
Safety of Anti-Reelin Therapeutic Approaches for Chronic Inflammatory Diseases

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Cohort
2.2. Experimental Autoimmune Encephalitis (EAE) Model
2.3. Aging Mice under a Western Diet
2.4. ELISA
2.5. Immunofluorescence
2.6. Immunoblotting
2.7. Field Electrophysiology and θ-Burst Long-Term Potentiation (LTP)
2.8. Statistical Analysis
3. Results
3.1. Reelin Is Stably Expressed throughout Adulthood
3.2. Peripheral Reelin Depletion Preserves Its Expression and Function in the CNS
3.3. Plasma Reelin Depletion Restores Endothelial Function without Abolishing Diapedesis
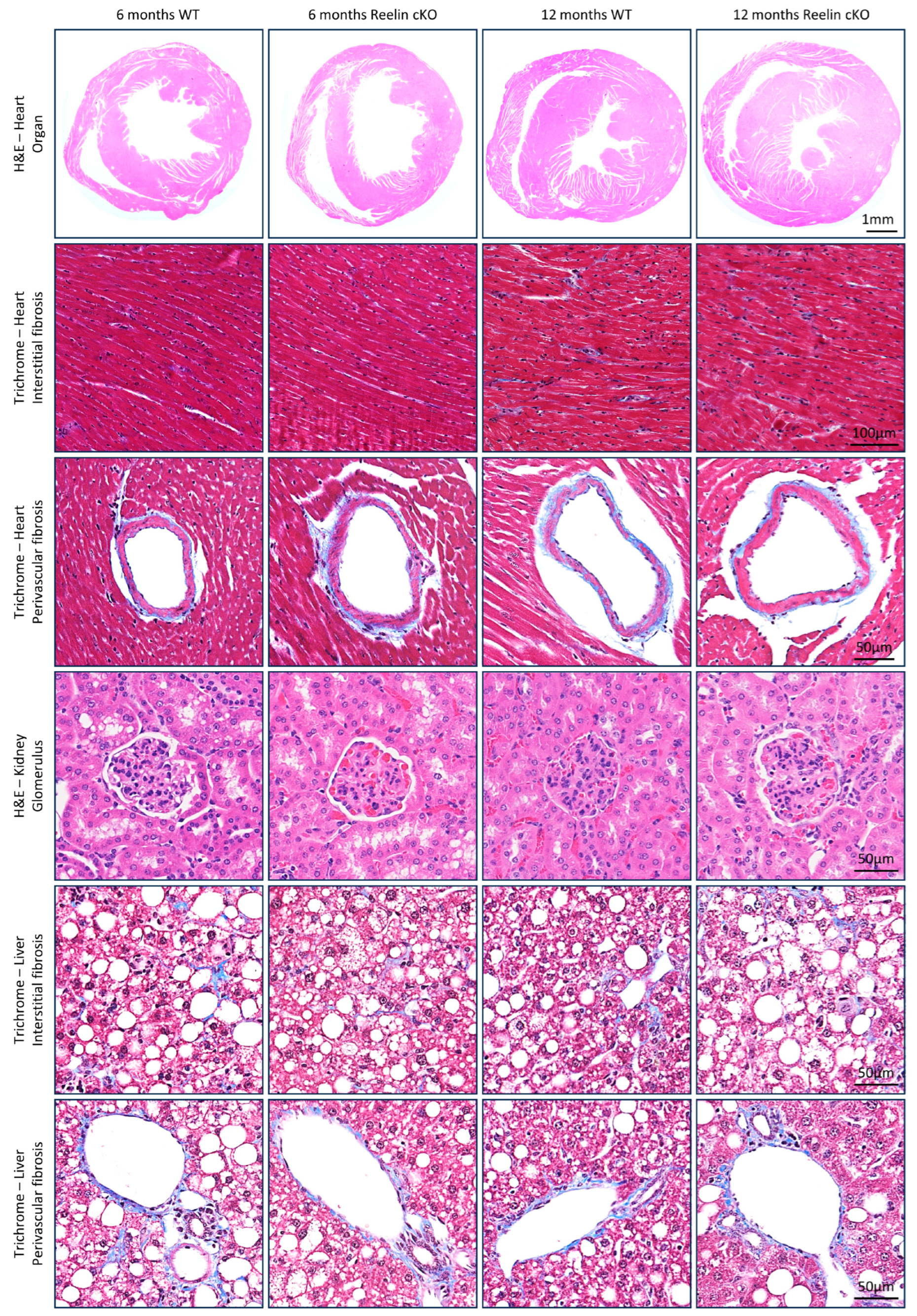
3.4. Plasma Reelin Depletion Does Not Adversely Affect Organ Function Outside the CNS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- D’Arcangelo, G. Reelin in the Years: Controlling Neuronal Migration and Maturation in the Mammalian Brain. Adv. Neurosci. 2014, 2014, 597395. [Google Scholar] [CrossRef]
- Jossin, Y. Reelin Functions, Mechanisms of Action and Signaling Pathways During Brain Development and Maturation. Biomolecules 2020, 10, 964. [Google Scholar] [CrossRef] [PubMed]
- Joly-Amado, A.; Kulkarni, N.; Nash, K.R. Reelin Signaling in Neurodevelopmental Disorders and Neurodegenerative Diseases. Brain Sci. 2023, 13, 1479. [Google Scholar] [CrossRef] [PubMed]
- Lakatosova, S.; Ostatnikova, D. Reelin and its complex involvement in brain development and function. Int. J. Biochem. Cell Biol. 2012, 44, 1501–1504. [Google Scholar] [CrossRef]
- Bock, H.H.; May, P. Canonical and Non-canonical Reelin Signaling. Front. Cell. Neurosci. 2016, 10, 166. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, N.C.; Anderson, R.C.; McDermott, K.W. Reelin: Diverse roles in central nervous system development, health and disease. Int. J. Biochem. Cell Biol. 2019, 112, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Ranaivoson, F.M.; von Daake, S.; Comoletti, D. Structural Insights into Reelin Function: Present and Future. Front. Cell. Neurosci. 2016, 10, 137. [Google Scholar] [CrossRef]
- Förster, E. Reelin, neuronal polarity and process orientation of cortical neurons. Neuroscience 2014, 269, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.T.; Weeber, E.J. Reelin and apoE actions on signal transduction, synaptic function and memory formation. Neuron Glia Biol. 2008, 4, 259–270. [Google Scholar] [CrossRef]
- Dlugosz, P.; Nimpf, J. The Reelin Receptors Apolipoprotein E receptor 2 (ApoER2) and VLDL Receptor. Int. J. Mol. Sci. 2018, 19, 3090. [Google Scholar] [CrossRef]
- Frotscher, M. Dual role of Cajal-Retzius cells and reelin in cortical development. Cell Tissue Res. 1997, 290, 315–322. [Google Scholar] [CrossRef]
- Kelsom, C.; Lu, W. Development and specification of GABAergic cortical interneurons. Cell Biosci. 2013, 3, 19. [Google Scholar] [CrossRef]
- Pesold, C.; Impagnatiello, F.; Pisu, M.G.; Uzunov, D.P.; Costa, E.; Guidotti, A.; Caruncho, H.J. Reelin is preferentially expressed in neurons synthesizing γ-aminobutyric acid in cortex and hippocampus of adult rats. Proc. Natl. Acad. Sci. USA 1998, 95, 3221–3226. [Google Scholar] [CrossRef]
- Weeber, E.J.; Beffert, U.; Jones, C.; Christian, J.M.; Förster, E.; Sweatt, J.D.; Herz, J. Reelin and ApoE Receptors Cooperate to Enhance Hippocampal Synaptic Plasticity and Learning. J. Biol. Chem. 2002, 277, 39944–39952. [Google Scholar] [CrossRef]
- Tsuneura, Y.; Nakai, T.; Mizoguchi, H.; Yamada, K. New Strategies for the Treatment of Neuropsychiatric Disorders Based on Reelin Dysfunction. Int. J. Mol. Sci. 2022, 23, 1829. [Google Scholar] [CrossRef]
- Hirota, Y.; Nakajima, K. Control of Neuronal Migration and Aggregation by Reelin Signaling in the Developing Cerebral Cortex. Front. Cell Dev. Biol. 2017, 5, 40. [Google Scholar] [CrossRef]
- Chai, X.; Förster, E.; Zhao, S.; Bock, H.H.; Frotscher, M. Reelin Stabilizes the Actin Cytoskeleton of Neuronal Processes by Inducing n-Cofilin Phosphorylation at Serine3. J. Neurosci. 2009, 29, 288–299. [Google Scholar] [CrossRef]
- Chai, X.; Frotscher, M. How does Reelin signaling regulate the neuronal cytoskeleton during migration? Neurogenesis 2016, 3, e1242455. [Google Scholar] [CrossRef]
- Frotscher, M.; Chai, X.; Bock, H.H.; Haas, C.A.; Förster, E.; Zhao, S. Role of Reelin in the development and maintenance of cortical lamination. J. Neural Transm. 2009, 116, 1451–1455. [Google Scholar] [CrossRef]
- Frotscher, M. Cajal—Retzius cells, Reelin, and the formation of layers. Curr. Opin. Neurobiol. 1998, 8, 570–575. [Google Scholar] [CrossRef]
- Fatemi, S.H. Reelin glycoprotein: Structure, biology and roles in health and disease. Mol. Psychiatry 2004, 10, 251–257. [Google Scholar] [CrossRef]
- Herz, J.; Chen, Y. Reelin, lipoprotein receptors and synaptic plasticity. Nat. Rev. Neurosci. 2006, 7, 850–859. [Google Scholar] [CrossRef]
