comp. Endocrinol. 2024, 351, 114481. [Google Scholar] [CrossRef]
- Wang, T.; Desmet, J.; Porte, C. Protective Role of Fetal Bovine Serum on PLHC-1 Spheroids Exposed to a Mixture of Plastic Additives: A Lipidomic Perspective. Toxicol. Vitr. 2024, 96, 105771. [Google Scholar] [CrossRef] [PubMed]
- Ryu, N.-E.; Lee, S.-H.; Park, H. Spheroid Culture System Methods and Applications for Mesenchymal Stem Cells. Cells 2019, 8, 1620. [Google Scholar] [CrossRef] [PubMed]
- Fennema, E.; Rivron, N.; Rouwkema, J.; van Blitterswijk, C.; De Boer, J. Spheroid Culture as a Tool for Creating 3D Complex Tissues. Trends Biotechnol. 2013, 31, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Damman, R.; Lucini Paioni, A.; Xenaki, K.T.; Beltrán Hernández, I.; van Bergen en Henegouwen, P.M.P.; Baldus, M. Development of in Vitro-Grown Spheroids as a 3D Tumor Model System for Solid-State NMR Spectroscopy. J. Biomol. NMR 2020, 74, 401–412. [Google Scholar] [CrossRef]
- Kim, S.; Kim, E.M.; Yamamoto, M.; Park, H.; Shin, H. Engineering Multi-cellular Spheroids for Tissue Engineering and Regenerative Medicine. Adv. Healthc. Mater. 2020, 9, 2000608. [Google Scholar] [CrossRef]
- Niibe, K.; Ohori-Morita, Y.; Zhang, M.; Mabuchi, Y.; Matsuzaki, Y.; Egusa, H. A Shaking-Culture Method for Generating Bone Marrow Derived Mesenchymal Stromal/Stem Cell-Spheroids with Enhanced Multipotency in Vitro. Front. Bioeng. Biotechnol. 2020, 8, 590332. [Google Scholar] [CrossRef]
- Siltanen, C.; Yaghoobi, M.; Haque, A.; You, J.; Lowen, J.; Soleimani, M.; Revzin, A. Microfluidic Fabrication of Bioactive Microgels for Rapid Formation and Enhanced Differentiation of Stem Cell Spheroids. Acta Biomater. 2016, 34, 125–132. [Google Scholar] [CrossRef]
- San JoseLorena, H. Microfluidic Encapsulation Supports Stem Cell Viability, Proliferation, and Neuronal Differentiation. Tissue Eng. Part. C Methods 2018, 4, 158–170. [Google Scholar]
- Kim, J.A.; Choi, J.-H.; Kim, M.; Rhee, W.J.; Son, B.; Jung, H.-K.; Park, T.H. High-Throughput Generation of Spheroids Using Magnetic Nanoparticles for Three-Dimensional Cell Culture. Biomaterials 2013, 34, 8555–8563. [Google Scholar] [CrossRef]
- Santo, V.E.; Estrada, M.F.; Rebelo, S.P.; Abreu, S.; Silva, I.; Pinto, C.; Veloso, S.C.; Serra, A.T.; Boghaert, E.; Alves, P.M. Adaptable Stirred-Tank Culture Strategies for Large Scale Production of Multicellular Spheroid-Based Tumor Cell Models. J. Biotechnol. 2016, 221, 118–129. [Google Scholar] [CrossRef]
- Strube, F.; Infanger, M.; Wehland, M.; Delvinioti, X.; Romswinkel, A.; Dietz, C.; Kraus, A. Alteration of Cytoskeleton Morphology and Gene Expression in Human Breast Cancer Cells under Simulated Microgravity. Cell J. 2020, 22, 106. [Google Scholar] [PubMed]
