
Blastocyst-Derived Lactic Acid May Regulate S100A6 Expression and Function in Mouse Decidualization via Stimulation of Uterine Epithelial Arachidonic Acid Secretion

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. S100A6 Expression in Early Pregnant Mouse Uteri
3.2. Active Blastocyst Induces S100A6 Expression
3.3. Blastocyst-Derived Lactic Acid Regulation of S100A6 Expression Is Dependent on the AA/COX2/PGI2/PPAR δ Pathway
3.4. Effects of S100A6 on Mouse In Vitro Decidualization
3.5. S100A6 Regulates Decidualization via RAGE
3.6. S100A6 Mediates Decidualization via RAGE/EGFR/ERK1/2
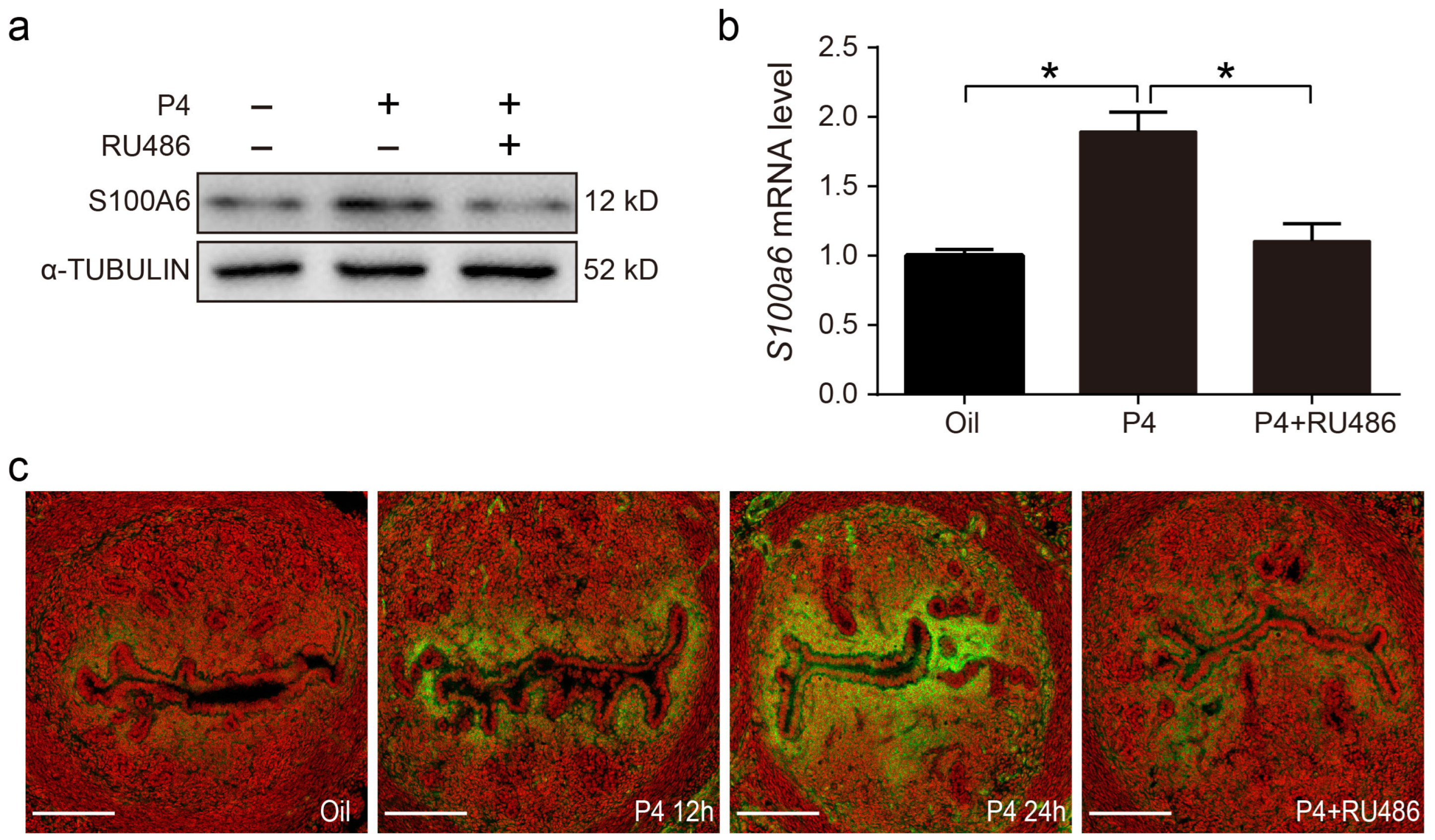
3.7. Progesterone Regulates S100A6 Expression via the Progesterone Receptor (PR)
3.8. Estrogen Regulates S100A6 Expression in an ERα-Dependent Manner
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AA | Arachidonic acid |
| COX2 | Cyclooxygenase 2 |
| EGFR | Epidermal growth-factor receptor |
| HMGB1 | High-mobility-group box 1 |
| MET | Mesenchymal-to-epithelial transition |
| PGI2 | Prostacyclin |
| PPAR δ | Peroxisome proliferator-activated receptor |
| Prl8a2 | Prolactin family 8, subfamily A, member 2 |
| RAGE | Receptor for advanced glycation end products |
References
- Ochoa-Bernal, M.A.; Fazleabas, A.T. Physiologic Events of Embryo Implantation and Decidualization in Human and Non-Human Primates. Int. J. Mol. Sci. 2020, 21, 1973. [Google Scholar] [CrossRef] [PubMed]
- Quenby, S. Implantation involves an initial pro-inflammatory response. Fertil. Steril. 2020, 114, 288–289. [Google Scholar] [CrossRef] [PubMed]
- Mor, G.; Cardenas, I. Inflammation and pregnancy: The role of the immune system at the implantation site. Ann. N. Y. Acad. Sci. 2011, 1221, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Erlebacher, A. Immunology of the maternal-fetal interface. Annu. Rev. Immunol. 2013, 31, 387–411. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhu, W. Early transient suppression of immune checkpoint proteins T-cell immunoglobulin mucin-3 and programmed cell death-1 in peripheral blood lymphocytes after blastocyst transfer is associated with successful implantation. Fertil. Steril. 2020, 114, 426–435. [Google Scholar] [CrossRef]
- D’Ippolito, S.; Tersigni, C. Inflammosome in the human endometrium: Further step in the evaluation of the “maternal side”. Fertil. Steril. 2016, 105, 111–118. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, T. Successful implantation is associated with a transient increase in serum proinflammatory cytokine profile followed by a switch to anti-inflammatory cytokine profile prior to confirmation of pregnancy. Fertil. Steril. 2021, 115, 1044–1053. [Google Scholar] [CrossRef]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef]
