

Advances in the Generation of Constructed Cardiac Tissue Derived from Induced Pluripotent Stem Cells for Disease Modeling and Therapeutic Discovery


Abstract
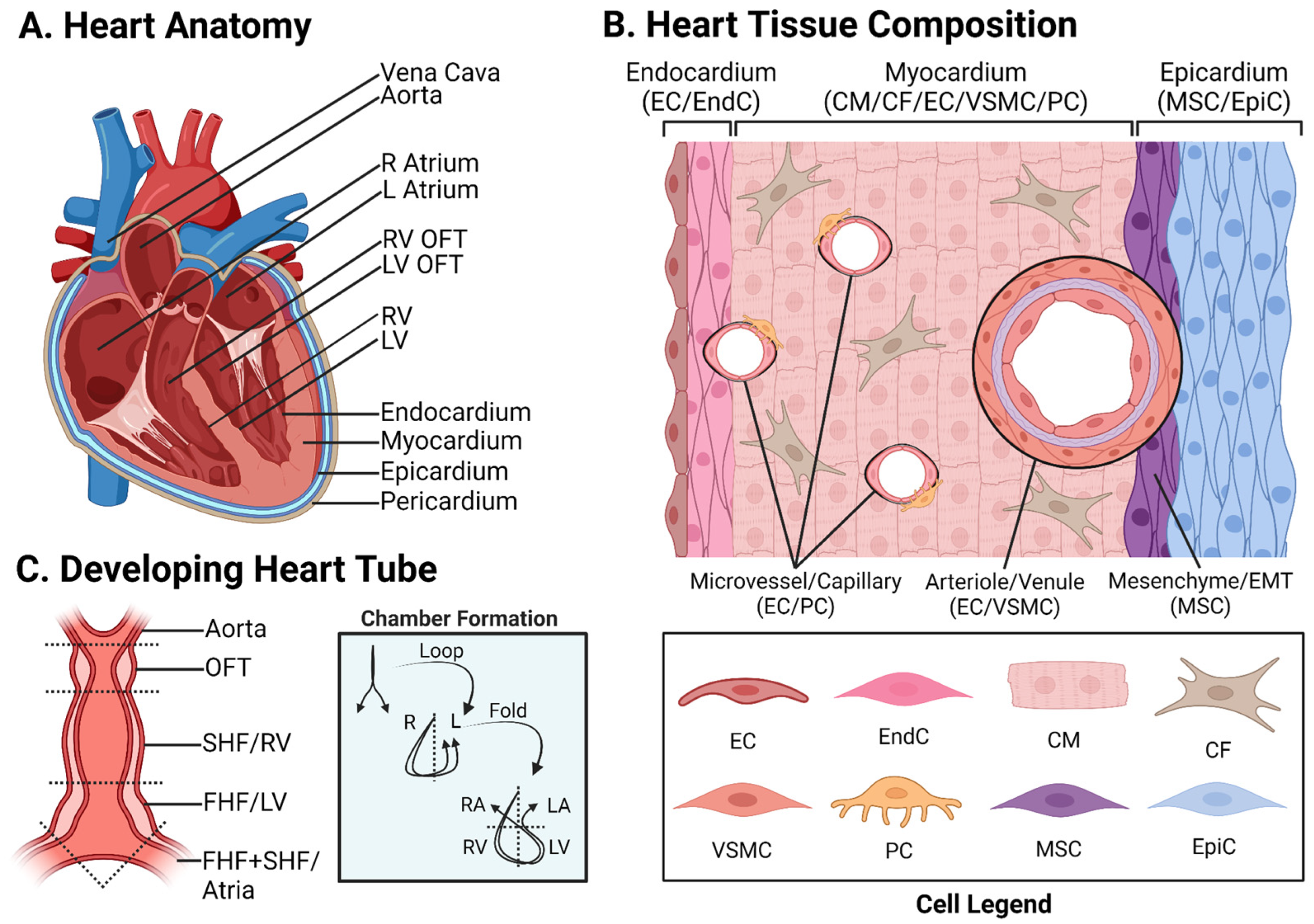
:1. Introduction
2. Constructed Cardiac Tissues (CCTs)
2.1. Engineered Heart Tissues (EHTs)
2.2. Cardiac Organoids (COs)
2.3. Heart-on-a-Chip (HoC)
3. Applications
3.1. Organ Modeling
3.2. Drug Discovery
3.3. Regenerative Medicine
4. Conclusions and Perspective
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations and Acronyms
| 2D | Two dimensional |
| 3D | Three dimensional |
| 3R | Reduce, refine, replace animal testing |
| AP | Action potential |
| AVC | Atrioventricular canal |
| Activin A | Activin group beta, form A dimer |
| BETi | Bromodomain and extraterminal family inhibitor |
| bFGF | Basic fibroblast growth factor, synonymous with FGF-2 |
| BMP-4 | Bone morphogenic protein four |
| BSA-FA | BSA-bound fatty acid |
| BSA | Bovine serum albumin |
| BoC | Body-on-a-chip |
| Cx43 | Connexin 43 gap-junction protein |
| CAD | Computer-assisted design |
| CCT | Constructed cardiac tissue |
| CF | Cardiac fibroblast |
| CHD | Congenital heart disease |
| CHIR | Small molecule Wnt activator |
| CHP | Cardiac homing peptide |
| CM | Cardiomyocyte |
| CMT | Cardiac microtissue |
| CO | Cardiac organoid |
| COVID-19 | Coronavirus disease 2019 |
| CRISPR | Clustered regularly interspaced short palindromic repeats |
| CS | Cardiac spheroid |
| CVD | Cardiovascular disease |
| DCM | Dilated cardiomyopathy |
| dECM | Decellularized ECM |
| Dex | Dexamethasone |
| Dox | Doxorubicin |
| EB | Embryoid body |
| EC | Endothelial cell |
| ECM | Extracellular matrix |
| ECT | Engineered cardiac tissue |
| EHT | Engineered heart tissue |
| EMT | Epithelial-to-mesenchymal transition |
| EndC | Endocardial cell |
| EpiC | Epicardial cell |
| ExT | Exosome therapy |
| FAO | Fatty acid oxidation |
| FDA | United States Food and Drug Administration |
| FHF | First heart field |
| FO | Fusion organoid |
| FRESH | Freeform reversible embedding in suspended hydrogel |
| GelMA | Gelatin conjugated with MA |
| HA | Hyaluronic acid |
| HCM | hypertrophic cardiomyopathy |
| HF | Heart failure |
| HTDS | High-throughput drug screening |
| HoC | Heart-on-a-chip |
| IC | Immune cell |
| ICD | Intercalated disc |
| IFN-y | Interferon-gamma |
| IGF-1 | Insulin-like growth factor one |
| IL-1β | Interleukin one beta |
| IL-6 | Interleukin six |
| iPSC-CCT | iPSC-derived CCT |
| iPSC | Induced pluripotent stem cell |
| IWP2 | Inhibitor of Wnt production two, small molecule Wnt inhibitor |
| IWR | Inhibitor of Wnt response, small molecule Wnt inhibitor |
| JCF | Juxtacardiac field |
| LPS | Lipopolysaccharide |
| LQTS | Long QT syndrome |
| MA-HA | HA conjugated with MA |
| MA | Methacrylate |
| MF | Macrophage |
| MI | Myocardial infarction |
| MMP | Matrix metalloproteinase |
| MSC | Mesenchymal stem cell |
| NO | Nitric oxide |
| OFT | Outflow tract |
| PAm | Polyacrylamide |
| PC | Pericyte |
| PCL | Polycaprolactone |
| PDA | Polydopamine |
| PDGF-BB | Platelet-derived growth factor from B dimer |
| PLGF-2 | Placental growth factor two |
| PDMS | Polydimethylsiloxane |
| PEG-RGD | PEG conjugated with RGD peptide |
| PEG | Polyethylene glycol |
| PGA | Polyglycolic acid |
| PGS | Poly(glycerol sebacate) |
| PINIPAAm | Poly(N-isopropylacrylamide) |
| PLA | Polylactic acid |
| PLGA | Poly(lactic-co-glycolic acid) |
| PMMA | Polymethylmethacrylate |
| PU | Polyurethane |
| PoMaC | Poly(octamethylene maleate (anhydride) citrate) |
| Poly(I:C) | Polyinosinic:polycytidylic acid |
| RGD | Arg-Gly-Asp peptide |
| RGDFK | Arg-Gly-Asp-Phe-Lys peptide |
| SA | Sinoatrial |
| SHF | Second heart field |
| SLA | Stereolithography |
| SQTS | Short QT Syndrome |
| SWIFT | Sacrificial writing into functional tissue |
| T3 | Triiodothyronine |
| TGF-β | Transforming growth factor-beta |
| TNF | Tumor necrosis factor, synonymous with TNF-α |
| VEGF-A | Vascular endothelial growth factor form A |
| VSMC | Vascular smooth muscle cell |
| Wnt-C59 | Small molecule Wnt inhibitor |
| Wnt | Wingless-related integration site |
| YAP | Yes-associated protein |
| YIGSR | Tyr-Ile-Gly-Ser-Arg peptide |
References
- Martin, M.; Gähwiler, E.K.N.; Generali, M.; Hoerstrup, S.P.; Emmert, M.Y. Advances in 3D Organoid Models for Stem Cell-Based Cardiac Regeneration. Int. J. Mol. Sci. 2023, 24, 5188. [Google Scholar] [CrossRef]
