
New Insights on the Male and Female Reproductive Organs of Centrorhynchus globocaudatus (Acanthocephala), Intestinal Parasite of Birds of Prey



Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Birds and Sampling
2.2. Light and Transmission Electron Microscopy (TEM)
3. Results
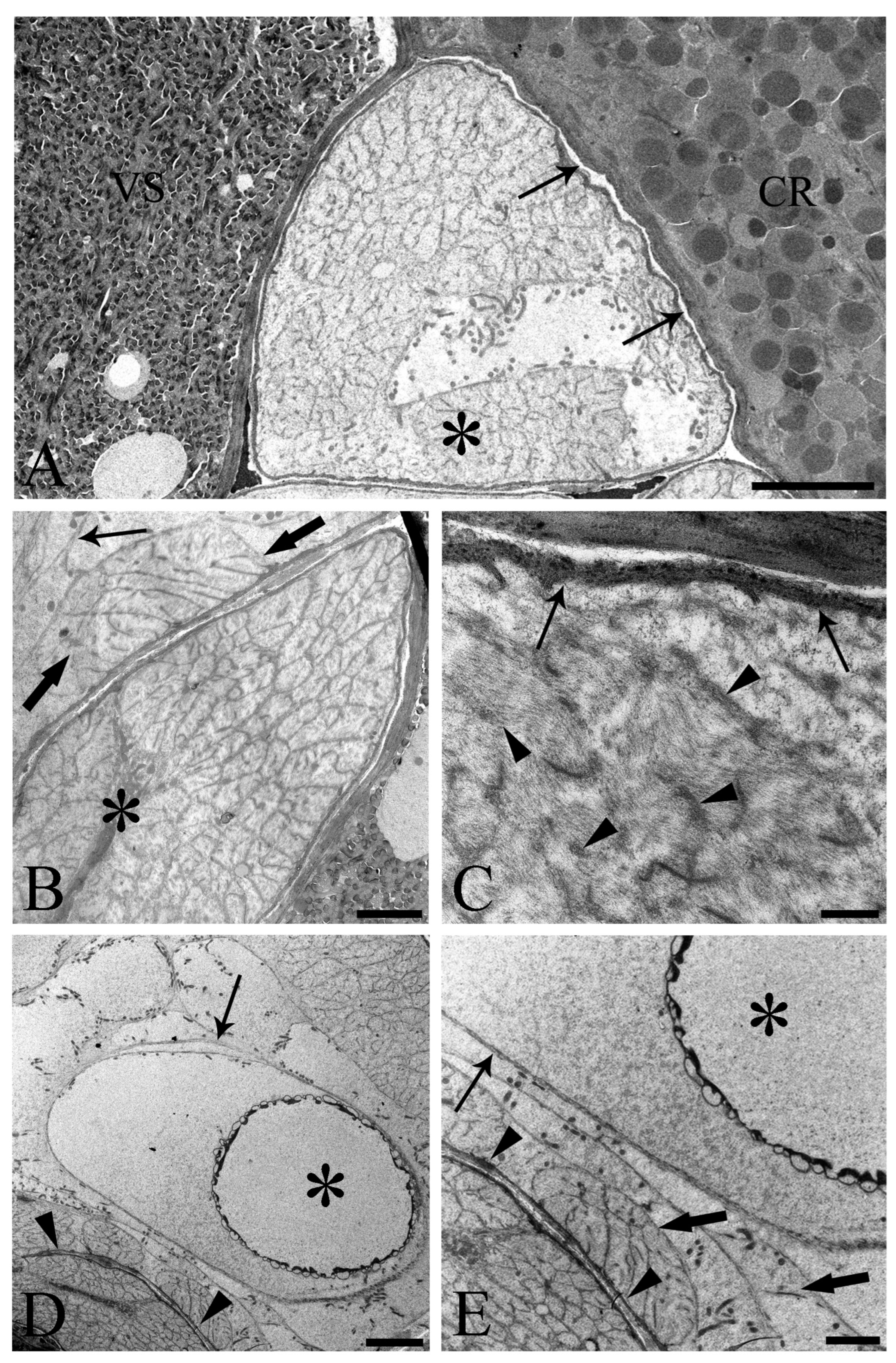
3.1. Light Microscopy
3.2. Transmission Electron Microscopy (TEM)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smales, L.R. Acanthocephala. In Handbook of Zoology. Gastrotricha, Cycloneuralia, and Gnathifera; Schmidt-Rhaesa, A., Ed.; De Gruyter: Berlin, Germany, 2015; Volume 3, pp. 317–336. ISBN 978-3-11-027381-6. [Google Scholar]
- Crompton, D.W.T.; Whitfield, P.J. Observations on the Functional Organization of the Ovarian Balls of Moniliformis and Polymorphus (Acanthocephala). Parasitology 1974, 69, 429–443. [Google Scholar] [CrossRef]
- Herlyn, H. Thorny-Headed Worms (Acanthocephala): Jaw-Less Members of Jaw-Bearing Worms That Parasitize Jawed Arthropods and Jawed Vertebrates. In The Evolution and Fossil Record of Parasitism; De Baets, K., Huntley, J.W., Eds.; Topics in Geobiology; Springer International Publishing: Cham, Switzerland, 2021; Volume 49, pp. 273–313. ISBN 978-3-030-42483-1. [Google Scholar]
- Sayyaf Dezfuli, B.; Giari, L. Acanthocephala. In Invertebrate Zoology—A Tree of Life Approach; Schierwater, B., DeSalle, R., Eds.; CRC Press: New York, NY, USA, 2022; pp. 369–378. ISBN 978-1-4822-3582-1. [Google Scholar]
- Bullock, W.L. Morphological Features as Tools and as Pitfalls in Acanthocephalan Systematics. In Problems in Systematics of Parasites; Schmidt, G.D., Ed.; University Park Press: Baltimore, MD, USA, 1969; pp. 9–45. [Google Scholar]
- Crompton, D.W.T.; Arnold, S.; Walters, D.E. The Number and Size of Ovarian Balls of Moniliformis (Acanthocephala) from Laboratory Rats. Parasitology 1976, 73, 65–72. [Google Scholar] [CrossRef]
- Crompton, D.W.T.; Whitfield, P.J. The Course of Infection and Egg Production of Polymorphus minutus (Acanthocephala) in Domestic Ducks. Parasitology 1968, 58, 231–246. [Google Scholar] [CrossRef]
- Kates, K.C. Some Observations on Experimental Infections of Pigs with the Thorn-Headed Worm, Macracanthorhynchus hirudinaceus. Am. J. Vet. Res. 1944, 5, 166–172. [Google Scholar]
