
Chemically Functionalized Single-Walled Carbon Nanotubes Prevent the Reduction in Plasmalemmal Glutamate Transporter EAAT1 Expression in, and Increase the Release of Selected Cytokines from, Stretch-Injured Astrocytes in Vitro
 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. In Vitro Experiments
2.2.1. Astrocyte Culture
2.2.2. Cell Injury Model
2.2.3. Single-Walled Carbon Nanotubes Preparation
2.2.4. Incubation of the Primary Astrocyte Culture with SWCNT-PEG or the Functionalization Agent PEG
2.3. Determination of Severity of the Astrocyte Injury
2.4. Dot Blot
2.5. Western Blotting
2.6. Cytokine Assay
2.7. Statistical Analyses
3. Results
3.1. SWCNT-PEG Do Not Affect Cell Viability of Astrocytes Exposed to Severe Stretch Injury
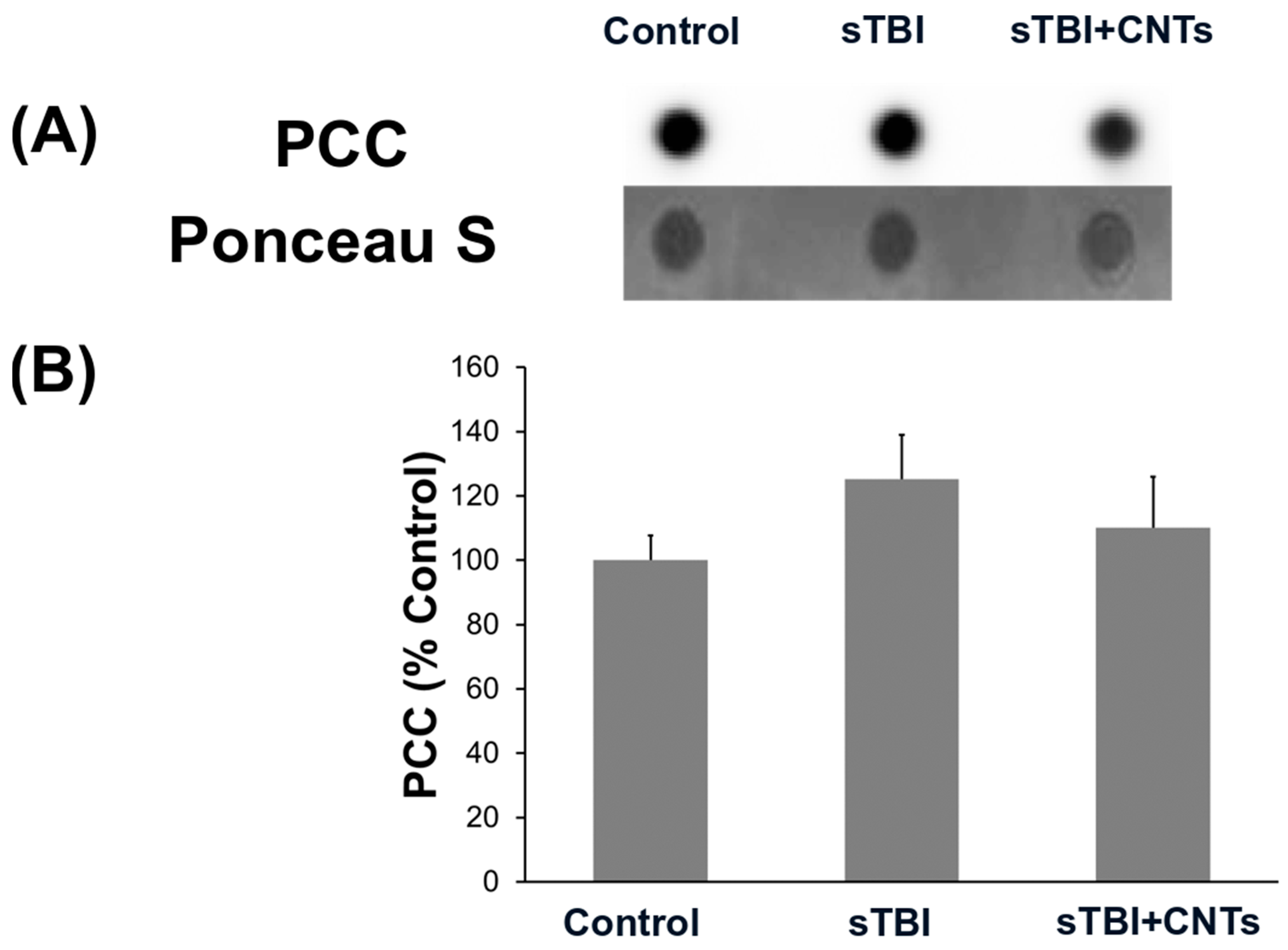
3.2. Severe Stretch Injury of Cultured Astrocytes and the Application of SWCNT-PEG Do Not Cause Changes in the Protein Oxidation Levels
3.3. Application of SWCNT-PEG to Astrocytes Rescues the Injury-Induced Loss of Plasmalemmal Glutamate Transporter EAAT1
3.4. Cytokine Array
4. Discussion
4.1. SWCNT-PEG Do Not Affect Viability of Astrocytes Exposed to Severe Stretch Injury
4.2. Application of SWCNT-PEG to Astrocytes Exposed to Severe Stretch Injury Prevents the Decrease in the Expression of EAAT1 in Severely Stretched Astrocytes
4.3. SWCNT-PEG Modulate the Secretory Function of Astrocytes Exposed to Severe Stretch Injury
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meaney, D.F.; Morrison, B.; Dale Bass, C. The Mechanics of Traumatic Brain Injury: A Review of What We Know and What We Need to Know for Reducing Its Societal Burden. J. Biomech. Eng. 2014, 136, 021008. [Google Scholar] [CrossRef] [PubMed]
- Khellaf, A.; Khan, D.Z.; Helmy, A. Recent Advances in Traumatic Brain Injury. J. Neurol. 2019, 266, 2878–2889. [Google Scholar] [CrossRef] [PubMed]
- Gruenbaum, B.F.; Zlotnik, A.; Fleidervish, I.; Frenkel, A.; Boyko, M. Glutamate Neurotoxicity and Destruction of the Blood–Brain Barrier: Key Pathways for the Development of Neuropsychiatric Consequences of TBI and Their Potential Treatment Strategies. Int. J. Mol. Sci. 2022, 23, 9628. [Google Scholar] [CrossRef] [PubMed]
- Cederberg, D.; Visse, E.; Marklund, N.; Siesjö, P. Prolonged and Intense Neuroinflammation after Severe Traumatic Brain Injury Assessed by Cerebral Microdialysis with 300 KDa Membranes. J. Neuroimmunol. 2023, 377, 578020. [Google Scholar] [CrossRef] [PubMed]
- Betancur, M.I.; Mason, H.D.; Alvarado-Velez, M.; Holmes, P.V.; Bellamkonda, R.V.; Karumbaiah, L. Chondroitin Sulfate Glycosaminoglycan Matrices Promote Neural Stem Cell Maintenance and Neuroprotection Post-Traumatic Brain Injury. ACS Biomater. Sci. Eng. 2017, 3, 420–430. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Shao, A.; Yao, Y.; Tu, S.; Deng, Y.; Zhang, J. Dual Roles of Astrocytes in Plasticity and Reconstruction after Traumatic Brain Injury. Cell Commun. Signal. 2020, 18, 62. [Google Scholar] [CrossRef] [PubMed]