- Gisone, I.; Cecchettini, A.; Ceccherini, E.; Persiani, E.; Morales, M.A.; Vozzi, F. Cardiac tissue engineering: Multiple approaches and potential applications. Front. Bioeng. Biotechnol. 2022, 10, 980393. [Google Scholar] [CrossRef] [PubMed]
- Basara, G.; Bahcecioglu, G.; Ozcebe, S.G.; Ellis, B.W.; Ronan, G.; Zorlutuna, P. Myocardial infarction from a tissue engineering and regenerative medicine point of view: A comprehensive review on models and treatments. Biophys. Rev. 2022, 3, 031305. [Google Scholar] [CrossRef] [PubMed]
- Mourad, O.; Yee, R.; Li, M.; Nunes, S.S. Modeling Heart Diseases on a Chip: Advantages and Future Opportunities. Circ. Res. 2023, 132, 483–497. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Bao, X.; Wang, Y.; Lu, T.; Zhang, L. Human induced pluripotent stem cell (hiPSC)-derived cardiomyocyte modelling of cardiovascular diseases for natural compound discovery. Biomed. Pharmacother. 2023, 157, 113970. [Google Scholar] [CrossRef] [PubMed]
- Umbarkar, P.; Ejantkar, S.; Ruiz Ramirez, S.Y.; Toro Cora, A.; Zhang, Q.; Tousif, S.; Lal, H. Cardiac fibroblast GSK-3α aggravates ischemic cardiac injury by promoting fibrosis, inflammation, and impairing angiogenesis. Basic. Res. Cardiol. 2023, 118, 35. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhang, Y.; Wang, J.; Yin, J.; Wang, Z.; Pei, R. Cardiac organoid: Multiple construction approaches and potential applications. J. Mater. Chem. B 2023, 11, 7567–7581. [Google Scholar] [CrossRef] [PubMed]
- Correia, C.D.; Ferreira, A.; Fernandes, M.T.; Silva, B.M.; Esteves, F.; Leitão, H.S.; Bragança, J.; Calado, S.M. Human Stem Cells for Cardiac Disease Modeling and Preclinical and Clinical Applications—Are We on the Road to Success? Cells 2023, 12, 1727. [Google Scholar] [CrossRef] [PubMed]
- Drakhlis, L.; Zweigerdt, R. Heart in a dish—Choosing the right in vitro model. Dis. Model. Mech. 2023, 16, dmm049961. [Google Scholar] [CrossRef]
- Fang, Y.; Sun, W.; Zhang, T.; Xiong, Z. Recent advances on bioengineering approaches for fabrication of functional engineered cardiac pumps: A review. Biomaterials 2022, 280, 121298. [Google Scholar] [CrossRef]
- Wu, T.; Liu, W. Functional hydrogels for the treatment of myocardial infarction. NPG Asia Mater. 2022, 14, 9. [Google Scholar] [CrossRef]
- Margiana, R.; Markov, A.; Zekiy, A.O.; Hamza, M.U.; Al-Dabbagh, K.A.; Al-Zubaidi, S.H.; Hameed, N.M.; Ahmad, I.; Sivaraman, R.; Kzar, H.H.; et al. Clinical application of mesenchymal stem cell in regenerative medicine: A narrative review. Stem Cell Res. Ther. 2022, 13, 366. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Xuan, X.; Hu, J.; Zhang, R.; Jin, H.; Dong, H. How vascular smooth muscle cell phenotype switching contributes to vascular disease. Cell Commun. Signal. 2022, 20, 180. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Liu, K.; Feng, Q.; Liao, Y. Cardiac Organoids: A 3D Technology for Modeling Heart Development and Disease. Stem Cell Rev. Rep. 2022, 18, 2593–2605. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Lei, D.; Yang, Q.; Yang, Y.; Jiang, C.; Shi, H.; Qian, B.; Long, Q.; Chen, W.; Chen, Y.; et al. A perfusable, multifunctional epicardial device improves cardiac function and tissue repair. Nat. Med. 2021, 27, 480–490. [Google Scholar] [CrossRef] [PubMed]
- Richards, D.J.; Li, Y.; Kerr, C.M.; Yao, J.; Beeson, G.C.; Coyle, R.C.; Chen, X.; Jia, J.; Damon, B.; Wilson, R.; et al. Human cardiac organoids for the modelling of myocardial infarction and drug cardiotoxicity. Nat. Biomed. Eng. 2020, 4, 446–462. [Google Scholar] [CrossRef]
- Mills, R.J.; Parker, B.L.; Quaife-Ryan, G.A.; Voges, H.K.; Needham, E.J.; Bornot, A.; Ding, M.; Andersson, H.; Polla, M.; Elliott, D.A.; et al. Drug Screening in Human PSC-Cardiac Organoids Identifies Pro-proliferative Compounds Acting via the Mevalonate Pathway. Cell Stem Cell 2019, 24, 895–907.e6. [Google Scholar] [CrossRef] [PubMed]
- Mills, R.J.; Humphrey, S.J.; Fortuna, P.R.J.; Lor, M.; Foster, S.R.; Quaife-Ryan, G.A.; Johnston, R.L.; Dumenil, T.; Bishop, C.; Rudraraju, R.; et al. BET inhibition blocks inflammation-induced cardiac dysfunction and SARS-CoV-2 infection. Cell 2021, 184, 2167–2182.e22. [Google Scholar] [CrossRef]