- Hamann, O. Die Nemathelminthen: Beiträge zur Kenntnis ihrer Entwicklung Ihres Baues und ihres Lebensgeschichte; G. Fischer: Jena, Germany, 1891. [Google Scholar]
- Kaiser, J.E. Die Acanthocephalen und ihre Entwickelung; Fischer Theodor: Cassel, France, 1893. [Google Scholar]
- Meyer, A. Acanthocephala. In Bronn’s Klassen und Ordnungen des Tierreichs. 4. Band; Bronn, H.G., Ed.; Akademische Verlagsgesellschaft MBH: Leipzig, Germany, 1933; pp. 335–528. [Google Scholar]
- Atkinson, K.H.A.; Byram, J.E. The Structure of the Ovarian Ball and Oogenesis in Moniliformis dubius (Acanthocephala). J. Morphol. 1976, 148, 391–426. [Google Scholar] [CrossRef] [PubMed]
- Asaolu, S.O.; Whitfield, P.J.; Crompton, D.W.T.; Maxwell, L. Observations on the Development of the Ovarian Balls of Moniliformis (Acanthocephala). Parasitology 1981, 83, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Peura, R.; Valtonen, E.T.; Crompton, D.W.T. Ovarian Tissue in Juvenile Palaeacanthocephalans: Corynosoma semerme, C. strumosum and Echinorhynchus gadi. Parasitology 1986, 93, 471–480. [Google Scholar] [CrossRef]
- Schmidt, G. Development and Life Cycles. In Biology of the Acanthocephala; Crompton, D.W.T., Nickol, B.B., Eds.; Cambridge University Press: Cambridge, UK, 1985; pp. 273–305. ISBN 978-0-521-10511-8. [Google Scholar]
- Herlyn, H.; Röhrig, H. Ultrastructure and Overall Organization of Ligament Sac, Uterine Bell, Uterus and Vagina in Paratenuisentis ambiguus (Acanthocephala, Eoacanthocephala)—The Character Evolution within the Acanthocephala. Acta Zool. 2003, 84, 239–247. [Google Scholar] [CrossRef]
- Davydenko, T.V.; Nikishin, V.P. Features of Tissue Organization in the Female Reproductive System of Acanthocephalus tenuirostris (Palaeacanthocephala, Echinorhynchida). Biol. Bull. 2023, 50, 1556–1566. [Google Scholar] [CrossRef]
- Whitfield, P.J. The Egg Sorting Function of the Uterine Bell of Polymorphus minutus (Acanthocephala). Parasitology 1970, 61, 111–126. [Google Scholar] [CrossRef]
- Whitfield, P.J. A Histological Description of the Uterine Bell of Polymorphus minutus (Acanthocephala). Parasitology 1968, 58, 671–682. [Google Scholar] [CrossRef]
- Asaolu, S.O. Morphology of the Reproductive System of Male Moniliformis dubius (Acanthocephala). Parasitology 1981, 82, 297–309. [Google Scholar] [CrossRef]