- Kennedy, T.G.; Gillio-Meina, C. Prostaglandins and the initiation of blastocyst implantation and decidualization. Reproduction 2007, 134, 635–643. [Google Scholar] [CrossRef]
- Brown, N.; Morrow, J.D. Restoration of on-time embryo implantation corrects the timing of parturition in cytosolic phospholipase A2 group IVA deficient mice. Biol. Reprod. 2009, 81, 1131–1138. [Google Scholar] [CrossRef]
- Lim, H.; Paria, B.C. Multiple female reproductive failures in cyclooxygenase 2 deficient mice. Cell 1997, 91, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Donato, R.; Sorci, G. S100A6 protein: Functional roles. Cell. Mol. Life Sci. 2017, 74, 2749–2760. [Google Scholar] [CrossRef] [PubMed]
- Ridinger, K.; Ilg, E.C. Clustered organization of S100 genes in human and mouse. Biochim. Biophys. Acta 1998, 1448, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Du, G. The S100 protein family in lung cancer. Clin. Chim. Acta 2021, 520, 67–70. [Google Scholar] [CrossRef]
- Saiki, Y.; Horii, A. Multiple functions of S100A10, an important cancer promoter. Pathol. Int. 2019, 69, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Braunstein, Z. S100 Proteins As an Important Regulator of Macrophage Inflammation. Front. Immunol. 2017, 8, 1908. [Google Scholar] [CrossRef] [PubMed]
- Sreejit, G.; Flynn, M.C. S100 family proteins in inflammation and beyond. Adv. Clin. Chem. 2020, 98, 173–231. [Google Scholar] [CrossRef]
- Tong, H.; Wang, L. S100A6 Activates Kupffer Cells via the p-P38 and p-JNK Pathways to Induce Inflammation, Mononuclear/macrophage Infiltration Sterile Liver Injury in Mice. Inflammation 2023, 46, 534–554. [Google Scholar] [CrossRef]
- Zhang, S.P.; Wu, Y.W. Up-regulation of RAGE and S100A6 in rats exposed to cigarette smoke. Environ. Toxicol. Pharmacol. 2009, 28, 259–264. [Google Scholar] [CrossRef]
- Tanabe, N.; Tomita, K. Co-Stimulation of AGEs and LPS Induces Inflammatory Mediators through PLCγ1/JNK/NF-κB Pathway in MC3T3-E1 Cells. Cells 2023, 12, 1383. [Google Scholar] [CrossRef]
- Yanai, T.; Tanaka, T. Immunohistochemical demonstration of cyclooxygenase-2 in glomus tumors. J. Bone Jt. Surg. Am. 2013, 95, 725–728. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.Y.; Wu, Y. In situ Synthesized Monosodium Urate Crystal Enhances Endometrium Decidualization via Sterile Inflammation During Pregnancy. Front. Cell Dev. Biol. 2021, 9, 702590. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, S.T. The regulation and function of acetylated high-mobility group box 1 during implantation and decidualization. Front. Immunol. 2023, 14, 1024706. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.W.; Chen, Z.C. Blastocyst-induced ATP release from luminal epithelial cells initiates decidualization through the P2Y2 receptor in mice. Sci. Signal. 2020, 13, 3396. [Google Scholar] [CrossRef] [PubMed]