- Yang, L.; Han, Y.; Jaffré, F.; Nilsson-Payant, B.E.; Bram, Y.; Wang, P.; Zhu, J.; Zhang, T.; Redmond, D.; Houghton, S.; et al. An Immuno-Cardiac Model for Macrophage-Mediated Inflammation in COVID-19 Hearts. Circ. Res. 2021, 129, 33–46. [Google Scholar] [CrossRef]
- Voges, H.K.; Foster, S.R.; Reynolds, L.; Parker, B.L.; Devilée, L.; Quaife-Ryan, G.A.; Fortuna, P.R.J.; Mathieson, E.; Fitzsimmons, R.; Lor, M.; et al. Vascular cells improve functionality of human cardiac organoids. Cell Rep. 2023, 42, 112322. [Google Scholar] [CrossRef]
- Deir, S.; Mozhdehbakhsh Mofrad, Y.; Mashayekhan, S.; Shamloo, A.; Mansoori-Kermani, A. Step-by-step fabrication of heart-on-chip systems as models for cardiac disease modeling and drug screening. Talanta 2023, 266 Pt 1, 124901. [Google Scholar] [CrossRef]
- Arhontoulis, D.C.; Kerr, C.M.; Richards, D.; Tjen, K.; Hyams, N.; Jones, J.A.; Deleon-Pennell, K.; Menick, D.; Bräuninger, H.; Lindner, D.; et al. Human cardiac organoids to model COVID-19 cytokine storm induced cardiac injuries. J. Tissue Eng. Regen. Med. 2022, 16, 799–811. [Google Scholar] [CrossRef]
- Kim, H.; Kamm, R.D.; Vunjak-Novakovic, G.; Wu, J.C. Progress in multicellular human cardiac organoids for clinical applications. Cell Stem Cell 2022, 29, 503–514. [Google Scholar] [CrossRef]
- Cho, S.; Discher, D.E.; Leong, K.W.; Vunjak-Novakovic, G.; Wu, J.C. Challenges and opportunities for the next generation of cardiovascular tissue engineering. Nat. Methods 2022, 19, 1064–1071. [Google Scholar] [CrossRef]
- Rogozinski, N.; Yanez, A.; Bhoi, R.; Lee, M.Y.; Yang, H. Current methods for fabricating 3D cardiac engineered constructs. iScience 2022, 25, 104330. [Google Scholar] [CrossRef]
- Yao, Y.; Wang, C. Dedifferentiation: Inspiration for devising engineering strategies for regenerative medicine. NPJ Regen. Med. 2020, 5, 14. [Google Scholar] [CrossRef]
- Chi, C.; Song, K. Cellular reprogramming of fibroblasts in heart regeneration. J. Mol. Cell Cardiol. 2023, 180, 84–93. [Google Scholar] [CrossRef]
- St Hilaire, C. Cardiovascular Organoids/3D Models Review Series: An Introduction. Circ. Res. 2023, 132, 481–482. [Google Scholar] [CrossRef]
- Singh, B.N.; Yucel, D.; Garay, B.I.; Tolkacheva, E.G.; Kyba, M.; Perlingeiro, R.C.R.; van Berlo, J.H.; Ogle, B.M. Proliferation and Maturation: Janus and the Art of Cardiac Tissue Engineering. Circ. Res. 2023, 132, 519–540. [Google Scholar] [CrossRef]
- Yang, H.; Yang, Y.; Kiskin, F.N.; Shen, M.; Zhang, J.Z. Recent advances in regulating the proliferation or maturation of human-induced pluripotent stem cell-derived cardiomyocytes. Stem Cell Res. Ther. 2023, 14, 228. [Google Scholar] [CrossRef]
- Guo, Y.; Pu, W.T. Cardiomyocyte Maturation: New Phase in Development. Circ. Res. 2020, 126, 1086–1106. [Google Scholar] [CrossRef]
- Zhang, H.; Lui, K.O.; Zhou, B. Endocardial Cell Plasticity in Cardiac Development, Diseases and Regeneration. Circ. Res. 2018, 122, 774–789. [Google Scholar] [CrossRef]
- Vandergriff, A.; Huang, K.; Shen, D.; Hu, S.; Hensley, M.T.; Caranasos, T.G.; Qian, L.; Cheng, K. Targeting regenerative exosomes to myocardial infarction using cardiac homing peptide. Theranostics 2018, 8, 1869–1878. [Google Scholar] [CrossRef]
- Liu, C.; Feng, X.; Li, G.; Gokulnath, P.; Xiao, J. Generating 3D human cardiac constructs from pluripotent stem cells. EBioMedicine 2022, 76, 103813. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Wang, L.; Wei, Y.; Krishnamurthy, P.; Walcott, G.P.; Menasché, P.; Zhang, J. Exosomes secreted by hiPSC-derived cardiac cells improve recovery from myocardial infarction in swine. Sci. Transl. Med. 2020, 12, eaay1318, Erratum in Sci. Transl. Med. 2020, 12, eabf1250.. [Google Scholar] [CrossRef]
- Zhuang, R.Z.; Lock, R.; Liu, B.; Vunjak-Novakovic, G. Opportunities and challenges in cardiac tissue engineering from an analysis of two decades of advances. Nat. Biomed. Eng. 2022, 6, 327–338. [Google Scholar] [CrossRef]
- Yu, B.; Zhao, S.R.; Yan, C.D.; Zhang, M.; Wu, J.C. Deconvoluting the Cells of the Human Heart with iPSC Technology: Cell Types, Protocols, and Uses. Curr. Cardiol. Rep. 2022, 24, 487–496. [Google Scholar] [CrossRef]
- Tan, Y.; Coyle, R.C.; Barrs, R.W.; Silver, S.E.; Li, M.; Richards, D.J.; Lin, Y.; Jiang, Y.; Wang, H.; Menick, D.R.; et al. Nanowired human cardiac organoid transplantation enables highly efficient and effective recovery of infarcted hearts. Sci. Adv. 2023, 9, eadf2898. [Google Scholar] [CrossRef] [PubMed]