- Lin, B.; Miao, Y.; Wang, J.; Fan, Z.; Du, L.; Su, Y.; Liu, B.; Hu, Z.; Xing, M. Surface Tension Guided Hanging-Drop: Producing Controllable 3D Spheroid of High-Passaged Human Dermal Papilla Cells and Forming Inductive Microtissues for Hair-Follicle Regeneration. ACS Appl. Mater. Interfaces 2016, 8, 5906–5916. [Google Scholar] [CrossRef] [PubMed]
- Tung, Y.-C.; Hsiao, A.Y.; Allen, S.G.; Torisawa, Y.; Ho, M.; Takayama, S. High-Throughput 3D Spheroid Culture and Drug Testing Using a 384 Hanging Drop Array. Analyst 2011, 136, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Tevis, K.M.; Cecchi, R.J.; Colson, Y.L.; Grinstaff, M.W. Mimicking the Tumor Microenvironment to Regulate Macrophage Phenotype and Assessing Chemotherapeutic Efficacy in Embedded Cancer Cell/Macrophage Spheroid Models. Acta Biomater. 2017, 50, 271–279. [Google Scholar] [CrossRef]
- Tong, W.H.; Fang, Y.; Yan, J.; Hong, X.; Singh, N.H.; Wang, S.R.; Nugraha, B.; Xia, L.; Fong, E.L.S.; Iliescu, C. Constrained Spheroids for Prolonged Hepatocyte Culture. Biomaterials 2016, 80, 106–120. [Google Scholar] [CrossRef]
- Kochanek, S.J.; Close, D.A.; Johnston, P.A. High Content Screening Characterization of Head and Neck Squamous Cell Carcinoma Multicellular Tumor Spheroid Cultures Generated in 384-Well Ultra-Low Attachment Plates to Screen for Better Cancer Drug Leads. Assay. Drug Dev. Technol. 2019, 17, 17–36. [Google Scholar] [CrossRef]
- Furukawa, K.S.; Ushida, T.; Sakai, Y.; Suzuki, M.; Tanaka, J.; Tateishi, T. Formation of Human Fibroblast Aggregates (Spheroids) by Rotational Culture. Cell Transplant. 2001, 10, 441–445. [Google Scholar] [CrossRef]
- Lee, G.; Kim, H.; Park, J.Y.; Kim, G.; Han, J.; Chung, S.; Yang, J.H.; Jeon, J.S.; Woo, D.-H.; Han, C.; et al. Generation of Uniform Liver Spheroids from Human Pluripotent Stem Cells for Imaging-Based Drug Toxicity Analysis. Biomaterials 2021, 269, 120529. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Species | Type of Cell | Method of Generation | Utility | Reference |
|---|---|---|---|---|---|
| Primary culture | Astyanax mexicanus | Hepatocyte | Wells + Shaking | Characterization Metabolism Transcriptomics | [83] |
| ZFL | Danio rerio | Hepatocyte | Scaffold-based | Toxicology | [80] |
| ZFL | Danio rerio | Hepatocyte | Magnetic levitation | Toxicology | [81] |
| ZFL | Danio rerio | Hepatocyte | Hanging drop + shaking Wells + Shaking | Characterization | [10] |
| ZFL | Danio rerio | Hepatocyte | Wells + Shaking | Characterization Transcriptomics | [82] |
| ZFL ZEM2S | Danio rerio | Hepatocyte Embryo | Wells + Shaking | Characterization | [7] |
| 14A-NFUD3, 19A-NFUD3, 15II-NFUD2, 17II-NFUD2 | Nothobranchius furzeri | Embryonic | Micromolded agarose gels | Characterization | [29] |