- Parshad, V.R.; Crompton, D.W.T. Aspects of Acanthocephalan Reproduction. In Advances in Parasitology; Elsevier: Amsterdam, The Netherlands, 1981; Volume 19, pp. 73–138. ISBN 978-0-12-031719-6. [Google Scholar]
- Miller, D.M.; Dunagan, T.T. Functional Morphology. In Biology of the Acanthocephala; Crompton, D.W.T., Nickol, B.B., Eds.; Cambridge University Press: Cambridge, UK, 1985; pp. 73–123. ISBN 978-0-521-10511-8. [Google Scholar]
- Hyman, L.H. The Invertebrates: Acanthocephala, Aschelminthes, and Entoprocta—The Pseudocoelomate Bilateria (Volume III), 1st ed.; McGraw-Hill: New York, NY, USA, 1951; ISBN 978-0-07-031661-4. [Google Scholar]
- Nicholas, W.L.; Hynes, H.B.N. The Embriology of Polymorphus minutus (Acanthocephala). Proc. Zool. Soc. Lond. 1963, 141, 791–801. [Google Scholar] [CrossRef]
- Marchand, B.; Mattei, X. Fertilization in Acanthocephala. II. Spermatozoon Penetration of Oocyte, Transformation of Gametes and Elaboration of the “Fertilization Membrane”. J. Submicrosc. Cytol. 1980, 12, 95–105. [Google Scholar]
- Robinson, E.S. The Chromosomes of Moniliformis Dubius (Acanthocephala). J. Parasitol. 1965, 51, 430–432. [Google Scholar] [CrossRef] [PubMed]
- Peura, R.; Valtonen, E.T.; Crompton, D.W.T. The Structure of the Mature Oocyte and Zygote of Corynosoma semerme (Acanthocephala). Parasitology 1982, 84, 475–479. [Google Scholar] [CrossRef]
- Marchand, B.; Mattei, X. Présence de Flagelles Spermatiques Dans Les Sphères Ovariennes Des Eoacanthocéphales. J. Ultrastruct. Res. 1976, 56, 331–338. [Google Scholar] [CrossRef]
- Parshad, V.R.; Crompton, D.W.T.; Nesheim, M.C. The Growth of Moniliformis (Acanthocephala) in Rats Fed on Various Monosaccharides and Disaccharides. Proc. R. Soc. Lond. B Biol. Sci. 1980, 209, 299–315. [Google Scholar] [CrossRef]
- Van Cleave, H.J. Acanthocephala of North American mammals. In Illinois Biological Monographs; University of Illinois Press: Urbana, IL, USA, 1953; Volume 23, pp. 1–179. [Google Scholar]
- Guraya, S.S. Histochemical Observations on the Developing Acanthocephalan Oocyte. Acta Embryol. Exp. (Palermo) 1969, 2, 147–155. [Google Scholar]
- Marchand, B.; Mattei, X. La Fécondation Chez Les Acanthocéphales: I. Modifications Ultrastructurales Des Sphères Ovariennes et Des Spermatozoïdes Après Insémination Des Femelles de l’acanthocéphale Neoechinorhynchus agilis. J. Ultrastruct. Res. 1979, 66, 32–39. [Google Scholar] [CrossRef]