- Shoji, J.Y.; Davis, R.P.; Mummery, C.L.; Krauss, S. Global Meta-Analysis of Organoid and Organ-on-Chip Research. Adv. Healthc. Mater. 2023, e2301067. [Google Scholar] [CrossRef]
- Meng, X.; Yao, D.; Imaizumi, K.; Chen, X.; Kelley, K.W.; Reis, N.; Thete, M.V.; Arjun McKinney, A.; Kulkarni, S.; Panagiotakos, G.; et al. Assembloid CRISPR screens reveal impact of disease genes in human neurodevelopment. Nature 2023, 622, 359–366. [Google Scholar] [CrossRef]
- Giraud, M.N.; Ahmed, S.; Ullrich, N.D. Human Stem Cell-Derived Cardiac Organoid-Like Structures: Generation and Applications. In Cardiovascular Applications of Stem Cells; Haider, K.H., Ed.; Springer: Singapore, 2023. [Google Scholar] [CrossRef]
- Gribaudo, S.; Robert, R.; van Sambeek, B.; Mirdass, C.; Lyubimova, A.; Bouhali, K.; Ferent, J.; Morin, X.; van Oudenaarden, A.; Nedelec, S. Self-organizing models of human trunk organogenesis recapitulate spinal cord and spine co-morphogenesis. Nat. Biotechnol. 2023. Online ahead of print. [Google Scholar] [CrossRef]
- Tenreiro, M.F.; Branco, M.A.; Cotovio, J.P.; Cabral, J.M.S.; Fernandes, T.G.; Diogo, M.M. Advancing organoid design through co-emergence, assembly, and bioengineering. Trends Biotechnol. 2023, 41, 923–938. [Google Scholar] [CrossRef]
- Vandana, J.J.; Manrique, C.; Lacko, L.A.; Chen, S. Human pluripotent-stem-cell-derived organoids for drug discovery and evaluation. Cell Stem Cell 2023, 30, 571–591. [Google Scholar] [CrossRef]
- Thomas, D.; Choi, S.; Alamana, C.; Parker, K.K.; Wu, J.C. Cellular and Engineered Organoids for Cardiovascular Models. Circ. Res. 2022, 130, 1780–1802. [Google Scholar] [CrossRef]
- Cho, J.; Lee, H.; Rah, W.; Chang, H.J.; Yoon, Y.S. From engineered heart tissue to cardiac organoid. Theranostics 2022, 12, 2758–2772. [Google Scholar] [CrossRef] [PubMed]
- Lewis-Israeli, Y.R.; Wasserman, A.H.; Gabalski, M.A.; Volmert, B.D.; Ming, Y.; Ball, K.A.; Yang, W.; Zou, J.; Ni, G.; Pajares, N.; et al. Self-assembling human heart organoids for the modeling of cardiac development and congenital heart disease. Nat. Commun. 2021, 12, 5142. [Google Scholar] [CrossRef] [PubMed]
- Yip, S.; Wang, N.; Sugimura, R. Give them vasculature and immune cells—How to fill the gap of organoids. Cells Tissues Organs. 2023, 212, 369–382. [Google Scholar] [CrossRef]
- Xu, R.; Boreland, A.J.; Li, X.; Erickson, C.; Jin, M.; Atkins, C.; Pang, Z.P.; Daniels, B.P.; Jiang, P. Developing human pluripotent stem cell-based cerebral organoids with a controllable microglia ratio for modeling brain development and pathology. Stem Cell Rep. 2021, 16, 1923–1937. [Google Scholar] [CrossRef] [PubMed]
- Patsch, C.; Challet-Meylan, L.; Thoma, E.C.; Urich, E.; Heckel, T.; O’Sullivan, J.F.; Grainger, S.J.; Kapp, F.G.; Sun, L.; Christensen, K.; et al. Generation of vascular endothelial and smooth muscle cells from human pluripotent stem cells. Nat. Cell Biol. 2015, 17, 994–1003. [Google Scholar] [CrossRef]
- Drakhlis, L.; Biswanath, S.; Farr, C.M.; Lupanow, V.; Teske, J.; Ritzenhoff, K.; Franke, A.; Manstein, F.; Bolesani, E.; Kempf, H.; et al. Human heart-forming organoids recapitulate early heart and foregut development. Nat. Biotechnol. 2021, 39, 737–746, Erratum in Nat. Biotechnol. 2021, 39, 737–746. [Google Scholar] [CrossRef]
- Branco, M.A.; Dias, T.P.; Cabral, J.M.S.; Pinto-do-Ó, P.; Diogo, M.M. Human multilineage pro-epicardium/foregut organoids support the development of an epicardium/myocardium organoid. Nat. Commun. 2022, 13, 6981. [Google Scholar] [CrossRef]
- Feyen, D.A.M.; McKeithan, W.L.; Bruyneel, A.A.N.; Spiering, S.; Hörmann, L.; Ulmer, B.; Zhang, H.; Briganti, F.; Schweizer, M.; Hegyi, B.; et al. Metabolic Maturation Media Improve Physiological Function of Human iPSC-Derived Cardiomyocytes. Cell Rep. 2020, 32, 107925. [Google Scholar] [CrossRef]
- Campostrini, G.; Meraviglia, V.; Giacomelli, E.; van Helden, R.W.J.; Yiangou, L.; Davis, R.P.; Bellin, M.; Orlova, V.V.; Mummery, C.L. Generation, functional analysis and applications of isogenic three-dimensional self-aggregating cardiac microtissues from human pluripotent stem cells. Nat. Protoc. 2021, 16, 2213–2256. [Google Scholar] [CrossRef]
- Andersen, P.; Tampakakis, E.; Jimenez, D.V.; Kannan, S.; Miyamoto, M.; Shin, H.K.; Saberi, A.; Murphy, S.; Sulistio, E.; Chelko, S.P.; et al. Precardiac organoids form two heart fields via Bmp/Wnt signaling. Nat. Commun. 2018, 9, 3140. [Google Scholar] [CrossRef] [PubMed]