- Beffert, U.; Weeber, E.J.; Durudas, A.; Qiu, S.; Masiulis, I.; Sweatt, J.D.; Li, W.-P.; Adelmann, G.; Frotscher, M.; Hammer, R.E.; et al. Modulation of Synaptic Plasticity and Memory by Reelin Involves Differential Splicing of the Lipoprotein Receptor Apoer2. Neuron 2005, 47, 567–579. [Google Scholar] [CrossRef]
- Gong, C.; Wang, T.-W.; Huang, H.S.; Parent, J.M. Reelin Regulates Neuronal Progenitor Migration in Intact and Epileptic Hippocampus. J. Neurosci. 2007, 27, 1803–1811. [Google Scholar] [CrossRef]
- Niu, S.; Renfro, A.; Quattrocchi, C.C.; Sheldon, M.; D’Arcangelo, G. Reelin Promotes Hippocampal Dendrite Development through the VLDLR/ApoER2-Dab1 Pathway. Neuron 2004, 41, 71–84. [Google Scholar] [CrossRef]
- Niu, S.; Yabut, O.; D’Arcangelo, G. The Reelin Signaling Pathway Promotes Dendritic Spine Development in Hippocampal Neurons. J. Neurosci. 2008, 28, 10339–10348. [Google Scholar] [CrossRef]
- Beary, M.; Hodgson, R.; Wildgust, H.J. A critical review of major mortality risk factors for all-cause mortality in first-episode schizophrenia: Clinical and research implications. J. Psychopharmacol. 2012, 26, 52–61. [Google Scholar] [CrossRef]
- Fatemi, S.H.; Kroll, J.L.; Stary, J.M. Altered levels of Reelin and its isoforms in schizophrenia and mood disorders. NeuroReport 2001, 12, 3209–3215. [Google Scholar] [CrossRef]
- Cuchillo-Ibáñez, I.; Andreo-Lillo, P.; Pastor-Ferrándiz, L.; Carratalá-Marco, F.; Sáez-Valero, J. Elevated Plasma Reelin Levels in Children With Autism. Front. Psychiatry 2020, 11, 242. [Google Scholar] [CrossRef]
- Guidotti, A.; Grayson, D.R.; Caruncho, H.J. Epigenetic RELN Dysfunction in Schizophrenia and Related Neuropsychiatric Disorders. Front. Cell. Neurosci. 2016, 10, 89. [Google Scholar] [CrossRef]
- Chen, N.; Bao, Y.; Xue, Y.; Sun, Y.; Hu, D.; Meng, S.; Lu, L.; Shi, J. Meta-analyses of RELN variants in neuropsychiatric disorders. Behav. Brain Res. 2017, 332, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Kelemenova, S.; Ostatnikova, D. Neuroendocrine pathways altered in autism. Special role of reelin.. 2009, 30, 429–436. [Google Scholar]
- Ishii, K.; Kubo, K.-I.; Nakajima, K. Reelin and Neuropsychiatric Disorders. Front. Cell. Neurosci. 2016, 10, 229. [Google Scholar] [CrossRef] [PubMed]
- Knuesel, I. Reelin-mediated signaling in neuropsychiatric and neurodegenerative diseases. Prog. Neurobiol. 2010, 91, 257–274. [Google Scholar] [CrossRef] [PubMed]
- Folsom, T.D.; Fatemi, S.H. The involvement of Reelin in neurodevelopmental disorders. Neuropharmacology 2012, 68, 122–135. [Google Scholar] [CrossRef] [PubMed]
- Lidón, L.; Urrea, L.; Llorens, F.; Gil, V.; Alvarez, I.; Diez-Fairen, M.; Aguilar, M.; Pastor, P.; Zerr, I.; Alcolea, D.; et al. Disease-Specific Changes in Reelin Protein and mRNA in Neurodegenerative Diseases. Cells 2020, 9, 1252. [Google Scholar] [CrossRef] [PubMed]
- Cuchillo-Ibañez, I.; Balmaceda, V.; Mata-Balaguer, T.; Lopez-Font, I.; Sáez-Valero, J. Reelin in Alzheimer’s Disease, Increased Levels but Impaired Signaling: When More is Less. J. Alzheimer’s Dis. 2016, 52, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Yu, N.-N.; Tan, M.-S.; Yu, J.-T.; Xie, A.-M.; Tan, L. The Role of Reelin Signaling in Alzheimer’s Disease. Mol. Neurobiol. 2015, 53, 5692–5700. [Google Scholar] [CrossRef] [PubMed]
- Krstic, D.; Knuesel, I. Deciphering the mechanism underlying late-onset Alzheimer disease. Nat. Rev. Neurol. 2012, 9, 25–34. [Google Scholar] [CrossRef]
- Lee, G.H.; D’arcangelo, G. New Insights into Reelin-Mediated Signaling Pathways. Front. Cell. Neurosci. 2016, 10, 122. [Google Scholar] [CrossRef]
- Alexander, A.; Herz, J.; Calvier, L. Reelin through the years: From brain development to inflammation. Cell Rep. 2023, 42, 112669. [Google Scholar] [CrossRef] [PubMed]
- Yamakage, Y.; Kato, M.; Hongo, A.; Ogino, H.; Ishii, K.; Ishizuka, T.; Kamei, T.; Tsuiji, H.; Miyamoto, T.; Oishi, H.; et al. A disintegrin and metalloproteinase with thrombospondin motifs 2 cleaves and inactivates Reelin in the postnatal cerebral cortex and hippocampus, but not in the cerebellum. Mol. Cell. Neurosci. 2019, 100, 103401. [Google Scholar] [CrossRef] [PubMed]
- Engeroff, K.; Warm, D.; Bittner, S.; Blanquie, O. Different activity patterns control various stages of Reelin synthesis in the developing neocortex. Cereb. Cortex 2023, 33, 9376–9386. [Google Scholar] [CrossRef] [PubMed]
- Sakai, K.; Shoji, H.; Kohno, T.; Miyakawa, T.; Hattori, M. Mice that lack the C-terminal region of Reelin exhibit behavioral abnormalities related to neuropsychiatric disorders. Sci. Rep. 2016, 6, 28636. [Google Scholar] [CrossRef] [PubMed]
- Jossin, Y.; Gui, L.; Goffinet, A.M. Processing of Reelin by Embryonic Neurons Is Important for Function in Tissue But Not in Dissociated Cultured Neurons. J. Neurosci. 2007, 27, 4243–4252. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.H.; Chhangawala, Z.; von Daake, S.; Savas, J.N.; Yates, J.R.; Comoletti, D.; D’Arcangelo, G. Reelin Induces Erk1/2 Signaling in Cortical Neurons Through a Non-canonical Pathway. J. Biol. Chem. 2014, 289, 20307–20317. [Google Scholar] [CrossRef]