| 14-NKAD3, 1-NKAD3, 7A-NKAD3 | Nothobranchius kadleci | Embryonic | Micromolded agarose gels | Characterization | [29] |
| Primary culture | Oncorhynchus mykiss | Hepatocyte | Wells + Shaking | Toxicology | [67] |
| RTL-W1 | Oncorhynchus mykiss | Hepatocyte | Petri dish + Shaking | Toxicology | [72] |
| Primary culture | Oncorhynchus mykiss | Hepatocyte | Wells + Shaking | Metabolism | [73] |
| Primary culture | Oncorhynchus mykiss | Hepatocyte | Wells + Shaking | Metabolism | [74] |
| Primary culture | Oncorhynchus mykiss | Hepatocyte | Petri dish + Shaking | Metabolism | [75] |
| Primary culture | Oncorhynchus mykiss | Hepatocyte | Petri dish + Shaking | Metabolism | [76] |
| Primary culture | Oncorhynchus mykiss | Hepatocyte | Petri dish + Shaking | Metabolism | [77] |
| Primary culture | Oncorhynchus mykiss | Hepatocyte | Wells + Shaking | Characterization | [78] |
| Primary culture | Oncorhynchus mykiss | Hepatocyte | Petri dish + Shaking | Characterization | [79] |
| RTG-2 | Oncorhynchus mykiss | Gonad | Wells + Shaking | Characterization Immunology | [66] |
| RTG-2 | Oncorhynchus mykiss | Gonad | Wells + Shaking | Characterization Metabolism | [68] |
| RTgutGC | Oncorhynchus mykiss | Gastro-intestinal | Wells + Shaking | Characterization Metabolism | [69] |
| RTK | Oncorhynchus mykiss | Kidney | Wells + Shaking | Characterization Immunology | [70] |
| RTHEP | Oncorhynchus mykiss | Liver | Wells | Toxicology | [71] |
| PLHC-1 | Poeciliopsis lucida | Hepatocyte | Wells | Toxicology | [31] |
| PLHC-1 | Poeciliopsis lucida | Hepatocyte | Wells | Toxicology | [30] |
| PLHC-1 | Poeciliopsis lucida | Hepatocyte | Wells | Toxicology | [86] |
| Primary culture | Salmo trutta | Hepatocyte | Wells + Shaking | Characterization | [44] |
| Primary culture | Salmo trutta | Hepatocyte | Wells + Shaking | Characterization Metabolism | [28] |
| deSc | Stephanolepis cirrhifer | Fibroblast | Flask | Characterization | [84] |
| Primary culture | Takifugu rubripes | Pituitary | Spinner flask and spheroid dish | Endocrinology | [85] |
| Method | Advantages | Disadvantages | References |
|---|---|---|---|
| Hanging drop |
|
| [37,88,89] |
| Low-adherence substrates |
|
| [37,90] |
| Wells |
|
| [37,66,91] |
| Microfluidics |
|
| [92,93] |
| Magnetic levitation |
|
| [37,39,94] |
| Spinner flask |
|
| [95,96] |
| Cell Line | Drug | Results | Reference |
|---|---|---|---|
| Primary brown trout hepatocyte culture | Estradiol (10−6 M) | mRNAs of trout estrogen receptor and vitellogenin stable over time with similar levels to in vivo. Continued and efficient production and release of vitellogenin throughout the entire culture period. | [79] |