- Hofbauer, P.; Jahnel, S.M.; Papai, N.; Giesshammer, M.; Deyett, A.; Schmidt, C.; Penc, M.; Tavernini, K.; Grdseloff, N.; Meledeth, C.; et al. Cardioids reveal self-organizing principles of human cardiogenesis. Cell 2021, 184, 3299–3317.e22. [Google Scholar] [CrossRef]
- Sahara, M. Recent Advances in Generation of In Vitro Cardiac Organoids. Int. J. Mol. Sci. 2023, 24, 6244. [Google Scholar] [CrossRef]
- Hong, Y.; Zhao, Y.; Li, H.; Yang, Y.; Chen, M.; Wang, X.; Luo, M.; Wang, K. Engineering the maturation of stem cell-derived cardiomyocytes. Front. Bioeng. Biotechnol. 2023, 11, 1155052. [Google Scholar] [CrossRef]
- Feng, W.; Schriever, H.; Jiang, S.; Bais, A.; Wu, H.; Kostka, D.; Li, G. Computational profiling of hiPSC-derived heart organoids reveals chamber defects associated with NKX2-5 deficiency. Commun. Biol. 2022, 5, 399. [Google Scholar] [CrossRef]
- Schmidt, C.; Deyett, A.; Ilmer, T.; Haendeler, S.; Torres Caballero, A.; Novatchkova, M.; Netzer, M.A.; Ceci Ginistrelli, L.; Mancheno Juncosa, E.; Bhattacharya, T.; et al. Multi-chamber cardioids unravel human heart development and cardiac defects. Cell 2023, 186, 5587–5605.e27. [Google Scholar] [CrossRef]
- Trimm, E.; Red-Horse, K. Vascular endothelial cell development and diversity. Nat. Rev. Cardiol. 2023, 20, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Pinto, A.R.; Ilinykh, A.; Ivey, M.J.; Kuwabara, J.T.; D’Antoni, M.L.; Debuque, R.; Chandran, A.; Wang, L.; Arora, K.; Rosenthal, N.A.; et al. Revisiting Cardiac Cellular Composition. Circ. Res. 2016, 118, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Meier, A.B.; Zawada, D.; De Angelis, M.T.; Martens, L.D.; Santamaria, G.; Zengerle, S.; Nowak-Imialek, M.; Kornherr, J.; Zhang, F.; Tian, Q.; et al. Epicardioid single-cell genomics uncovers principles of human epicardium biology in heart development and disease. Nat. Biotechnol. 2023, 41, 1787–1800. [Google Scholar] [CrossRef]
- Muhl, L.; Genové, G.; Leptidis, S.; Liu, J.; He, L.; Mocci, G.; Sun, Y.; Gustafsson, S.; Buyandelger, B.; Chivukula, I.V.; et al. Single-cell analysis uncovers fibroblast heterogeneity and criteria for fibroblast and mural cell identification and discrimination. Nat. Commun. 2020, 11, 3953, Erratum in Nat. Commun. 2020, 11, 4493. [Google Scholar] [CrossRef]
- Strobel, H.A.; Moss, S.M.; Hoying, J.B. Vascularized Tissue Organoids. Bioengineering 2023, 10, 124. [Google Scholar] [CrossRef]
- Joddar, B.; Natividad-Diaz, S.L.; Padilla, A.E.; Esparza, A.A.; Ramirez, S.P.; Chambers, D.R.; Ibaroudene, H. Engineering approaches for cardiac organoid formation and their characterization. Transl. Res. 2022, 250, 46–67. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.G.; Kim, Y.J.; Son, M.Y.; Oh, M.S.; Kim, J.; Ryu, B.; Kang, K.R.; Baek, J.; Chung, G.; Woo, D.H.; et al. Generation of human iPSCs derived heart organoids structurally and functionally similar to heart. Biomaterials 2022, 290, 121860. [Google Scholar] [CrossRef]
- Mousavi, A.; Stefanek, E.; Jafari, A.; Ajji, Z.; Naghieh, S.; Akbari, M.; Savoji, H. Tissue-engineered heart chambers as a platform technology for drug discovery and disease modeling. Biomater. Adv. 2022, 138, 212916. [Google Scholar] [CrossRef]
- Ergir, E.; Oliver-De La Cruz, J.; Fernandes, S.; Cassani, M.; Niro, F.; Pereira-Sousa, D.; Vrbský, J.; Vinarský, V.; Perestrelo, A.R.; Debellis, D.; et al. Generation and maturation of human iPSC-derived 3D organotypic cardiac microtissues in long-term culture. Sci. Rep. 2022, 12, 17409. [Google Scholar] [CrossRef]
- Ng, W.H.; Varghese, B.; Jia, H.; Ren, X. Alliance of Heart and Endoderm: Multilineage Organoids to Model Co-development. Circ. Res. 2023, 132, 511–518. [Google Scholar] [CrossRef]
- Silva, A.C.; Matthys, O.B.; Joy, D.A.; Kauss, M.A.; Natarajan, V.; Lai, M.H.; Turaga, D.; Blair, A.P.; Alexanian, M.; Bruneau, B.G.; et al. Co-emergence of cardiac and gut tissues promotes cardiomyocyte maturation within human iPSC-derived organoids. Cell Stem Cell 2021, 28, 2137–2152.e6. [Google Scholar] [CrossRef]
- Olmsted, Z.T.; Paluh, J.L. A combined human gastruloid model of cardiogenesis and neurogenesis. iScience 2022, 25, 104486. [Google Scholar] [CrossRef] [PubMed]