- Hattori, M.; Kohno, T. Regulation of Reelin functions by specific proteolytic processing in the brain. J. Biochem. 2021, 169, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Ogino, H.; Hisanaga, A.; Kohno, T.; Kondo, Y.; Okumura, K.; Kamei, T.; Sato, T.; Asahara, H.; Tsuiji, H.; Fukata, M.; et al. Secreted Metalloproteinase ADAMTS-3 Inactivates Reelin. J. Neurosci. 2017, 37, 3181–3191. [Google Scholar] [CrossRef]
- Nakamura, K.; Beppu, M.; Sakai, K.; Yagyu, H.; Matsumaru, S.; Kohno, T.; Hattori, M. The C-terminal region of Reelin is necessary for proper positioning of a subset of Purkinje cells in the postnatal cerebellum. Neuroscience 2016, 336, 20–29. [Google Scholar] [CrossRef]
- Okugawa, E.; Ogino, H.; Shigenobu, T.; Yamakage, Y.; Tsuiji, H.; Oishi, H.; Kohno, T.; Hattori, M. Physiological significance of proteolytic processing of Reelin revealed by cleavage-resistant Reelin knock-in mice. Sci. Rep. 2020, 10, 4471. [Google Scholar] [CrossRef]
- Okoro, E.U.; Zhang, H.; Guo, Z.; Yang, F.; Smith, C.; Yang, H. A Subregion of Reelin Suppresses Lipoprotein-Induced Cholesterol Accumulation in Macrophages. PLoS ONE 2015, 10, e0136895. [Google Scholar] [CrossRef] [PubMed]
- Koie, M.; Okumura, K.; Hisanaga, A.; Kamei, T.; Sasaki, K.; Deng, M.; Baba, A.; Kohno, T.; Hattori, M. Cleavage within Reelin Repeat 3 Regulates the Duration and Range of the Signaling Activity of Reelin Protein. J. Biol. Chem. 2014, 289, 12922–12930. [Google Scholar] [CrossRef] [PubMed]
- Calvier, L.; Demuth, G.; Manouchehri, N.; Wong, C.; Sacharidou, A.; Mineo, C.; Shaul, P.W.; Monson, N.L.; Kounnas, M.Z.; Stüve, O.; et al. Reelin depletion protects against autoimmune encephalomyelitis by decreasing vascular adhesion of leukocytes. Sci. Transl. Med. 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Esmaeilzadeh-Gharehdaghi, E.; Razmara, E.; Bitaraf, A.; Jamshidi, A.; Mahmoudi, M.; Garshasbi, M. Functional Analysis of RELN S2486G Mutation and its Contribution to Pathogenesis of Ankylosing Spondylitis. Int. J. Infect. Dis. 2020, 23, 688–696. [Google Scholar] [CrossRef]
- Garshasbi, M.; Mahmoudi, M.; Razmara, E.; Vojdanian, M.; Aslani, S.; Farhadi, E.; Jensen, L.R.; Arzaghi, S.M.; Poursani, S.; Bitaraf, A.; et al. Identification of RELN variant p.(Ser2486Gly) in an Iranian family with ankylosing spondylitis; the first association of RELN and AS. Eur. J. Hum. Genet. 2020, 28, 754–762. [Google Scholar] [CrossRef] [PubMed]
- Magnani, A.; Pattacini, L.; Boiardi, L.; Casali, B.; Salvarani, C. Reelin levels are increased in synovial fluid of patients with rheumatoid arthritis. Clin. Exp. Rheumatol. 2010, 28, 546–548. [Google Scholar] [PubMed]
- Tseng, W.; Chen, T.; Huang, C.; Huang, Y.; Yeh, C.; Tsai, H.; Lee, H.; Kao, C.; Lin, S.; Liao, H.; et al. Impaired thrombin generation in Reelin-deficient mice: A potential role of plasma Reelin in hemostasis. J. Thromb. Haemost. 2014, 12, 2054–2064. [Google Scholar] [CrossRef]
- Ding, Y.; Huang, L.; Xian, X.; Yuhanna, I.S.; Wasser, C.R.; Frotscher, M.; Mineo, C.; Shaul, P.W.; Herz, J. Loss of Reelin protects against atherosclerosis by reducing leukocyte–endothelial cell adhesion and lesion macrophage accumulation. Sci. Signal. 2016, 9, ra29. [Google Scholar] [CrossRef] [PubMed]
- Roberts, B.L.; Bennett, B.J.; Bennett, C.M.; Carroll, J.M.; Dalbøge, L.S.; Hall, C.; Hassouneh, W.; Heppner, K.M.; Kirigiti, M.A.; Lindsley, S.R.; et al. Reelin is modulated by diet-induced obesity and has direct actions on arcuate proopiomelanocortin neurons. Mol. Metab. 2019, 26, 18–29. [Google Scholar] [CrossRef]
- Gowert, N.S.; Krüger, I.; Klier, M.; Donner, L.; Kipkeew, F.; Gliem, M.; Bradshaw, N.J.; Lutz, D.; Köber, S.; Langer, H.; et al. Loss of Reelin protects mice against arterial thrombosis by impairing integrin activation and thrombus formation under high shear conditions. Cell. Signal. 2017, 40, 210–221. [Google Scholar] [CrossRef]
- Carvajal, A.E.; Vázquez-Carretero, M.D.; García-Miranda, P.; Peral, M.J.; Calonge, M.L.; Ilundain, A.A. Reelin expression is up-regulated in mice colon in response to acute colitis and provides resistance against colitis. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2017, 1863, 462–473. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Morales, J.M.; Vázquez-Carretero, M.D.; García-Miranda, P.; Carvajal, A.E.; Calonge, M.L.; Ilundain, A.A.; Peral, M.J. Reelin Protects against Colon Pathology via p53 and May Be a Biomarker for Colon Cancer Progression. Biology 2022, 11, 1406. [Google Scholar] [CrossRef]
- Carvajal, A.E.; Serrano-Morales, J.M.; Vázquez-Carretero, M.D.; García-Miranda, P.; Calonge, M.L.; Peral, M.J.; Ilundain, A.A. Reelin protects from colon pathology by maintaining the intestinal barrier integrity and repressing tumorigenic genes. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2017, 1863, 2126–2134. [Google Scholar] [CrossRef] [PubMed]
- Perez-Costas, E.; Fenton, E.Y.; Caruncho, H.J. Reelin expression in brain endothelial cells: An electron microscopy study. BMC Neurosci. 2015, 16, 16. [Google Scholar] [CrossRef]