- Thordarson, G.; Southard, J.N. Purification and characterization of mouse decidual calcyclin: A novel stimulator of mouse placental lactogen-II secretion. Endocrinology 1991, 129, 1257–1265. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.Y.; Liu, C.K. BHPF exposure impairs mouse and human decidualization. Environ. Pollut. 2022, 304, 119222. [Google Scholar] [CrossRef]
- Fu, T.; Zheng, H.T. Oncostatin M expression in the mouse uterus during early pregnancy promotes embryo implantation and decidualization. FEBS Lett. 2019, 593, 2040–2050. [Google Scholar] [CrossRef]
- Song, Z.; Li, B. Caveolin-1 Regulation and Function in Mouse Uterus during Early Pregnancy and under Human In Vitro Decidualization. Int. J. Mol. Sci. 2022, 23, 3699. [Google Scholar] [CrossRef]
- Liu, Y.F.; Li, M.Y. ERα-dependent stimulation of LCN2 in uterine epithelium during mouse early pregnancy. Reproduction 2020, 159, 493–501. [Google Scholar] [CrossRef]
- Yang, Z.S.; Pan, H.Y. Regulation and Function of Laminin A5 during Mouse and Human Decidualization. Int. J. Mol. Sci. 2021, 23, 199. [Google Scholar] [CrossRef]
- Gu, X.W.; Yang, Y. ATP mediates the interaction between human blastocyst and endometrium. Cell Prolif. 2020, 53, e12737. [Google Scholar] [CrossRef] [PubMed]
- Gardner, D.K. Lactate production by the mammalian blastocyst: Manipulating the microenvironment for uterine implantation and invasion? Bioessays 2015, 37, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, C.A.; Orwig, K.E. Dual expression of prolactin-related protein in decidua and trophoblast tissues during pregnancy in rats. Biol. Reprod. 1997, 56, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Leclerc, E.; Fritz, G. Binding of S100 proteins to RAGE: An update. Biochim. Biophys. Acta 2009, 1793, 993–1007. [Google Scholar] [CrossRef] [PubMed]
- Graczyk-Jarzynka, A.; Sobiak, B. S100A6 activates EGFR and its downstream signaling in HaCaT keratinocytes. J. Cell. Physiol. 2019, 234, 17561–17569. [Google Scholar] [CrossRef] [PubMed]
- DeMayo, F.J.; Lydon, J.P. New insights into progesterone receptor signaling in the endometrium required for embryo implantation. J. Mol. Endocrinol. 2020, 65, T1–T14. [Google Scholar] [CrossRef]
- Naderain, H.; Khanlarkhani, N. Comparison of the effects of progesterone and 17 beta-estradiol on Schwann cell markers expression in rat adipose-derived stem cells. Vet. Res. Forum 2018, 9, 307–313. [Google Scholar] [CrossRef]
- Ma, W.G.; Song, H. Estrogen is a critical determinant that specifies the duration of the window of uterine receptivity for implantation. Proc. Natl. Acad. Sci. USA 2003, 100, 2963–2968. [Google Scholar] [CrossRef]
- Hong, E.J.; Park, S.H. Identification of estrogen regulated genes by microarray analysis of the uterus of immature rats exposed to endocrine disrupting chemicals. Reprod. Biol. Endocrinol. 2006, 4, 49. [Google Scholar] [CrossRef]