| Primary brown trout hepatocyte culture | β-naphthoflavone (0.36 μM) | 7-Ethoxyresorufin O-deethylase (EROD) activity not inducted in 3-day spheroids, whilst significant increase was seen in 28-day 3D cultures. | [77] |
| Primary brown trout hepatocyte culture | β-naphthoflavone (0.36 μM) Testosterone (0.17 μM) | Cytochrome P450 levels lowered until day 5; since then, remained stable one month. EROD increased significantly for one month, resembling data obtained from fresh hepatocytes. Testosterone hydroxylase activity was 30% of the activity found in fresh brown trout hepatocytes. | [74] |
| Primary brown trout hepatocyte culture | 7-ethoxyresorufin (8 µM) 7-ethoxycoumarin (100 µM) Ibuprofen sodium salt (10 µM) | Higher EROD levels than in hepatocyte cultures but lower than freshly isolated hepatocytes. Spheroids should increase predictability rather than S9 fraction because of their prolonged metabolic activity. | [75] |
| Primary brown trout hepatocyte culture | Fluorescent compounds and specific inhibitors | Positive functionality of efflux transporters in mature spheroids. | [76] |
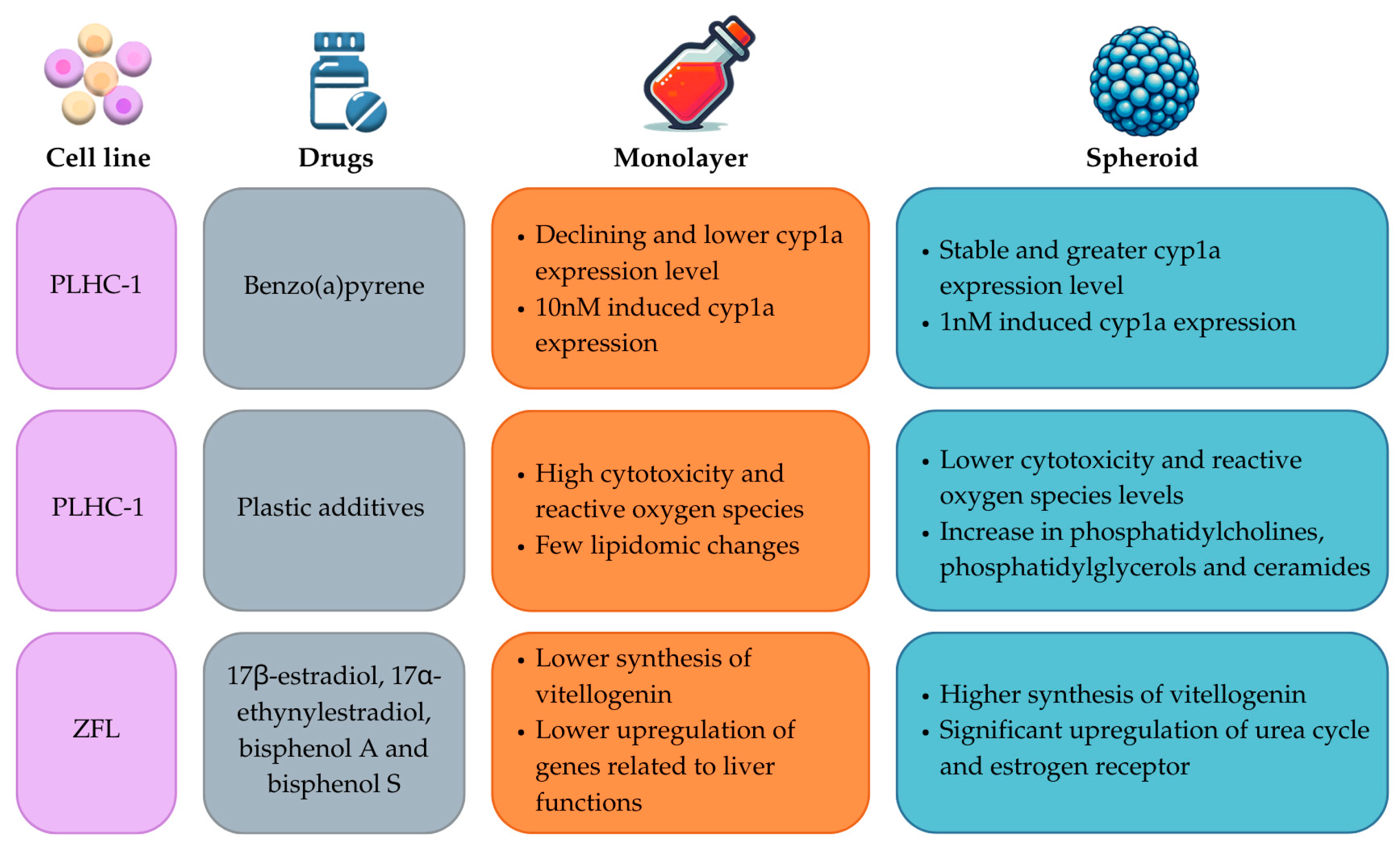
| PLHC-1 | Benzo(a)pyrene (1 nM) | Cell death and decrease in Cyp1a expression. | [31] |
| Primary brown trout hepatocyte culture | Atenolol, carbamazepine, diazepam, diclofenac, metoprolol, phenylbutazone, and propranolol (100 μg∙L−1) | Diclofenac intrinsic hepatic clearance was similar to the S9 fraction, while propranolol was 5-fold higher. Atenolol, metoprolol, diazepam, and carbamazepine not metabolized by spheroids. | [73] |
| Primary brown trout hepatocyte culture | Pyrene (25 nM) | The optimal number of spheroids per reaction was 100 and duration was 30 h. Effective biotransformation of pyrene into OH-PYR-Glu metabolite by spheroids from 2 h to 30 h of exposure. More prolonged duration of exposure than fraction S9 is needed, but also, the data resemble in vivo conditions more. | [67] |
| ZFL | 17β-estradiol, 17α-ethynylestradiol, bisphenol A, and bisphenol S (0.0002, 0.002 and 2 mM) | Significant upregulation of several genes in spheroids compared to 2D cultures such as ctnnb1, urea cycle, hepatic cytochrome P450, glycogen and glucose metabolism, nuclear receptors, and transcriptional factors. Increase in vitellogenesis over time. | [82] |
| Primary brown trout hepatocyte culture | 5α-dihydrotestosterone (DHT) (10 and 100 µM) | Spheroids treated with the highest DHT concentration (100 µM) decreased sphericity and loss of 3D dense disposal; also, PPARγ and Acox1-3I genes were downregulated. For both treatments, there was an increase in the immunochemistry signal for caspase-3, upregulation of the Acsl1 gene. The treatment with lower dose upregulated Fabp1 gene. | [28] |