- Pei, L.; Ouyang, Z.; Zhang, H.; Huang, S.; Jiang, R.; Liu, B.; Tang, Y.; Feng, M.; Yuan, M.; Wang, H.; et al. Thrombospondin 1 and Reelin act through Vldlr to regulate cardiac growth and repair. Basic Res. Cardiol. 2023, 119, 169–192. [Google Scholar] [CrossRef] [PubMed]
- Krueger, I.; Gremer, L.; Mangels, L.; Klier, M.; Jurk, K.; Willbold, D.; Bock, H.H.; Elvers, M. Reelin Amplifies Glycoprotein VI Activation and AlphaIIb Beta3 Integrin Outside-In Signaling via PLC Gamma 2 and Rho GTPases. Arter. Thromb. Vasc. Biol. 2020, 40, 2391–2403. [Google Scholar] [CrossRef] [PubMed]
- Tseng, W.-L.; Huang, C.-L.; Chong, K.-Y.; Liao, C.-H.; Stern, A.; Cheng, J.-C.; Tseng, C.-P. Reelin is a platelet protein and functions as a positive regulator of platelet spreading on fibrinogen. Cell. Mol. Life Sci. 2009, 67, 641–653. [Google Scholar] [CrossRef] [PubMed]
- Sturm, L.; Roth, L.; Zoldan, K.; Schultheiss, M.; Boettler, T.; Huber, J.P.; Kaeser, R.; Thimme, R.; Bettinger, D. Blood reelin in the progression of chronic liver disease. Adv. Med. Sci. 2021, 66, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Carotti, S.; Perrone, G.; Amato, M.; Gentilucci, U.V.; Righi, D.; Francesconi, M.; Pellegrini, C.; Zalfa, F.; Zingariello, M.; Picardi, A.; et al. Reelin expression in human liver of patients with chronic hepatitis C infection. Eur. J. Histochem. 2017, 61, 2745. [Google Scholar] [CrossRef] [PubMed]
- Botella-López, A.; de Madaria, E.; Jover, R.; Bataller, R.; Sancho-Bru, P.; Candela, A.; Compañ, A.; Pérez-Mateo, M.; Martínez, S.; Sáez-Valero, J. Reelin is overexpressed in the liver and plasma of bile duct ligated rats and its levels and glycosylation are altered in plasma of humans with cirrhosis. Int. J. Biochem. Cell Biol. 2007, 40, 766–775. [Google Scholar] [CrossRef]
- Csomor, P.; Sziklai, I.; Karosi, T. Controversies in RELN/reelin expression in otosclerosis. Eur. Arch. Oto-Rhino-Laryngol. 2011, 269, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Calvier, L.; Xian, X.; Lee, R.G.; Sacharidou, A.; Mineo, C.; Shaul, P.W.; Kounnas, M.Z.; Tsai, S.; Herz, J. Reelin Depletion Protects Against Atherosclerosis by Decreasing Vascular Adhesion of Leukocytes. Arter. Thromb. Vasc. Biol. 2021, 41, 1309–1318. [Google Scholar] [CrossRef] [PubMed]
- Calvier, L.; Manouchehri, N.; Sacharidou, A.; Mineo, C.; Shaul, P.W.; Hui, D.Y.; Kounnas, M.Z.; Stüve, O.; Herz, J. Apolipoprotein E receptor 2 deficiency decreases endothelial adhesion of monocytes and protects against autoimmune encephalomyelitis. Sci. Immunol. 2021, 6. [Google Scholar] [CrossRef] [PubMed]
- Calvier, L.; Drelich, A.; Hsu, J.; Tseng, C.-T.; Mina, Y.; Nath, A.; Kounnas, M.Z.; Herz, J. Circulating Reelin promotes inflammation and modulates disease activity in acute and long COVID-19 cases. Front. Immunol. 2023, 14, 1185748. [Google Scholar] [CrossRef] [PubMed]
- Sacharidou, A.; Shaul, P.W.; Mineo, C. New Insights in the Pathophysiology of Antiphospholipid Syndrome. Semin. Thromb. Hemost. 2017, 44, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Riitano, G.; Capozzi, A.; Recalchi, S.; Augusto, M.; Conti, F.; Misasi, R.; Garofalo, T.; Sorice, M.; Manganelli, V. Role of Lipid Rafts on LRP8 Signaling Triggered by Anti-β2-GPI Antibodies in Endothelial Cells. Biomedicines 2023, 11, 3135. [Google Scholar] [CrossRef] [PubMed]
- Jeong, E.-M.; Kim, C.-S.; Lee, S.H.; Baik, E.J.; Moon, C.K.; Jung, Y.-S. Cadmium stimulates the expression of ICAM-1 via NF-κB activation in cerebrovascular endothelial cells. Biochem. Biophys. Res. Commun. 2004, 320, 887–892. [Google Scholar] [CrossRef] [PubMed]
- Mo, S.-J.; Son, E.-W.; Lee, S.-R.; Lee, S.-M.; Shin, D.-H.; Pyo, S. CML-1 inhibits TNF-α-induced NF-κB activation and adhesion molecule expression in endothelial cells through inhibition of IkBα kinase. J. Ethnopharmacol. 2007, 109, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.-Z.; Tsukahara, H.; Ohshima, Y.; Todoroki, Y.; Hiraoka, M.; Maeda, M.; Mayumi, M. Effects of antioxidants and nitric oxide on TNF-α-induced adhesion molecule expression and NF-κB activation in human dermal microvascular endothelial cells. Life Sci. 2004, 75, 1159–1170. [Google Scholar] [CrossRef]
- Gareus, R.; Kotsaki, E.; Xanthoulea, S.; van der Made, I.; Gijbels, M.J.; Kardakaris, R.; Polykratis, A.; Kollias, G.; de Winther, M.P.; Pasparakis, M. Endothelial Cell-Specific NF-κB Inhibition Protects Mice from Atherosclerosis. Cell Metab. 2008, 8, 372–383. [Google Scholar] [CrossRef]
- Gaua, R.J.; Yangb, H.L.; Chowc, S.N.; Suend, J.L.; Lu, F.-J. Humic Acid Suppresses the LPS-Induced Expression of Cell-Surface Adhesion Proteins through the Inhibition of NF-κB Activation. Toxicol. Appl. Pharmacol. 2000, 166, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Morigi, M.; Angioletti, S.; Imberti, B.; Donadelli, R.; Micheletti, G.; Figliuzzi, M.; Remuzzi, A.; Zoja, C.; Remuzzi, G. Leukocyte-endothelial interaction is augmented by high glucose concentrations and hyperglycemia in a NF-kB-dependent fashion. J. Clin. Investig. 1998, 101, 1905–1915. [Google Scholar] [CrossRef] [PubMed]