- Escartin, C.; Galea, E.; Lakatos, A.; O’Callaghan, J.P.; Petzold, G.C.; Serrano-Pozo, A.; Steinhäuser, C.; Volterra, A.; Carmignoto, G.; Agarwal, A.; et al. Reactive Astrocyte Nomenclature, Definitions, and Future Directions. Nat. Neurosci. 2021, 24, 312–325. [Google Scholar] [CrossRef]
- Lepore, A.C.; O’Donnell, J.; Kim, A.S.; Yang, E.J.; Tuteja, A.; Haidet-Phillips, A.; O’Banion, C.P.; Maragakis, N.J. Reduction in Expression of the Astrocyte Glutamate Transporter, GLT1, Worsens Functional and Histological Outcomes Following Traumatic Spinal Cord Injury. Glia 2011, 59, 1996–2005. [Google Scholar] [CrossRef]
- Allaman, I.; Bélanger, M.; Magistretti, P.J. Astrocyte-Neuron Metabolic Relationships: For Better and for Worse. Trends Neurosci. 2011, 34, 76–87. [Google Scholar] [CrossRef]
- Siracusa, R.; Fusco, R.; Cuzzocrea, S. Astrocytes: Role and Functions in Brain Pathologies. Front. Pharmacol. 2019, 10, 1114. [Google Scholar] [CrossRef]
- Pajarillo, E.; Rizor, A.; Lee, J.; Aschner, M.; Lee, E. The Role of Astrocytic Glutamate Transporters GLT-1 and GLAST in Neurological Disorders: Potential Targets for Neurotherapeutics. Neuropharmacology 2019, 161, 107559. [Google Scholar] [CrossRef]
- Furuta, A.; Rothstein, J.D.; Martin, L.J. Glutamate Transporter Protein Subtypes Are Expressed Differentially during Rat CNS Development. J. Neurosci. 1997, 17, 8363–8375. [Google Scholar] [CrossRef] [PubMed]
- Beschorner, R.; Dietz, K.; Schauer, N.; Mittelbronn, M.; Schluesener, H.J.; Trautmann, K.; Meyermann, R.; Simon, P. Expression of EAAT1 Reflects a Possible Neuroprotective Function of Reactive Astrocytes and Activated Microglia Following Human Traumatic Brain Injury. Histol. Histopathol. 2007, 22, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Piao, C.-S.; Holloway, A.L.; Hong-Routson, S.; Wainwright, M.S. Depression Following Traumatic Brain Injury in Mice Is Associated with Down-Regulation of Hippocampal Astrocyte Glutamate Transporters by Thrombin. J. Cereb. Blood Flow Metab. 2019, 39, 58–73. [Google Scholar] [CrossRef] [PubMed]
- van Landeghem, F.K.H.; Weiss, T.; Oehmichen, M.; von Deimling, A. Decreased Expression of Glutamate Transporters in Astrocytes after Human Traumatic Brain Injury. J. Neurotrauma 2006, 23, 1518–1528. [Google Scholar] [CrossRef] [PubMed]
- Xiang, C.; Zhang, Y.; Guo, W.; Liang, X.-J. Biomimetic Carbon Nanotubes for Neurological Disease Therapeutics as Inherent Medication. Acta Pharm. Sin. B 2020, 10, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Hu, H.; Malarkey, E.B.; Zhao, B.; Montana, V.; Haddon, R.C.; Parpura, V. Chemically Functionalized Water Soluble Single-Walled Carbon Nanotubes Modulate Neurite Outgrowth. J. Nanosci. Nanotechnol. 2005, 5, 1707–1712. [Google Scholar] [CrossRef]