- Rossi, G.; Broguiere, N.; Miyamoto, M.; Boni, A.; Guiet, R.; Girgin, M.; Kelly, R.G.; Kwon, C.; Lutolf, M.P. Capturing Cardiogenesis in Gastruloids. Cell Stem Cell 2021, 28, 230–240.e6. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Rafatian, N.; Feric, N.T.; Cox, B.J.; Aschar-Sobbi, R.; Wang, E.Y.; Aggarwal, P.; Zhang, B.; Conant, G.; Ronaldson-Bouchard, K.; et al. A Platform for Generation of Chamber-Specific Cardiac Tissues and Disease Modeling. Cell 2019, 176, 913–927.e18. [Google Scholar] [CrossRef] [PubMed]
- Fang, G.; Chen, Y.; Lu, H.; Jin, D. Advances in spheroids and organoids on a chip. Adv. Funct. Mater. 2023, 33, 2215043. [Google Scholar] [CrossRef]
- Häneke, T.; Sahara, M. Progress in Bioengineering Strategies for Heart Regenerative Medicine. Int. J. Mol. Sci. 2022, 23, 3482. [Google Scholar] [CrossRef] [PubMed]
- Roh, T.T.; Chen, Y.; Paul, H.T.; Guo, C.; Kaplan, D.L. 3D bioengineered tissue model of the large intestine to study inflammatory bowel disease. Biomaterials 2019, 225, 119517. [Google Scholar] [CrossRef]
- Zhu, D.; Li, Z.; Huang, K.; Caranasos, T.G.; Rossi, J.S.; Cheng, K. Minimally invasive delivery of therapeutic agents by hydrogel injection into the pericardial cavity for cardiac repair. Nat. Commun. 2021, 12, 1412. [Google Scholar] [CrossRef]
- Knight, W.E.; Cao, Y.; Lin, Y.H.; Chi, C.; Bai, B.; Sparagna, G.C.; Zhao, Y.; Du, Y.; Londono, P.; Reisz, J.A.; et al. Maturation of Pluripotent Stem Cell-Derived Cardiomyocytes Enables Modeling of Human Hypertrophic Cardiomyopathy. Stem Cell Rep. 2021, 16, 519–533. [Google Scholar] [CrossRef]
- Tenreiro, M.F.; Louro, A.F.; Alves, P.M.; Serra, M. Next generation of heart regenerative therapies: Progress and promise of cardiac tissue engineering. NPJ Regen. Med. 2021, 6, 30. [Google Scholar] [CrossRef] [PubMed]
- Ronaldson-Bouchard, K.; Ma, S.P.; Yeager, K.; Chen, T.; Song, L.; Sirabella, D.; Morikawa, K.; Teles, D.; Yazawa, M.; Vunjak-Novakovic, G. Advanced maturation of human cardiac tissue grown from pluripotent stem cells. Nature 2018, 556, 239–243, Erratum in Nature 2019, 572, E16–E17. [Google Scholar] [CrossRef] [PubMed]
- Parikh, S.S.; Blackwell, D.J.; Gomez-Hurtado, N.; Frisk, M.; Wang, L.; Kim, K.; Dahl, C.P.; Fiane, A.; Tønnessen, T.; Kryshtal, D.O.; et al. Thyroid and Glucocorticoid Hormones Promote Functional T-Tubule Development in Human-Induced Pluripotent Stem Cell-Derived Cardiomyocytes. Circ. Res. 2017, 121, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Lebreton, F.; Lavallard, V.; Bellofatto, K.; Bonnet, R.; Wassmer, C.H.; Perez, L.; Kalandadze, V.; Follenzi, A.; Boulvain, M.; Kerr-Conte, J.; et al. Insulin-producing organoids engineered from islet and amniotic epithelial cells to treat diabetes. Nat. Commun. 2019, 10, 4491, Erratum in Nat. Commun. 2020, 11, 5159. [Google Scholar] [CrossRef] [PubMed]
- Haenseler, W.; Sansom, S.N.; Buchrieser, J.; Newey, S.E.; Moore, C.S.; Nicholls, F.J.; Chintawar, S.; Schnell, C.; Antel, J.P.; Allen, N.D.; et al. A Highly Efficient Human Pluripotent Stem Cell Microglia Model Displays a Neuronal-Co-culture-Specific Expression Profile and Inflammatory Response. Stem Cell Rep. 2017, 8, 1727–1742. [Google Scholar] [CrossRef] [PubMed]
- Rebelo, S.P.; Pinto, C.; Martins, T.R.; Harrer, N.; Estrada, M.F.; Loza-Alvarez, P.; Cabeçadas, J.; Alves, P.M.; Gualda, E.J.; Sommergruber, W.; et al. 3D-3-culture: A tool to unveil macrophage plasticity in the tumour microenvironment. Biomaterials 2018, 163, 185–197. [Google Scholar] [CrossRef]
- Liang, P.Y.; Chang, Y.; Jin, G.; Lian, X.; Bao, X. Wnt signaling directs human pluripotent stem cells into vascularized cardiac organoids with chamber-like structures. Front. Bioeng. Biotechnol. 2022, 10, 1059243. [Google Scholar] [CrossRef]
- Salewskij, K.; Penninger, J.M. Blood Vessel Organoids for Development and Disease. Circ. Res. 2023, 132, 498–510. [Google Scholar] [CrossRef]
- Faustino Martins, J.M.; Fischer, C.; Urzi, A.; Vidal, R.; Kunz, S.; Ruffault, P.L.; Kabuss, L.; Hube, I.; Gazzerro, E.; Birchmeier, C.; et al. Self-Organizing 3D Human Trunk Neuromuscular Organoids. Cell Stem Cell 2020, 26, 172–186.e6, Erratum in Cell Stem Cell 2020, 27, 498. [Google Scholar] [CrossRef]
- Embryonic and Induced Pluripotent Stem Cell Differentiation Pathways & Lineage-Specific Markers. Available online: https://www.rndsystems.com/pathways/embryonic-induced-pluripotent-stem-cell-differentiation-pathways-lineage-specific-markers (accessed on 4 December 2023).