- Cowan, C.; Muraleedharan, C.K.; O’Donnell, J.J.; Singh, P.K.; Lum, H.; Kumar, A.; Xu, S. MicroRNA-146 Inhibits Thrombin-Induced NF- B Activation and Subsequent Inflammatory Responses in Human Retinal Endothelial Cells. Investig. Opthalmology Vis. Sci. 2014, 55, 4944–4951. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Yang, H.; Lu, X.; Wang, L.; Li, H.; Chen, S.; Wang, X.; Shen, C.; Huang, J.; Lu, X.; et al. MiR-520b inhibits endothelial activation by targeting NF-κB p65-VCAM1 axis. Biochem. Pharmacol. 2021, 188, 114540. [Google Scholar] [CrossRef] [PubMed]
- Baker, R.G.; Hayden, M.S.; Ghosh, S. NF-κB, Inflammation, and Metabolic Disease. Cell Metab. 2011, 13, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Zhu, M.-M.; Zhang, M.-H.; Wang, R.-S.; Tan, X.-B.; Song, J.; Ding, S.-M.; Jia, X.-B.; Hu, S.-Y. Protection of glycyrrhizic acid against AGEs-induced endothelial dysfunction through inhibiting RAGE/NF-κB pathway activation in human umbilical vein endothelial cells. J. Ethnopharmacol. 2013, 148, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.A.; Fuseler, J.W. Regulation of NF-κB activation and nuclear translocation by exogenous nitric oxide (NO) donors in TNF-α activated vascular endothelial cells. Nitric Oxide 2007, 16, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Madonna, R.; De Caterina, R. Relevance of new drug discovery to reduce NF-κB activation in cardiovascular disease. Vasc. Pharmacol. 2012, 57, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Lindner, V. The NF-κB and IκB System in Injured Arteries. Pathobiology 1998, 66, 311–320. [Google Scholar] [CrossRef]
- Min, J.-K.; Kim, Y.-M.; Kim, S.W.; Kwon, M.-C.; Kong, Y.-Y.; Hwang, I.K.; Won, M.H.; Rho, J.; Kwon, Y.-G. TNF-Related Activation-Induced Cytokine Enhances Leukocyte Adhesiveness: Induction of ICAM-1 and VCAM-1 via TNF Receptor-Associated Factor and Protein Kinase C-Dependent NF-KappaB Activation in Endothelial Cells. J. Immunol. 2005, 175, 531–540. [Google Scholar] [CrossRef]
- Collins, T.; Read, M.A.; Neish, A.S.; Whitley, M.Z.; Thanos, D.; Maniatis, T. Transcriptional regulation of endothelial cell adhesion molecules: NF-kappa B and cytokine-inducible enhancers. FASEB J. 1995, 9, 899–909. [Google Scholar] [CrossRef]
- Harris, D.P.; Bandyopadhyay, S.; Maxwell, T.J.; Willard, B.; DiCorleto, P.E. Tumor Necrosis Factor (TNF)-α Induction of CXCL10 in Endothelial Cells Requires Protein Arginine Methyltransferase 5 (PRMT5)-mediated Nuclear Factor (NF)-κB p65 Methylation. J. Biol. Chem. 2014, 289, 15328–15339. [Google Scholar] [CrossRef]
- Prisco, A.R.; Hoffmann, B.R.; Kaczorowski, C.C.; McDermott-Roe, C.; Stodola, T.J.; Exner, E.C.; Greene, A.S. Tumor Necrosis Factor α Regulates Endothelial Progenitor Cell Migration via CADM1 and NF-kB. Stem Cells 2016, 34, 1922–1933. [Google Scholar] [CrossRef]
- Romay-Penabad, Z.; Aguilar-Valenzuela, R.; Urbanus, R.T.; Derksen, R.H.W.M.; Pennings, M.T.T.; Papalardo, E.; Shilagard, T.; Vargas, G.; Hwang, Y.; de Groot, P.G.; et al. Apolipoprotein E receptor 2 is involved in the thrombotic complications in a murine model of the antiphospholipid syndrome. Blood 2011, 117, 1408–1414. [Google Scholar] [CrossRef]
- Samama, B.; Boehm, N. Reelin immunoreactivity in lymphatics and liver during development and adult life. Anat. Rec. Part A Discov. Mol. Cell. Evol. Biol. 2005, 285A, 595–599. [Google Scholar] [CrossRef]
- Smalheiser, N.R.; Costa, E.; Guidotti, A.; Impagnatiello, F.; Auta, J.; Lacor, P.; Kriho, V.; Pappas, G.D. Expression of reelin in adult mammalian blood, liver, pituitary pars intermedia, and adrenal chromaffin cells. Proc. Natl. Acad. Sci. USA 2000, 97, 1281–1286. [Google Scholar] [CrossRef]
- Kobold, D.; Grundmann, A.; Piscaglia, F.; Eisenbach, C.; Neubauer, K.; Steffgen, J.; Ramadori, G.; Knittel, T. Expression of reelin in hepatic stellate cells and during hepatic tissue repair: A novel marker for the differentiation of HSC from other liver myofibroblasts. J. Hepatol. 2002, 36, 607–613. [Google Scholar] [CrossRef]
- Calvier, L.; Alexander, A.E.; Herz, J. The “6B” Strategy: Build Back a Better Blood–Brain Barrier. Immuno 2022, 2, 506–511. [Google Scholar] [CrossRef]
- Katsuyama, Y.; Terashima, T. Developmental anatomy of reeler mutant mouse. Dev. Growth Differ. 2009, 51, 271–286. [Google Scholar] [CrossRef]
- Terashima, T.; Inoue, K.; Inoue, Y. Dislocated neurons and neural network: Hodological study of the motor cortex of the reeler mutant mouse. Hokkaido J. Med. Sci. 1991, 66, 268–285. [Google Scholar]
- Goffinet, A. Events governing organization of postmigratory neurons: Studies on brain development in normal and reeler mice. Brain Res. Rev. 1984, 7, 261–296. [Google Scholar] [CrossRef]
- Luque, J.M. Puzzling out the reeler brainteaser: Does reelin signal to unique neural lineages? Brain Res. 2007, 1140, 41–50. [Google Scholar] [CrossRef]