- Gottipati, M.K.; Kalinina, I.; Bekyarova, E.; Haddon, R.C.; Parpura, V. Chemically Functionalized Water-Soluble Single-Walled Carbon Nanotubes Modulate Morpho-Functional Characteristics of Astrocytes. Nano Lett. 2012, 12, 4742–4747. [Google Scholar] [CrossRef]
- Gottipati, M.K.; Bekyarova, E.; Haddon, R.C.; Parpura, V. Chemically Functionalized Single-Walled Carbon Nanotubes Enhance the Glutamate Uptake Characteristics of Mouse Cortical Astrocytes. Amino Acids 2015, 47, 1379–1388. [Google Scholar] [CrossRef]
- Roman, J.A.; Niedzielko, T.L.; Haddon, R.C.; Parpura, V.; Floyd, C.L. Single-Walled Carbon Nanotubes Chemically Functionalized with Polyethylene Glycol Promote Tissue Repair in a Rat Model of Spinal Cord Injury. J. Neurotrauma 2011, 28, 2349–2362. [Google Scholar] [CrossRef]
- Wanner, I.-B. An in Vitro Trauma Model to Study Rodent and Human Astrocyte Reactivity. Methods Mol. Biol. 2012, 814, 189–219. [Google Scholar] [CrossRef] [PubMed]
- Ellis, E.F.; McKinney, J.S.; Willoughby, K.A.; Liang, S.; Povlishock, J.T. A New Model for Rapid Stretch-Induced Injury of Cells in Culture: Characterization of the Model Using Astrocytes. J. Neurotrauma 1995, 12, 325–339. [Google Scholar] [CrossRef] [PubMed]
- Gottipati, M.K.; Verkhratsky, A.; Parpura, V. Probing Astroglia with Carbon Nanotubes: Modulation of Form and Function. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130598. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Ruijter, J.M.; Thygesen, H.H.; Schoneveld, O.J.L.M.; Das, A.T.; Berkhout, B.; Lamers, W.H. Factor Correction as a Tool to Eliminate Between-Session Variation in Replicate Experiments: Application to Molecular Biology and Retrovirology. Retrovirology 2006, 3, 2. [Google Scholar] [CrossRef] [PubMed]
- Bardi, G.; Nunes, A.; Gherardini, L.; Bates, K.; Al-Jamal, K.T.; Gaillard, C.; Prato, M.; Bianco, A.; Pizzorusso, T.; Kostarelos, K. Functionalized Carbon Nanotubes in the Brain: Cellular Internalization and Neuroinflammatory Responses. PLoS ONE 2013, 8, e80964. [Google Scholar] [CrossRef] [PubMed]
- Wanner, I.B.; Deik, A.; Torres, M.; Rosendahl, A.; Neary, J.T.; Lemmon, V.P.; Bixby, J.L. A New in Vitro Model of the Glial Scar Inhibits Axon Growth. Glia 2008, 56, 1691–1709. [Google Scholar] [CrossRef] [PubMed]
- Greco, T.M.; Seeholzer, S.H.; Mak, A.; Spruce, L.; Ischiropoulos, H. Quantitative Mass Spectrometry-Based Proteomics Reveals the Dynamic Range of Primary Mouse Astrocyte Protein Secretion. J. Proteome Res. 2010, 9, 2764–2774. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Matteoli, M.; Parpura, V.; Mothet, J.-P.; Zorec, R. Astrocytes as Secretory Cells of the Central Nervous System: Idiosyncrasies of Vesicular Secretion. EMBO J. 2016, 35, 239–257. [Google Scholar] [CrossRef]