- Unagolla, J.M.; Jayasuriya, A.C. Recent advances in organoid engineering: A comprehensive review. Appl. Mater. Today 2022, 29, 101582. [Google Scholar] [CrossRef]
- Gallet, R.; Dawkins, J.; Valle, J.; Simsolo, E.; de Couto, G.; Middleton, R.; Tseliou, E.; Luthringer, D.; Kreke, M.; Smith, R.R.; et al. Exosomes secreted by cardiosphere-derived cells reduce scarring, attenuate adverse remodelling, and improve function in acute and chronic porcine myocardial infarction. Eur. Heart J. 2017, 38, 201–211. [Google Scholar] [CrossRef]
- Antes, T.J.; Middleton, R.C.; Luther, K.M.; Ijichi, T.; Peck, K.A.; Liu, W.J.; Valle, J.; Echavez, A.K.; Marbán, E. Targeting extracellular vesicles to injured tissue using membrane cloaking and surface display. J. Nanobiotechnol. 2018, 16, 61. [Google Scholar] [CrossRef]
- Cheng, G.; Zhu, D.; Huang, K.; Caranasos, T.G. Minimally invasive delivery of a hydrogel-based exosome patch to prevent heart failure. J. Mol. Cell Cardiol. 2022, 169, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Peres Moreno Maia-Joca, R.; Ong, C.S.; Wilson, I.; DiSilvestre, D.; Tomaselli, G.F.; Reich, D.H. Enhancement of human iPSC-derived cardiomyocyte maturation by chemical conditioning in a 3D environment. J. Mol. Cell. Cardiol. 2020, 138, 1–11. [Google Scholar] [CrossRef]
- Ng, W.H.; Johnston, E.K.; Tan, J.J.; Bliley, J.M.; Feinberg, A.W.; Stolz, D.B.; Sun, M.; Wijesekara, P.; Hawkins, F.; Kotton, D.N.; et al. Recapitulating human cardio-pulmonary co-development using simultaneous multilineage differentiation of pluripotent stem cells. eLife 2022, 11, e67872. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.R.; Zhang, Y.S.; Kim, D.J.; Manbohi, A.; Avci, H.; Silvestri, A.; Aleman, J.; Hu, N.; Kilic, T.; Keung, W.; et al. Aptamer-Based Microfluidic Electrochemical Biosensor for Monitoring Cell-Secreted Trace Cardiac Biomarkers. Anal. Chem. 2016, 88, 10019–10027. [Google Scholar] [CrossRef] [PubMed]
- Yin, F.; Zhang, X.; Wang, L.; Wang, Y.; Zhu, Y.; Li, Z.; Tao, T.; Chen, W.; Yu, H.; Qin, J. HiPSC-derived multi-organoids-on-chip system for safety assessment of antidepressant drugs. Lab Chip 2021, 21, 571–581. [Google Scholar] [CrossRef]
- White, M.J.V.; Briquez, P.S.; White, D.A.V.; Hubbell, J.A. VEGF-A, PDGF-BB and HB-EGF engineered for promiscuous super affinity to the extracellular matrix improve wound healing in a model of type 1 diabetes. NPJ Regen. Med. 2021, 6, 76. [Google Scholar] [CrossRef]
- Adelipour, M.; Lubman, D.M.; Kim, J. Potential applications of mesenchymal stem cells and their derived exosomes in regenerative medicine. Expert. Opin. Biol. Ther. 2023, 23, 491–507. [Google Scholar] [CrossRef]
- Cao, D.J. Macrophages in Cardiovascular Homeostasis and Disease. Circulation 2018, 138, 2452–2455. [Google Scholar] [CrossRef]
- Chi, C.; Knight, W.E.; Riching, A.S.; Zhang, Z.; Tatavosian, R.; Zhuang, Y.; Moldovan, R.; Rachubinski, A.L.; Gao, D.; Xu, H.; et al. Interferon hyperactivity impairs cardiogenesis in Down syndrome via downregulation of canonical Wnt signaling. iScience 2023, 26, 107012. [Google Scholar] [CrossRef]
- Roland, T.J.; Strauss, G.L.; Bushra, N.; Muschol, M.; Koria, P. Isoelectric point (pI)-based phase separation (pI-BPS) purification of elastin-like polypeptides (ELPs) containing charged, biologically active fusion proteins (ELP-FPs). Biotechnol. Prog. 2023, 39, e3381. [Google Scholar] [CrossRef]
- Hu, S.; Wang, X.; Li, Z.; Zhu, D.; Cores, J.; Wang, Z.; Li, J.; Mei, X.; Cheng, X.; Su, T.; et al. Platelet membrane and stem cell exosome hybrid enhances cellular uptake and targeting to heart injury. Nano Today 2021, 39, 101210. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| iPSC-CCT Platform | Author | Pros | Cons | Architecture | Vascularization | Maturation |
|---|---|---|---|---|---|---|
| Cardiac Spheroid | Arhontoulis et al., 2022 [22] | Crosstalk between multiple cell types captures inflammatory signaling Sufficient throughput for effective drug screening | Sequential introduction of cell lines limits the emergence of structures | Aggregate of iPSC-derived cardiomyocytes, and established cardiac fibroblast, vascular stem cell, and mesenchymal stem cell lines | Inclusion of vascular stem cells, while not itself forming vasculature, nonetheless captures physiological responses to injury and infection like endothelial damage | Crosstalk between cell types upregulates adrenergic signaling, driving maturation, though culture periods were less than a week, limiting observed maturation |
| Cardiac Spheroid to Organoid | Tan et al., 2023 [38] | Highly biocompatible given the presence of multiple cell types Perfusable, host vascular integration upon transplant in infarcted, immunodeficient mice Nanowires improve host electrical integration | Limited structural complexity | Organized aggregate of iPSC-derived cardiomyocytes, human cardiac fibroblasts, and established vascular stem cell lines, with optional nanowires | Inclusion of vascular stem cells and continued culturing primes the spheroid/organoid to promote angiogenesis at the infarcted site of transplantation | Crosstalk between cell types upregulates adrenergic signaling, driving maturation within the spheroid/organoid and with host tissue |
| Cardiac Organoid | Lewis-Israeli et al., 2021 [47] | Self-assembles into all heart layers Epicardial spreading and microvascular network formation Multiple chambers per organoid Relatively inexpensive with few growth factors used | Batch-to-batch variability Difficult to apply electromechanical stimuli to batches | Microvascularized, chambered, three-layered heart miniature organ grown from iPSC aggregate | Produces microvasculature with minimal growth factors via Wnt reactivation for epicardial spreading and microvessel networks | Metabolic maturation by supplementing culture media with fatty acids to promote fatty acid oxidation in cardiomyocytes after about a week in culture |