- D’Arcangelo, G.; Curran, T. Reeler: New Tales on an Old Mutant Mouse. Bioessays 1998, 20, 235–244. [Google Scholar] [CrossRef]
- D’arcangelo, G. Reelin mouse mutants as models of cortical development disorders. Epilepsy Behav. 2006, 8, 81–90. [Google Scholar] [CrossRef]
- Lambert de Rouvroit, C.; Goffinet, A.M. The Reeler Mouse as a Model of Brain Development. Adv Anat Embryol Cell Biol 1998, 150, 1–106. [Google Scholar]
- Lossi, L.; Castagna, C.; Granato, A.; Merighi, A. The Reeler Mouse: A Translational Model of Human Neurological Conditions, or Simply a Good Tool for Better Understanding Neurodevelopment? J. Clin. Med. 2019, 8, 2088. [Google Scholar] [CrossRef]
- D’Arcangelo, G. The Reeler Mouse: Anatomy of a Mutant. Int. Rev. Neurobiol. 2005, 71, 383–417. [Google Scholar] [CrossRef]
- Falconer, D.S. Two new mutants, ‘trembler’ and ‘reeler’, with neurological actions in the house mouse (Mus musculus L.). J. Genet. 1951, 50, 192–205. [Google Scholar] [CrossRef]
- D’Arcangelo, G.; Miao, G.G.; Chen, S.-C.; Scares, H.D.; Morgan, J.I.; Curran, T. A protein related to extracellular matrix proteins deleted in the mouse mutant reeler. Nature 1995, 374, 719–723. [Google Scholar] [CrossRef]
- Ogawa, M.; Miyata, T.; Nakajimat, K.; Yagyu, K.; Seike, M.; Ikenaka, K.; Yamamoto, H.; Mikoshibat, K. The reeler gene-associated antigen on cajal-retzius neurons is a crucial molecule for laminar organization of cortical neurons. Neuron 1995, 14, 899–912. [Google Scholar] [CrossRef]
- Yoneshima, H.; Nagata, E.; Matsumoto, M.; Yamada, M.; Nakajima, K.; Miyata, T.; Ogawa, M.; Mikoshiba, K. A novel neurological mutant mouse, yotari, which exhibits reeler-like phenotype but expresses CR-50 antigen/Reelin. Neurosci. Res. 1997, 29, 217–223. [Google Scholar] [CrossRef]
- Drakew, A.; Frotscher, M.; Deller, T.; Ogawa, M.; Heimrich, B. Developmental distribution of a reeler gene-related antigen in the rat hippocampal formation visualized by CR-50 immunocytochemistry. Neuroscience 1997, 82, 1079–1086. [Google Scholar] [CrossRef]
- Nakajima, K.; Mikoshiba, K.; Miyata, T.; Kudo, C.; Ogawa, M. Disruption of hippocampal development in vivo by CR-50 mAb against Reelin. Proc. Natl. Acad. Sci. USA 1997, 94, 8196–8201. [Google Scholar] [CrossRef]
- D’arcangelo, G.; Nakajima, K.; Miyata, T.; Ogawa, M.; Mikoshiba, K.; Curran, T. Reelin Is a Secreted Glycoprotein Recognized by the CR-50 Monoclonal Antibody. J. Neurosci. 1997, 17, 23–31. [Google Scholar] [CrossRef]
- Utsunomiya-Tate, N.; Kubo, K.-I.; Tate, S.-I.; Kainosho, M.; Katayama, E.; Nakajima, K.; Mikoshiba, K. Reelin molecules assemble together to form a large protein complex, which is inhibited by the function-blocking CR-50 antibody. Proc. Natl. Acad. Sci. USA 2000, 97, 9729–9734. [Google Scholar] [CrossRef]
- Miyata, T.; Nakajima, K.; Mikoshiba, K.; Ogawa, M. Regulation of Purkinje Cell Alignment by Reelin as Revealed with CR-50 Antibody. J. Neurosci. 1997, 17, 3599–3609. [Google Scholar] [CrossRef]
- Lane-Donovan, C.; Philips, G.T.; Wasser, C.R.; Durakoglugil, M.S.; Masiulis, I.; Upadhaya, A.; Pohlkamp, T.; Coskun, C.; Kotti, T.; Steller, L.; et al. Reelin protects against amyloid β toxicity in vivo. Sci. Signal. 2015, 8, ra67. [Google Scholar] [CrossRef]
- Bittner, S.; Afzali, A.M.; Wiendl, H.; Meuth, S.G. Myelin Oligodendrocyte Glycoprotein (MOG35-55) Induced Experimental Autoimmune Encephalomyelitis (EAE) in C57BL/6 Mice. J. Vis. Exp. 2014, e51275. [Google Scholar] [CrossRef]
- Autry, A.E.; Adachi, M.; Nosyreva, E.; Na, E.S.; Los, M.F.; Cheng, P.-F.; Kavalali, E.T.; Monteggia, L.M. NMDA receptor blockade at rest triggers rapid behavioural antidepressant responses. Nature 2011, 475, 91–95. [Google Scholar] [CrossRef]
- Kato, K.; Sekino, Y.; Takahashi, H.; Yasuda, H.; Shirao, T. Increase in AMPA receptor-mediated miniature EPSC amplitude after chronic NMDA receptor blockade in cultured hippocampal neurons. Neurosci. Lett. 2007, 418, 4–8. [Google Scholar] [CrossRef]
- Ajami, B.; Bennett, J.L.; Krieger, C.; McNagny, K.M.; Rossi, F.M.V. Infiltrating monocytes trigger EAE progression, but do not contribute to the resident microglia pool. Nat. Neurosci. 2011, 14, 1142–1149. [Google Scholar] [CrossRef]
- Lucchinetti, C.; Brück, W.; Parisi, J.; Scheithauer, B.; Rodriguez, M.; Lassmann, H. Heterogeneity of Multiple Sclerosis Lesions: Implications for the Pathogenesis of Demyelination. Ann. Neurol. 2000, 47, 707–717. [Google Scholar] [CrossRef]
- Manouchehri, N.; Hussain, R.Z.; Cravens, P.D.; Esaulova, E.; Artyomov, M.N.; Edelson, B.T.; Wu, G.F.; Cross, A.H.; Doelger, R.; Loof, N.; et al. CD11c+CD88+CD317+myeloid cells are critical mediators of persistent CNS autoimmunity. Proc. Natl. Acad. Sci. USA 2021, 118, e2014492118. [Google Scholar] [CrossRef]
- Calvier, L.; Martinez-Martinez, E.; Miana, M.; Cachofeiro, V.; Rousseau, E.; Sádaba, J.R.; Zannad, F.; Rossignol, P.; López-Andrés, N. The Impact of Galectin-3 Inhibition on Aldosterone-Induced Cardiac and Renal Injuries. JACC Hear. Fail. 2015, 3, 59–67. [Google Scholar] [CrossRef]