- Baker, J.R.; Jeffery, R. Distinct roles for S100a8 in early embryo development and in the maternal deciduum. Dev. Dyn. 2011, 240, 2194–2203. [Google Scholar] [CrossRef]
- Rai, A.; Poh, Q.H. Proteomic profiling of human uterine extracellular vesicles reveal dynamic regulation of key players of embryo implantation and fertility during menstrual cycle. Proteomics 2021, 21, e2000211. [Google Scholar] [CrossRef] [PubMed]
- Sadigh, A.R.; Mihanfar, A. S100 protein family and embryo implantation. J. Cell. Biochem. 2019, 120, 19229–19244. [Google Scholar] [CrossRef]
- Xie, J.; Mendez, J.D. Cellular signalling of the receptor for advanced glycation end products (RAGE). Cell Signal. 2013, 25, 2185–2197. [Google Scholar] [CrossRef] [PubMed]
- Jurewicz, E.; Wyroba, E. Tubulin-dependent secretion of S100A6 and cellular signaling pathways activated by S100A6-integrin beta1 interaction. Cell Signal. 2018, 42, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Chavakis, T.; Bierhaus, A. The pattern recognition receptor (RAGE) is a counterreceptor for leukocyte integrins: A novel pathway for inflammatory cell recruitment. J. Exp. Med. 2003, 198, 1507–1515. [Google Scholar] [CrossRef] [PubMed]
- Hudson, B.I.; Lippman, M.E. Targeting RAGE Signaling in Inflammatory Disease. Annu. Rev. Med. 2018, 69, 349–364. [Google Scholar] [CrossRef]
- Sims, G.P.; Rowe, D.C. HMGB1 and RAGE in inflammation and cancer. Annu. Rev. Immunol. 2010, 28, 367–388. [Google Scholar] [CrossRef]
- Lee, C.H.; Kim, T.H. Extracellular signal-regulated kinase 1/2 signaling pathway is required for endometrial decidualization in mice and human. PLoS ONE 2013, 8, e75282. [Google Scholar] [CrossRef]
- Liu, J.; Gao, F. HB-EGF regulates Prss56 expression during mouse decidualization via EGFR/ERK/EGR2 signaling pathway. J. Endocrinol. 2017, 234, 247–254. [Google Scholar] [CrossRef]
- He, B.; Zhang, H. Blastocyst activation engenders transcriptome reprogram affecting X-chromosome reactivation and inflammatory trigger of implantation. Proc. Natl. Acad. Sci. USA 2019, 116, 16621–16630. [Google Scholar] [CrossRef]
- Joo, J.H.; Kim, J.W. Involvement of NF-kappaB in the regulation of S100A6 gene expression in human hepatoblastoma cell line HepG2. Biochem. Biophys. Res. Commun. 2003, 307, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Lesniak, W.; Szczepanska, A. Calcyclin (S100A6) expression is stimulated by agents evoking oxidative stress via the antioxidant response element. Biochim. Biophys. Acta 2005, 1744, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Leśniak, W.; Filipek, A. S100A6 Protein Expression and Function in Norm and Pathology. Int. J. Mol. Sci. 2023, 24, 1341. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.; Dey, S.K. PPAR delta functions as a prostacyclin receptor in blastocyst implantation. Trends Endocrinol. Metab. 2000, 11, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Pakrasi, P.L.; Jain, A.K. Cyclooxygenase-2 derived PGE2 and PGI2 play an important role via EP2 and PPARdelta receptors in early steps of oil induced decidualization in mice. Placenta 2008, 29, 523–530. [Google Scholar] [CrossRef]
- Chakraborty, I.; Das, S.K. Developmental expression of the cyclo-oxygenase-1 and cyclo-oxygenase-2 genes in the peri-implantation mouse uterus and their differential regulation by the blastocyst and ovarian steroids. J. Mol. Endocrinol. 1996, 16, 107–122. [Google Scholar] [CrossRef]
- Lim, H.; Gupta, R.A. Cyclo-oxygenase-2-derived prostacyclin mediates embryo implantation in the mouse via PPARdelta. Genes Dev. 1999, 13, 1561–1574. [Google Scholar] [CrossRef]
- Mor, G.; Abrahams, V.M. Potential role of macrophages as immunoregulators of pregnancy. Reprod. Biol. Endocrinol. 2003, 1, 119. [Google Scholar] [CrossRef]
- Gnainsky, Y.; Granot, I. Local injury of the endometrium induces an inflammatory response that promotes successful implantation. Fertil. Steril. 2010, 94, 2030–2036. [Google Scholar] [CrossRef]
- Li, D.K.; Liu, L. Exposure to non-steroidal anti-inflammatory drugs during pregnancy and risk of miscarriage: Population based cohort study. BMJ 2003, 327, 368. [Google Scholar] [CrossRef]
- Zhao, J.; Endoh, I. S100A8 modulates mast cell function and suppresses eosinophil migration in acute asthma. Antioxid. Redox Signal. 2011, 14, 1589–1600. [Google Scholar] [CrossRef] [PubMed]
- Tong, L.; Lan, W. S100A proteins as molecular targets in the ocular surface inflammatory diseases. Ocul. Surf. 2014, 12, 23–31. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.-Y.; Wang, Y.; Wu, Y.; Zhao, X.-Y.; Yang, Z.-S.; Li, B.; Chen, S.-T.; He, Y.-Y.; Yang, Z.-M. Blastocyst-Derived Lactic Acid May Regulate S100A6 Expression and Function in Mouse Decidualization via Stimulation of Uterine Epithelial Arachidonic Acid Secretion. Cells 2024, 13, 206. https://doi.org/10.3390/cells13030206
Li M-Y, Wang Y, Wu Y, Zhao X-Y, Yang Z-S, Li B, Chen S-T, He Y-Y, Yang Z-M. Blastocyst-Derived Lactic Acid May Regulate S100A6 Expression and Function in Mouse Decidualization via Stimulation of Uterine Epithelial Arachidonic Acid Secretion. Cells. 2024; 13(3):206. https://doi.org/10.3390/cells13030206
Chicago/Turabian StyleLi, Meng-Yuan, Ying Wang, Ying Wu, Xu-Yu Zhao, Zhen-Shan Yang, Bo Li, Si-Ting Chen, Yu-Ying He, and Zeng-Ming Yang. 2024. "Blastocyst-Derived Lactic Acid May Regulate S100A6 Expression and Function in Mouse Decidualization via Stimulation of Uterine Epithelial Arachidonic Acid Secretion" Cells 13, no. 3: 206. https://doi.org/10.3390/cells13030206