| Cardiac Pulmonary Organoid | W.H. Ng et al., 2022 [95] | Captures inter=germ-layer tissue cooperation Relatively inexpensive with few growth factors used Self-directed spatial sorting of tissues from different germ lineages | Batch-to-batch variability Though iPSCs are co-differentiated, they are initially in 2D cultures, which restricts early organization | Spatially distinct heart and lung miniature organ system grown from iPSC aggregate | Angiogenic factors added early in culture but used for mesoderm and endoderm specification, thus lacking vasculature formation | Crosstalk between germ layers promotes mutual development via dual Wnt and TGF-β signaling modulation followed by retinoic acid and glucocorticoid treatment |
| Cardiac Foregut Organoid | Silva et al., 2021 [71] | Captures inter-germ-layer tissue cooperation Epicardial spreading and microvasculature Tissue lifespan longer than a year | Batch-to-batch variability Lack of chamber-like morphology | Beating myocardial core surrounded by non-beating epicardial layer grown from iPSC aggregate | Epicardial-permissive media with ascorbic acid after about a week in culture allows microvessels to emerge with epicardial-derived vascular cells | Crosstalk between germ layers promotes mutual development, and prolonged culture of over a year enables maturation into steady-state, chamber-specific cardiomyocytes |
| Cardiac Assembloid | Schmidt et al., 2023 [60] | Heart field and chamber-specific organoids Action potential propagates from atrial organoid to left and then right ventricle Flexible model; can further study individual chamber-specific organoids Low batch-to-batch variation for an organoid platform | Requires customized 3D printing of molds with silicone casting to properly align the three organoids into a linear heart-tube-like structure Absence of epicardium and microvasculature Relatively expensive with many growth factors used | Atrial, left ventricular, and right ventricular organoids use linearly inside a mold into a heart tube-like structure grown from separate iPSC aggregates | Angiogenic factor supplementation improves endocardial and endothelial emergence but without significant vasculature formation | Chamber-specific or combined media for cardiomyocyte maturation via fatty acid metabolism, hormones, and glucocorticoids |
| Engineered Cardiac Organoid | Mills et al., 2019, 2021 [17,18] | Self-directed formation of structure and microvessels Biologically complex for an engineered tissue Macroscale 3D structure for physiological drug response Sufficient throughput for effective drug screening | Requires specialized, expensive machinery Lack of chamber-like morphology | iPSC-derived cardiomyocytes and iPSC-derived stromal cells of epicardial lineage densely suspended in hydrogel | Inclusion of iPSC-derived vascular cells enables microvascular formation in an isogenic platform | Suspension of organoids in engineered heart tissue stretcher allows for contractile stimulation alongside eventual fatty acid supplementation |
| Engineered Heart Tissue | Ronaldson-Bouchard et al., 2018 [81] | Exceptional electrophysiological, metabolic, and mechanical maturation markers Mature cardiomyocytes have good drug sensitivity Ramped, non-uniform stimulation improves maturation | Requires several specialized machines Limited biological complexity with no epicardium, endocardium, or cardiac fibroblasts | Mechanically suspended cardiomyocyte and cardiac fibroblast seeded in a fibrin hydrogel | Angiogenic factors added early in culture but used for cardiac specification, thus lacking vasculature formation | Gradually increasing intensity of mechanical stretch and electrical stimulation leads to robust, anisotropic/elongated cardiomyocytes |
| Heart-on-a-Chip | Shin et al., 2016 [96] | In-built microelectrodes for sensing and data collection Infusion system for tested drugs Detection of molecules indicating drug cardiotoxicity Using microscale fluid volumes improves cost efficiency | Sealing and sterility in microfluidics is difficult Dedicated electronic systems required Only a single cell type dramatically limits physiological relevance | Microfluidic bioreactor culture of cardiac spheroids composed of embryonic stem cell-derived cardiomyocytes | Limited biological complexity does not enable vasculature formation | Difficulty maintaining microfluidic culture does not enable significant cardiomyocyte maturation |
| Heart-and-Liver-on-a-Chip | F. Yin, et al., 2021 [97] | Captures key drug interactions through the larger circulatory system Reveals drug effects as other tissues process the drug into downstream metabolites Accommodates large sample size for sensitive detection of analytes | Several custom parts required for novel co-culture system Lack of in-built sensors necessitates manual media extraction for analysis of analytes | Several iPSC-derived cardiac and hepatic miniature organs held in place with micropillars and separated by a permeable membrane in microfluidics systems | Though not possessing vascular cells, macroscopic vascular relationships are modeled | Hormonal treatment after about three weeks of culture during hepatic tissue integration and cardiac maturation |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roland, T.J.; Song, K. Advances in the Generation of Constructed Cardiac Tissue Derived from Induced Pluripotent Stem Cells for Disease Modeling and Therapeutic Discovery. Cells 2024, 13, 250. https://doi.org/10.3390/cells13030250
Roland TJ, Song K. Advances in the Generation of Constructed Cardiac Tissue Derived from Induced Pluripotent Stem Cells for Disease Modeling and Therapeutic Discovery. Cells. 2024; 13(3):250. https://doi.org/10.3390/cells13030250
Chicago/Turabian StyleRoland, Truman J., and Kunhua Song. 2024. "Advances in the Generation of Constructed Cardiac Tissue Derived from Induced Pluripotent Stem Cells for Disease Modeling and Therapeutic Discovery" Cells 13, no. 3: 250. https://doi.org/10.3390/cells13030250