- Yuan, X.; Zhang, X.; Sun, L.; Wei, Y.; Wei, X. Cellular Toxicity and Immunological Effects of Carbon-Based Nanomaterials. Part. Fibre Toxicol. 2019, 16, 18. [Google Scholar] [CrossRef]
- Simon-Deckers, A.; Gouget, B.; Mayne-L’hermite, M.; Herlin-Boime, N.; Reynaud, C.; Carrière, M. In Vitro Investigation of Oxide Nanoparticle and Carbon Nanotube Toxicity and Intracellular Accumulation in A549 Human Pneumocytes. Toxicology 2008, 253, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Foldbjerg, R.; Irving, E.S.; Wang, J.; Thorsen, K.; Sutherland, D.S.; Autrup, H.; Beer, C. The Toxic Effects of Single-Walled Carbon Nanotubes Are Linked to the Phagocytic Ability of Cells. Toxicol. Res. 2014, 3, 228–241. [Google Scholar] [CrossRef]
- Nel, A.; Xia, T.; Mädler, L.; Li, N. Toxic Potential of Materials at the Nanolevel. Science 2006, 311, 622–627. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, Y.; Li, Z.; Chen, T.; Lantz, S.M.; Howard, P.C.; Paule, M.G.; Slikker, W.; Watanabe, F.; Mustafa, T.; et al. Mechanistic Toxicity Evaluation of Uncoated and PEGylated Single-Walled Carbon Nanotubes in Neuronal PC12 Cells. ACS Nano 2011, 5, 7020–7033. [Google Scholar] [CrossRef] [PubMed]
- Fukamachi, S.; Furuta, A.; Ikeda, T.; Ikenoue, T.; Kaneoka, T.; Rothstein, J.D.; Iwaki, T. Altered Expressions of Glutamate Transporter Subtypes in Rat Model of Neonatal Cerebral Hypoxia-Ischemia. Brain Res. Dev. Brain Res. 2001, 132, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Moretto, M.B.; Arteni, N.S.; Lavinsky, D.; Netto, C.A.; Rocha, J.B.T.; Souza, D.O.; Wofchuk, S. Hypoxic-Ischemic Insult Decreases Glutamate Uptake by Hippocampal Slices from Neonatal Rats: Prevention by Guanosine. Exp. Neurol. 2005, 195, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Namura, S.; Maeno, H.; Takami, S.; Jiang, X.-F.; Kamichi, S.; Wada, K.; Nagata, I. Inhibition of Glial Glutamate Transporter GLT-1 Augments Brain Edema after Transient Focal Cerebral Ischemia in Mice. Neurosci. Lett. 2002, 324, 117–120. [Google Scholar] [CrossRef]
- Rao, V.L.; Bowen, K.K.; Dempsey, R.J. Transient Focal Cerebral Ischemia Down-Regulates Glutamate Transporters GLT-1 and EAAC1 Expression in Rat Brain. Neurochem. Res. 2001, 26, 497–502. [Google Scholar] [CrossRef]
- Shen, Y.; Lu, H.; Xu, R.; Tian, H.; Xia, X.; Zhou, F.H.; Wang, L.; Dong, J.; Sun, L. The Expression of GLAST and GLT1 in a Transient Cerebral Ischemia Mongolian Gerbil Model. Neuropsychiatr. Dis. Treat. 2020, 16, 789–800. [Google Scholar] [CrossRef]
- Rao, V.L.; Başkaya, M.K.; Doğan, A.; Rothstein, J.D.; Dempsey, R.J. Traumatic Brain Injury Down-Regulates Glial Glutamate Transporter (GLT-1 and GLAST) Proteins in Rat Brain. J. Neurochem. 1998, 70, 2020–2027. [Google Scholar] [CrossRef]
- Maragakis, N.J.; Rothstein, J.D. Mechanisms of Disease: Astrocytes in Neurodegenerative Disease. Nat. Clin. Pract. Neurol. 2006, 2, 679–689. [Google Scholar] [CrossRef]