- Stüve, O.; Marra, C.M.; Jerome, K.R.; Cook, L.; Cravens, P.D.; Cepok, S.; Frohman, E.M.; Phillips, J.T.; Arendt, G.; Hemmer, B.; et al. Immune surveillance in multiple sclerosis patients treated with natalizumab. Ann. Neurol. 2006, 59, 743–747. [Google Scholar] [CrossRef]
- Cutter, G.R.; Stüve, O. Does risk stratification decrease the risk of natalizumab-associated PML? Where is the evidence? Mult. Scler. J. 2014, 20, 1304–1305. [Google Scholar] [CrossRef]
- Hussey, S.; Bergman, M. The Gut Microbiota and Effects on Metabolism. In Pathobiology of Human Disease: A Dynamic Encyclopedia of Disease Mechanisms; Academic Press: Amsterdam, The Netherlands, 2014; ISBN 978-0-12-386457-4. [Google Scholar]
- Medzhitov, R. Inflammation 2010: New Adventures of an Old Flame. Cell 2010, 140, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.E.; Shugart, Y.Y.; Huang, D.T.; Al Shahwan, S.; Grant, P.E.; Hourihane, J.O.; Martin, N.D.; Walsh, C.A. Autosomal recessive lissencephaly with cerebellar hypoplasia is associated with human RELN mutations. Nat. Genet. 2000, 26, 93–96. [Google Scholar] [CrossRef]
- Tsao, C.W.; Aday, A.W.; Almarzooq, Z.I.; Anderson, C.A.; Arora, P.; Avery, C.L.; Baker-Smith, C.M.; Beaton, A.Z.; Boehme, A.K.; Buxton, A.E.; et al. Heart Disease and Stroke Statistics—2023 Update: A Report From the American Heart Association. Circulation 2023, 147, E93–E621. [Google Scholar] [CrossRef]
- Calvier, L.; Miana, M.; Reboul, P.; Cachofeiro, V.; Martinez-Martinez, E.; de Boer, R.A.; Poirier, F.; Lacolley, P.; Zannad, F.; Rossignol, P.; et al. Galectin-3 mediates aldosterone-induced vascular fibrosis. Atheroscler. Thromb. Vasc. Biol. 2013, 33, 67–75. [Google Scholar] [CrossRef]
- Martinez-Martinez, E.; Ibarrola, J.; Calvier, L.; Fernandez-Celis, A.; Leroy, C.; Cachofeiro, V.; Rossignol, P.; Lopez-Andres, N. Galectin-3 Blockade Reduces Renal Fibrosis in Two Normotensive Experimental Models of Renal Damage. PLoS ONE 2016, 11, e0166272. [Google Scholar] [CrossRef]
- Martínez-Martínez, E.; Ibarrola, J.; Fernández-Celis, A.; Calvier, L.; Leroy, C.; Cachofeiro, V.; Rossignol, P.; López-Andrés, N. Galectin-3 pharmacological inhibition attenuates early renal damage in spontaneously hypertensive rats. J. Hypertens. 2018, 36, 368–376. [Google Scholar] [CrossRef]
- López-Andrés, N.; Calvier, L.; Labat, C.; Fay, R.; Díez, J.; Benetos, A.; Zannad, F.; Lacolley, P.; Rossignol, P. Absence of Cardiotrophin 1 Is Associated With Decreased Age-Dependent Arterial Stiffness and Increased Longevity in Mice. Hypertension 2013, 61, 120–129. [Google Scholar] [CrossRef]
- López-Andrés, N.; Rousseau, A.; Akhtar, R.; Calvier, L.; Iñigo, C.; Labat, C.; Zhao, X.; Cruickshank, K.; Díez, J.; Zannad, F.; et al. Cardiotrophin 1 Is Involved in Cardiac, Vascular, and Renal Fibrosis and Dysfunction. Hypertension 2012, 60, 563–573. [Google Scholar] [CrossRef]
- Calvier, L.; Chouvarine, P.; Legchenko, E.; Hoffmann, N.; Geldner, J.; Borchert, P.; Jonigk, D.; Mozes, M.M.; Hansmann, G. PPARγ Links BMP2 and TGFβ1 Pathways in Vascular Smooth Muscle Cells, Regulating Cell Proliferation and Glucose Metabolism. Cell Metab. 2017, 25, 1118–1134.e7. [Google Scholar] [CrossRef]
- Calvier, L.; Legchenko, E.; Grimm, L.; Sallmon, H.; Hatch, A.; Plouffe, B.D.; Schroeder, C.; Bauersachs, J.; Murthy, S.K.; Hansmann, G. Galectin-3 and aldosterone as potential tandem biomarkers in pulmonary arterial hypertension. Heart 2016, 102, 390–396. [Google Scholar] [CrossRef]
- Calvier, L.; Boucher, P.; Herz, J.; Hansmann, G. LRP1 Deficiency in Vascular SMC Leads to Pulmonary Arterial Hypertension That Is Reversed by PPARγ Activation. Circ. Res. 2019, 124, 1778–1785. [Google Scholar] [CrossRef]
- Kökény, G.; Calvier, L.; Legchenko, E.; Chouvarine, P.; Mózes, M.M.; Hansmann, G. PPARγ is a gatekeeper for extracellular matrix and vascular cell homeostasis. Curr. Opin. Nephrol. Hypertens. 2020, 29, 171–179. [Google Scholar] [CrossRef]
- Kökény, G.; Calvier, L.; Hansmann, G. PPARγ and TGFβ—Major Regulators of Metabolism, Inflammation, and Fibrosis in the Lungs and Kidneys. Int. J. Mol. Sci. 2021, 22, 10431. [Google Scholar] [CrossRef] [PubMed]
- Calvier, L.; Chouvarine, P.; Legchenko, E.; Hansmann, G. Transforming Growth Factor β1– and Bone Morphogenetic Protein 2/PPARγ–regulated MicroRNAs in Pulmonary Arterial Hypertension. Am. J. Respir. Crit. Care Med. 2017, 196, 1227–1228. [Google Scholar] [CrossRef] [PubMed]
- Calvier, L.; Chouvarine, P.; Legchenko, E.; Kokeny, G.; Mozes, M.M.; Hansmann, G. Chronic TGF-β1 Signaling in Pulmonary Arterial Hypertension Induces Sustained Canonical Smad3 Pathways in Vascular Smooth Muscle Cells. Am. J. Respir. Cell Mol. Biol. 2019, 61, 121–123. [Google Scholar] [CrossRef]
- Hansmann, G.; Calvier, L.; Risbano, M.G.; Chan, S.Y. Activation of the Metabolic Master Regulator PPARγ: A Potential PIOneering Therapy for Pulmonary Arterial Hypertension. Am. J. Respir. Cell Mol. Biol. 2020, 62, 143–156. [Google Scholar] [CrossRef]