| RTH-149 | Carbamazepine, propranolol, clozapine, fluoxetine, haloperidol, levomepromazine, quetiapine, sertraline, venlafaxine, clotrimazole, ketoconazole, diclofenac, ibuprofen, naproxen (9 concentrations ranging from 0.78 to 1000 μM) | Sensitivity of spheroids to sertraline, fluoxetine, levomepromazine, quetiapine, and diclofenac, especially the first three with cell viabilities (EC50s) ≤ 10 µM. The spheroids became less susceptible with the time of culture except for fluoxetine, which was affected significantly more compared to the spheroids at 72 h post-culture than 24 h. | [71] |
| Primary rainbow trout hepatocyte culture | Carbamazepine, propranolol, clozapine, fluoxetine, haloperidol, levomepromazine, quetiapine, sertraline, venlafaxine, clotrimazole, ketoconazole, diclofenac, ibuprofen, naproxen (9 concentrations ranging from 0.78 to 1000 μM) | Lower toxicity of compounds than in RTH-149 spheroids, with clozapine and haloperidol not having toxic effects. All the compounds had higher EC50s compared with RTH-149 cell line, except for ketoconazole. | [71] |
| PLHC-1 | di(2-ethylhexyl) phthalate (DEHP), dibutyl phthalate (DBP), bisphenol A (BPA), bisphenol F (BPF), 4-tert-octylphenol (OP), 4-tert-nonylphenol (NP), bisphenol A bis(3-chloro-2-hydroxypropyl) ether (BADGE 2HCl), triclosan (TCS), 3,3,5,5’-tetrabromobisphenol A (TBBPA), tritolyl phosphate (TPP) (Concentrations ranging from 1 to 50 µM) | Plastic mixture showed a decrease in EC50 in the spheroids, but not as much as in monolayer culture. A smaller amount of induced ROS species was found in spheroids compared with monolayer cultures. Enrichment of ceramides and upregulation of 19 lipids and downregulation of 6 lipids in spheroids lipidomic response. Increase in phosphatidylcholines, phosphatidylethanolamines, and ceramides, and decrease in cholesteryl esters. Increase in phosphatidylcholines/phosphatidylethanolamines ratios. | [30] |
| PLHC-1 | bis(2-ethylhexyl) phthalate (DEHP), dibutyl phthalate (DBP), bisphenol A (BPA), bisphenol F (BPF), 4-tert-octylphenol (OP), 4-tert-nonylphenol (NP), bisphenol A bis(3-chloro-2-hydroxypropyl) ether (BADGE·2HCl), triclosan (TCS), 3,3′,5,5′-tetrabromobisphenol A (TBBPA), and tritolyl phosphate (TPP) (Concentrations ranging from 1 to 50 µM) | Spheroids cultured in absence of fetal bovine serum (FBS) had an upregulation of phosphatidylcholines, alkyl, and alkenyl ether-linked ceramides and a downregulation of ether-linked phosphatidylethanolamines and phosphatidylglycerols; meanwhile, spheroids with included FBS in culture media showed only a decrease in diglycerides and ether-linked phosphatidylethanolamines, which shows a greater effect of the plastic additives to the spheroids with no FBS in culture media. Also, plastic additives had a higher cytotoxicity in cells without FBS. | [86] |