- Lau, C.L.; O’Shea, R.D.; Broberg, B.V.; Bischof, L.; Beart, P.M. The Rho Kinase Inhibitor Fasudil Up-Regulates Astrocytic Glutamate Transport Subsequent to Actin Remodelling in Murine Cultured Astrocytes. Br. J. Pharmacol. 2011, 163, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Sheean, R.K.; Lau, C.L.; Shin, Y.S.; O’Shea, R.D.; Beart, P.M. Links between L-Glutamate Transporters, Na+/K+-ATPase and Cytoskeleton in Astrocytes: Evidence Following Inhibition with Rottlerin. Neuroscience 2013, 254, 335–346. [Google Scholar] [CrossRef]
- Zagami, C.J.; O’Shea, R.D.; Lau, C.L.; Cheema, S.S.; Beart, P.M. Regulation of Glutamate Transporters in Astrocytes: Evidence for a Relationship between Transporter Expression and Astrocytic Phenotype. Neurotox. Res. 2005, 7, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Holt, B.D.; Short, P.A.; Rape, A.D.; Wang, Y.; Islam, M.F.; Dahl, K.N. Carbon Nanotubes Reorganize Actin Structures in Cells and Ex Vivo. ACS Nano 2010, 4, 4872–4878. [Google Scholar] [CrossRef] [PubMed]
- Sofroniew, M.V. Astrocyte Reactivity: Subtypes, States, and Functions in CNS Innate Immunity. Trends Immunol. 2020, 41, 758–770. [Google Scholar] [CrossRef] [PubMed]
- Lazic, A.; Balint, V.; Stanisavljevic Ninkovic, D.; Peric, M.; Stevanovic, M. Reactive and Senescent Astroglial Phenotypes as Hallmarks of Brain Pathologies. Int. J. Mol. Sci. 2022, 23, 4995. [Google Scholar] [CrossRef]
- Zhou, H.; Hu, L.; Li, J.; Ruan, W.; Cao, Y.; Zhuang, J.; Xu, H.; Peng, Y.; Zhang, Z.; Xu, C.; et al. AXL Kinase-Mediated Astrocytic Phagocytosis Modulates Outcomes of Traumatic Brain Injury. J. Neuroinflamm. 2021, 18, 154. [Google Scholar] [CrossRef]
- Semple, B.D.; Frugier, T.; Morganti-Kossmann, M.C. CCL2 Modulates Cytokine Production in Cultured Mouse Astrocytes. J. Neuroinflamm. 2010, 7, 67. [Google Scholar] [CrossRef]
- Lau, L.T.; Yu, A.C. Astrocytes Produce and Release Interleukin-1, Interleukin-6, Tumor Necrosis Factor Alpha and Interferon-Gamma Following Traumatic and Metabolic Injury. J. Neurotrauma 2001, 18, 351–359. [Google Scholar] [CrossRef]
- Choi, B.-R.; Johnson, K.R.; Maric, D.; McGavern, D.B. Monocyte-Derived IL-6 Programs Microglia to Rebuild Damaged Brain Vasculature. Nat. Immunol. 2023, 24, 1110–1123. [Google Scholar] [CrossRef] [PubMed]
- Awatsuji, H.; Furukawa, Y.; Hirota, M.; Murakami, Y.; Nii, S.; Furukawa, S.; Hayashi, K. Interleukin-4 and -5 as Modulators of Nerve Growth Factor Synthesis/Secretion in Astrocytes. J. Neurosci. Res. 1993, 34, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Gadani, S.P.; Cronk, J.C.; Norris, G.T.; Kipnis, J. IL-4 in the Brain: A Cytokine to Remember. J. Immunol. 2012, 189, 4213–4219. [Google Scholar] [CrossRef] [PubMed]