- Qin, X.; Lin, L.; Cao, L.; Zhang, X.; Song, X.; Hao, J.; Zhang, Y.; Wei, R.; Huang, X.; Lu, J.; et al. Extracellular matrix protein Reelin promotes myeloma progression by facilitating tumor cell proliferation and glycolysis. Sci. Rep. 2017, 7, 45305. [Google Scholar] [CrossRef]
- Becker, J.; Fröhlich, J.; Perske, C.; Pavlakovic, H.; Wilting, J. Reelin signalling in neuroblastoma: Migratory switch in metastatic stages. Int. J. Oncol. 2012, 41, 681–689. [Google Scholar] [CrossRef]
- Lin, L.; Yan, F.; Zhao, D.; Lv, M.; Liang, X.; Dai, H.; Qin, X.; Zhang, Y.; Hao, J.; Sun, X.; et al. Reelin promotes the adhesion and drug resistance of multiple myeloma cells via integrin β1 signaling and STAT3. Oncotarget 2016, 7, 9844–9858. [Google Scholar] [CrossRef]
- Perrone, G.; Vincenzi, B.; Zagami, M.; Santini, D.; Panteri, R.; Flammia, G.; Verzì, A.; Lepanto, D.; Morini, S.; Russo, A.; et al. Reelin expression in human prostate cancer: A marker of tumor aggressiveness based on correlation with grade. Mod. Pathol. 2007, 20, 344–351. [Google Scholar] [CrossRef]
- Gleason, D.F. Histologic grading of prostate cancer: A perspective. Hum. Pathol. 1992, 23, 273–279. [Google Scholar] [CrossRef]
- Ganti, A.K.; Klein, A.B.; Cotarla, I.; Seal, B.; Chou, E. Update of Incidence, Prevalence, Survival, and Initial Treatment in Patients With Non–Small Cell Lung Cancer in the US. JAMA Oncol. 2021, 7, 1824–1832. [Google Scholar] [CrossRef]
- Galluzzi, L.; Vitale, I.; Michels, J.; Brenner, C.; Szabadkai, G.; Harel-Bellan, A.; Castedo, M.; Kroemer, G. Systems biology of cisplatin resistance: Past, present and future. Cell Death Dis. 2014, 5, e1257. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.S. Chemotherapy Resistance in Lung Cancer. Adv. Exp. Med. Biol. 2016, 893, 189–209. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-M.; Yang, F.; Li, J.; Yuan, W.-Q.; Wang, H.; Luo, Y.-Q. Reelin Promotes Cisplatin Resistance by Induction of Epithelial-Mesenchymal Transition via p38/GSK3β/Snail Signaling in Non-Small Cell Lung Cancer. J. Pharmacol. Exp. Ther. 2020, 26, e925298-1–e925298-11. [Google Scholar] [CrossRef]
- Brown, F.C.; Cifani, P.; Drill, E.; He, J.; Still, E.; Zhong, S.; Balasubramanian, S.; Pavlick, D.; Yilmazel, B.; Knapp, K.M.; et al. Genomics of primary chemoresistance and remission induction failure in paediatric and adult acute myeloid leukaemia. Br. J. Haematol. 2016, 176, 86–91. [Google Scholar] [CrossRef]
- Sato, N.; Fukushima, N.; Chang, R.; Matsubayashi, H.; Goggins, M. Differential and Epigenetic Gene Expression Profiling Identifies Frequent Disruption of the RELN Pathway in Pancreatic Cancers. Gastroenterology 2006, 130, 548–565. [Google Scholar] [CrossRef]
- Li, X.; Fan, W.; Yao, A.; Song, H.; Ge, Y.; Yan, M.; Shan, Y.; Zhang, C.; Li, P.; Jia, L. Downregulation of reelin predicts poor prognosis for glioma. Biomarkers Med. 2020, 14, 651–663. [Google Scholar] [CrossRef]
- Ndoye, A.; Miskin, R.P.; DiPersio, C.M. Integrin α3β1 Represses Reelin Expression in Breast Cancer Cells to Promote Invasion. Cancers 2021, 13, 344. [Google Scholar] [CrossRef]
- Stein, T.; Cosimo, E.; Yu, X.; Smith, P.R.; Simon, R.; Cottrell, L.; Pringle, M.-A.; Bell, A.K.; Lattanzio, L.; Sauter, G.; et al. Loss of Reelin Expression in Breast Cancer Is Epigenetically Controlled and Associated with Poor Prognosis. Am. J. Pathol. 2010, 177, 2323–2333. [Google Scholar] [CrossRef]
- Okamura, Y.; Nomoto, S.; Kanda, M.; Hayashi, M.; Nishikawa, Y.; Fujii, T.; Sugimoto, H.; Takeda, S.; Nakao, A. Reduced Expression of Reelin (RELN) Gene Is Associated With High Recurrence Rate of Hepatocellular Carcinoma. Ann. Surg. Oncol. 2010, 18, 572–579. [Google Scholar] [CrossRef]
- Luo, Y.; Huang, K.; Zheng, J.; Zhang, J.; Zhang, L. TGF-β1 promotes cell migration in hepatocellular carcinoma by suppressing reelin expression. Gene 2018, 688, 19–25. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calvier, L.; Alexander, A.; Marckx, A.T.; Kounnas, M.Z.; Durakoglugil, M.; Herz, J. Safety of Anti-Reelin Therapeutic Approaches for Chronic Inflammatory Diseases. Cells 2024, 13, 583. https://doi.org/10.3390/cells13070583
Calvier L, Alexander A, Marckx AT, Kounnas MZ, Durakoglugil M, Herz J. Safety of Anti-Reelin Therapeutic Approaches for Chronic Inflammatory Diseases. Cells. 2024; 13(7):583. https://doi.org/10.3390/cells13070583
Chicago/Turabian StyleCalvier, Laurent, Anna Alexander, Austin T. Marckx, Maria Z. Kounnas, Murat Durakoglugil, and Joachim Herz. 2024. "Safety of Anti-Reelin Therapeutic Approaches for Chronic Inflammatory Diseases" Cells 13, no. 7: 583. https://doi.org/10.3390/cells13070583
APA StyleCalvier, L., Alexander, A., Marckx, A. T., Kounnas, M. Z., Durakoglugil, M., & Herz, J. (2024). Safety of Anti-Reelin Therapeutic Approaches for Chronic Inflammatory Diseases. Cells, 13(7), 583. https://doi.org/10.3390/cells13070583