- Yamaguti, S. Acanthocephala. In Systema Helminthum; Interscience Publications, John Wiley and Sons: New York, NY, USA; London, UK, 1963; Volume 5, p. 423. [Google Scholar]
- Sayyaf Dezfuli, B.; Rubini, S.; DePasquale, J.A.; Pironi, F. Ultrastructure of Male Centrorhynchus globocaudatus (Acanthocephala) Cement Apparatus and Function of Cement Gland Secretion. J. Helminthol. 2020, 94, e161. [Google Scholar] [CrossRef] [PubMed]
- Crompton, D.W.T. Reproduction. In Biology of the Acanthocephala; Crompton, D.W.T., Nickol, B.B., Eds.; Cambridge University Press: Cambridge, UK, 1985; pp. 213–271. ISBN 978-0-521-10511-8. [Google Scholar]
- Yamaguti, S.; Miyata, I. Notes on Moniliformis dubius, Meyer, 1933. Livro Jubil. Prof Travassos Rio Jan. Braz. 1938, 3, 567–568. [Google Scholar]
- Amin, O.M.; Dailey, M.D. Redescription of Dollfusentis heteracanthus (Acanthocephala: Illiosentidae) from Bonefish, Alhula Vulpes, in the West Indies. J. Helminthol. Soc. Wash. 1996, 63, 31–34. [Google Scholar]
- Dezfuli, B.S.; De Biaggi, S. Copulation of Acanthocephalus anguillae (Acanthocephala). Parasitol. Res. 2000, 86, 524–526. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sayyaf Dezfuli, B.; Pironi, F.; Rossetti, E.; Herlyn, H. New Insights on the Male and Female Reproductive Organs of Centrorhynchus globocaudatus (Acanthocephala), Intestinal Parasite of Birds of Prey. Cells 2024, 13, 356. https://doi.org/10.3390/cells13040356
Sayyaf Dezfuli B, Pironi F, Rossetti E, Herlyn H. New Insights on the Male and Female Reproductive Organs of Centrorhynchus globocaudatus (Acanthocephala), Intestinal Parasite of Birds of Prey. Cells. 2024; 13(4):356. https://doi.org/10.3390/cells13040356
Chicago/Turabian StyleSayyaf Dezfuli, Bahram, Flavio Pironi, Emanuele Rossetti, and Holger Herlyn. 2024. "New Insights on the Male and Female Reproductive Organs of Centrorhynchus globocaudatus (Acanthocephala), Intestinal Parasite of Birds of Prey" Cells 13, no. 4: 356. https://doi.org/10.3390/cells13040356
APA StyleSayyaf Dezfuli, B., Pironi, F., Rossetti, E., & Herlyn, H. (2024). New Insights on the Male and Female Reproductive Organs of Centrorhynchus globocaudatus (Acanthocephala), Intestinal Parasite of Birds of Prey. Cells, 13(4), 356. https://doi.org/10.3390/cells13040356