- Kamm, K.; Vanderkolk, W.; Lawrence, C.; Jonker, M.; Davis, A.T. The Effect of Traumatic Brain Injury upon the Concentration and Expression of Interleukin-1beta and Interleukin-10 in the Rat. J. Trauma 2006, 60, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.M.; Stillings, S.A.; Leclerc, J.L.; Phillips, H.; Edwards, N.J.; Robicsek, S.A.; Hoh, B.L.; Blackburn, S.; Doré, S. Role of Interleukin-10 in Acute Brain Injuries. Front. Neurol. 2017, 8, 244. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Cao, F.; Cui, L.; Ciric, B.; Zhang, G.-X.; Rostami, A. IL-9 Signaling Affects Central Nervous System Resident Cells during Inflammatory Stimuli. Exp. Mol. Pathol. 2015, 99, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, S.; Thome, R.; Konno, S.; Mari, E.R.; Rasouli, J.; Hwang, D.; Boehm, A.; Li, Y.; Zhang, G.-X.; Ciric, B.; et al. IL-9 Controls Central Nervous System Autoimmunity by Suppressing GM-CSF Production. J. Immunol. 2020, 204, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Shan, Y.; Lin, Y.; Liao, S.; Zhang, B.; Zeng, Q.; Wang, Y.; Deng, Z.; Chen, C.; Hu, X.; et al. Neutralization of Interleukin-9 Ameliorates Experimental Stroke by Repairing the Blood-Brain Barrier via down-Regulation of Astrocyte-Derived Vascular Endothelial Growth Factor-A. FASEB J. 2019, 33, 4376–4387. [Google Scholar] [CrossRef]
- Kovacic, J.C.; Muller, D.W.M.; Graham, R.M. Actions and Therapeutic Potential of G-CSF and GM-CSF in Cardiovascular Disease. J. Mol. Cell. Cardiol. 2007, 42, 19–33. [Google Scholar] [CrossRef]
- Konishi, Y.; Chui, D.H.; Hirose, H.; Kunishita, T.; Tabira, T. Trophic Effect of Erythropoietin and Other Hematopoietic Factors on Central Cholinergic Neurons in Vitro and in Vivo. Brain Res. 1993, 609, 29–35. [Google Scholar] [CrossRef]
- Schäbitz, W.-R.; Kollmar, R.; Schwaninger, M.; Juettler, E.; Bardutzky, J.; Schölzke, M.N.; Sommer, C.; Schwab, S. Neuroprotective Effect of Granulocyte Colony-Stimulating Factor after Focal Cerebral Ischemia. Stroke 2003, 34, 745–751. [Google Scholar] [CrossRef] [PubMed]
- Schneider, A.; Krüger, C.; Steigleder, T.; Weber, D.; Pitzer, C.; Laage, R.; Aronowski, J.; Maurer, M.H.; Gassler, N.; Mier, W.; et al. The Hematopoietic Factor G-CSF Is a Neuronal Ligand That Counteracts Programmed Cell Death and Drives Neurogenesis. J. Clin. Investig. 2005, 115, 2083–2098. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Dai, Y.; Ma, X.; Wang, S.; Xu, X.; Pei, X.; Li, R.; Wang, H. Different Changes in Granulocyte-Colony Stimulating Factor and Its Correlation with Inflammatory Biomarkers in Patients after Traumatic Brain Injury. NeuroReport 2020, 31, 293. [Google Scholar] [CrossRef] [PubMed]