| Cell line | Pathogen | Results | Reference |
| RTG-2 | Saprolegnia parasitica | Positive viability of spheroids. Successful infiltration of mycelia into the spheroids. | [66] |
| RTK-1 | Stripped jack Nervous Necrosis Virus (SJNNV) | Visible viral infection at 2 days post-infection (dpi) in spheroids; 60% of spheroid cells tested positive for SJNNV and expressed viral intracellular protein. Higher production of SJNNV in spheroids rather than in 2D culture. | [70] |
| Cell line | Hormone | Results | Reference |
| Torafugu primary pituitary spheroids | Torafugu serum | Luteinizing hormone synthesis was found to be dose-dependent with torafugu serum, while follicle-stimulating hormone levels did not correlate with serum exposure. During puberty onset the proliferation of the pituitary spheroids was at its maxim. | [85] |
| Cell line | Drug | Results | Reference |
| ZFL | Carbamazepine | High CYP1a1 activity in spheroids, superior to monolayer culture. Upregulation of metabolite carbamazepine-10,11-epoxide in spheroids. | [81] |
| RTgutGC | Pharmaceutical compounds | Oxygen-viable zone within the spheroid reduced over time and hypoxic necrotic zone expanded with greater sizes. Spheroids with diameters less than 200 μm suggested for metabolic studies. | [69] |
| Hepatocyte primary culture | Resazurin | Stable metabolic activity of the spheroids, especially between days 12 and 20. Metabolism and detoxification, efflux transport, and estrogenic signaling results were viable for toxicological xenobiotic research. | [44] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Mercader, A.; Monzón-Atienza, L.; Montero, D.; Bravo, J.; Acosta, F. Fish Cell Spheroids, a Promising In Vitro Model to Mimic In Vivo Research: A Review. Cells 2024, 13, 1818. https://doi.org/10.3390/cells13211818
Gómez-Mercader A, Monzón-Atienza L, Montero D, Bravo J, Acosta F. Fish Cell Spheroids, a Promising In Vitro Model to Mimic In Vivo Research: A Review. Cells. 2024; 13(21):1818. https://doi.org/10.3390/cells13211818
Chicago/Turabian StyleGómez-Mercader, Antonio, Luis Monzón-Atienza, Daniel Montero, Jimena Bravo, and Félix Acosta. 2024. "Fish Cell Spheroids, a Promising In Vitro Model to Mimic In Vivo Research: A Review" Cells 13, no. 21: 1818. https://doi.org/10.3390/cells13211818
APA StyleGómez-Mercader, A., Monzón-Atienza, L., Montero, D., Bravo, J., & Acosta, F. (2024). Fish Cell Spheroids, a Promising In Vitro Model to Mimic In Vivo Research: A Review. Cells, 13(21), 1818. https://doi.org/10.3390/cells13211818