- Corrigan, F.; Arulsamy, A.; Teng, J.; Collins-Praino, L.E. Pumping the Brakes: Neurotrophic Factors for the Prevention of Cognitive Impairment and Dementia after Traumatic Brain Injury. J. Neurotrauma 2017, 34, 971–986. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Ping, S.; Kyle, M.; Chin, L.; Zhao, L.-R. SCF + G-CSF Treatment in the Chronic Phase of Severe TBI Enhances Axonal Sprouting in the Spinal Cord and Synaptic Pruning in the Hippocampus. Acta Neuropathol. Commun. 2021, 9, 63. [Google Scholar] [CrossRef] [PubMed]
- Nishihara, T.; Ochi, M.; Sugimoto, K.; Takahashi, H.; Yano, H.; Kumon, Y.; Ohnishi, T.; Tanaka, J. Subcutaneous Injection Containing IL-3 and GM-CSF Ameliorates Stab Wound-Induced Brain Injury in Rats. Exp. Neurol. 2011, 229, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Shultz, S.R.; Tan, X.L.; Wright, D.K.; Liu, S.J.; Semple, B.D.; Johnston, L.; Jones, N.C.; Cook, A.D.; Hamilton, J.A.; O’Brien, T.J. Granulocyte-Macrophage Colony-Stimulating Factor Is Neuroprotective in Experimental Traumatic Brain Injury. J. Neurotrauma 2014, 31, 976–983. [Google Scholar] [CrossRef]
- Boraschi, D.; Italiani, P.; Weil, S.; Martin, M.U. The Family of the Interleukin-1 Receptors. Immunol. Rev. 2018, 281, 197–232. [Google Scholar] [CrossRef]
- Harrer, C.; Otto, F.; Radlberger, R.F.; Moser, T.; Pilz, G.; Wipfler, P.; Harrer, A. The CXCL13/CXCR5 Immune Axis in Health and Disease—Implications for Intrathecal B Cell Activities in Neuroinflammation. Cells 2022, 11, 2649. [Google Scholar] [CrossRef]
- Pilz, G.; Wipfler, P.; Otto, F.; Hitzl, W.; Afazel, S.; Haschke-Becher, E.; Trinka, E.; Harrer, A. Cerebrospinal Fluid CXLC13 Indicates Disease Course in Neuroinfection: An Observational Study. J. Neuroinflamm. 2019, 16, 13. [Google Scholar] [CrossRef]
- Rayasam, A.; Kijak, J.A.; Kissel, L.; Choi, Y.H.; Kim, T.; Hsu, M.; Joshi, D.; Laaker, C.J.; Cismaru, P.; Lindstedt, A.; et al. CXCL13 Expressed on Inflamed Cerebral Blood Vessels Recruit IL-21 Producing TFH Cells to Damage Neurons Following Stroke. J. Neuroinflamm. 2022, 19, 125. [Google Scholar] [CrossRef] [PubMed]
- Ciechanowska, A.; Popiolek-Barczyk, K.; Ciapała, K.; Pawlik, K.; Oggioni, M.; Mercurio, D.; de Simoni, M.G.; Mika, J. Traumatic brain injury in mice induces changes in the expression of the XCL1/XCR1 and XCL1/ITGA9 axes. Pharmacol. Rep. 2020, 72, 1579–1592. [Google Scholar] [CrossRef] [PubMed]
- Moro, N.; Ghavim, S.S.; Sutton, R.L. Massive Efflux of Adenosine Triphosphate into the Extracellular Space Immediately after Experimental Traumatic Brain Injury. Exp. Ther. Med. 2021, 21, 575. [Google Scholar] [CrossRef]
- Malarkey, E.B.; Parpura, V. Temporal Characteristics of Vesicular Fusion in Astrocytes: Examination of Synaptobrevin 2-Laden Vesicles at Single Vesicle Resolution. J. Physiol. 2011, 589, 4271–4300. [Google Scholar] [CrossRef] [PubMed]
- Montana, V.; Ni, Y.; Sunjara, V.; Hua, X.; Parpura, V. Vesicular Glutamate Transporter-Dependent Glutamate Release from Astrocytes. J. Neurosci. 2004, 24, 2633–2642. [Google Scholar] [CrossRef]
- Gottipati, M.K.; Bekyarova, E.; Haddon, R.C.; Parpura, V. Chemically Functionalized Water-Soluble Single-Walled Carbon Nanotubes Obstruct Vesicular/Plasmalemmal Recycling in Astrocytes Down-Stream of Calcium Ions. Cells 2020, 9, 1597. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Immunogen | Manufacturer; Catalog Number; RRID; Host Species and Antibody Type | Dilution |
|---|---|---|---|
| anti-GFAP | native GFAP purified from pig spinal cord | Cell Signaling Technology (Danvers, MA, USA); 3670; AB_561049; mouse monoclonal | 1:1000 |
| anti-EAAT1 | synthetic peptide corresponding to residues surrounding Glu230 of human EAAT1 | Cell Signaling Technology; 5685; AB_10694915; rabbit monoclonal | 1:1000 |
| anti-β-actin | β-actin (C4) raised against gizzard actin from chicken | Santa Cruz Biotechnology (Dallas, TX, USA); sc-47778; AB_2714189; mouse monoclonal | 1:1000 |
| biotinylated goat anti-rabbit | gamma immunoglobulins | Thermo Fisher Scientific; 65-6140; AB_2533969; goat polyclonal | 1:3000 |
| biotinylated goat anti-mouse | gamma immunoglobulins | Thermo Fisher Scientific; A16070; AB_2534743; goat polyclonal | 1:3000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gržeta Krpan, N.; Harej Hrkać, A.; Janković, T.; Dolenec, P.; Bekyarova, E.; Parpura, V.; Pilipović, K. Chemically Functionalized Single-Walled Carbon Nanotubes Prevent the Reduction in Plasmalemmal Glutamate Transporter EAAT1 Expression in, and Increase the Release of Selected Cytokines from, Stretch-Injured Astrocytes in Vitro. Cells 2024, 13, 225. https://doi.org/10.3390/cells13030225
Gržeta Krpan N, Harej Hrkać A, Janković T, Dolenec P, Bekyarova E, Parpura V, Pilipović K. Chemically Functionalized Single-Walled Carbon Nanotubes Prevent the Reduction in Plasmalemmal Glutamate Transporter EAAT1 Expression in, and Increase the Release of Selected Cytokines from, Stretch-Injured Astrocytes in Vitro. Cells. 2024; 13(3):225. https://doi.org/10.3390/cells13030225
Chicago/Turabian StyleGržeta Krpan, Nika, Anja Harej Hrkać, Tamara Janković, Petra Dolenec, Elena Bekyarova, Vladimir Parpura, and Kristina Pilipović. 2024. "Chemically Functionalized Single-Walled Carbon Nanotubes Prevent the Reduction in Plasmalemmal Glutamate Transporter EAAT1 Expression in, and Increase the Release of Selected Cytokines from, Stretch-Injured Astrocytes in Vitro" Cells 13, no. 3: 225. https://doi.org/10.3390/cells13030225
APA StyleGržeta Krpan, N., Harej Hrkać, A., Janković, T., Dolenec, P., Bekyarova, E., Parpura, V., & Pilipović, K. (2024). Chemically Functionalized Single-Walled Carbon Nanotubes Prevent the Reduction in Plasmalemmal Glutamate Transporter EAAT1 Expression in, and Increase the Release of Selected Cytokines from, Stretch-Injured Astrocytes in Vitro. Cells, 13(3), 225. https://doi.org/10.3390/cells13030225